Интернет-журнал "Домашняя лаборатория", 2008 №1 [Журнал «Домашняя лаборатория»] (fb2) читать онлайн
- Интернет-журнал "Домашняя лаборатория", 2008 №1 4.03 Мб, 634с. скачать: (fb2) читать: (полностью) - (постранично) - Журнал «Домашняя лаборатория»
[Настройки текста] [Cбросить фильтры]
[Оглавление]
Журнал «Домашняя лаборатория» 2008, № 1
ИСТОРИЯ
Баллада о вересковом мёде
Р. Стивенсон
Heather Ale
From the bonny bells of heather
They brewed a drink long-syne,
Was sweeter far than honey,
Was stronger far than wine.
They brewed it and they drank it,
And lay in a blessed swound
For days and days together
In their dwellings underground.
There rose a king in Scotland,
A fell man to his foes,
He smote the Piets in battle,
He hunted them like roes.
Over miles of the red mountain
He hunted as they fled,
And strewed the dwarfish bodies
Of the dying and the dead.
Summer came in the country,
Red was the heather bell;
But the manner of the brewing
Was none alive to tell.
In graves that were like children's
On many a mountain head,
The Brewsters of the Heather
Lay numbered with the dead.
The king in the red moorland
Rode on a summer's day;
And the bees hummed, and the curlews
Cried beside the way.
The king rode, and was angry,
Black was his brow and pale,
To rule in a land of heather
And lack the Heather Ale.
It fortuned that his vassals,
Riding free on the heath,
Came on a stone that was fallen
And vermin hid beneath.
Rudely plucked from their hiding,
Never a word they spoke:
A son and his aged father —
Last of the dwarfish folk.
The king sat high on his charger,
He looked on the little men;
And the dwarfish and swarthy couple
Looked at the king again.
Down by the shore he had them;
And there on the giddy brink —
"I will give you life, ye vermin,
For the secret of the drink."
There stood the son and father
And they looked high and low;
The heather was red around them,
The sea rumbled below.
And up and spoke the father,
Shrill was his voice to hear:
"I have a word in private,
A word for the royal ear.
"Life is dear to the aged,
And honour a little thing;
I would gladly sell the secret,"
Quoth the Piet to the King.
His voice was small as a sparrow's,
And shrill and wonderful clear:
"I would gladly sell my secret,
Only my son I fear.
"For life is a little matter,
And death is nought to the young;
And I dare not sell my honour
Under the eye of my son.
Take HIM, О king, and bind him,
And cast him far in the deep;
And it's I will tell the secret
That I have sworn to keep."
They took the son and bound him,
Neck and heels in a thong,
And a lad took him and swung him,
And flung him far and strong,
And the sea swallowed his body,
Like that of a child of ten; —
And there on the cliff stood the father,
Last of the dwarfish men.
"True was the word I told you:
Only my son I feared;
For I doubt the sapling courage
That goes without the beard.
But now in vain is the torture,
Fire shall never avail:
Here dies in my bosom
The secret of Heather Ale."
Вересковый мёд
Из вереска напиток
Забыт давным-давно.
А был он слаще мёда,
Пьянее, чем вино.
В котлах его варили
И пили всей семьей
Малютки медовары
В пещерах под землей.
Пришел король шотландский.
Безжалостный к врагам,
Прижал он бедных пиктов
К скалистым берегам.
На вересковом поле,
На поле боевом
Лежал живой на мертвом
И мертвый — на живом.
Лето в стране настало,
Вереск опять цветет,
Но некому готовить
Вересковый мёд.
В своих могилках тесных,
В горах родной земли
Малютки медовары
Приют себе нашли.
Король по склону едет
Над морем на коне,
И рядом реют чайки
С дорогой наравне.
Король глядит угрюмо:
«Опять в краю моем
Цветет медвяный вереск,
А меда мы не пьем!»
Но вот его вассалы
Приметили двоих
Последних медоваров,
Оставшихся в живых.
Вышли они из-под камня,
Щурясь на белый свет,
— Старый горбатый карлик
И мальчик пятнадцати лет.
К берегу моря крутому
Их привели на допрос,
Но ни один из пленных
Слова не произнес.
Сидел король шотландский,
Не шевелясь, в седле.
А маленькие люди
Стояли на земле.
Гневно король промолвил:
— Пытка обоих ждет,
Если не скажете, черти,
Как вы готовили мёд!
Сын и отец молчали,
Стоя у края скалы.
Вереск звенел над ними,
В море катились валы.
И вдруг голосок раздался:
— Слушай, шотландский король
Поговорить с тобою
С глазу на глаз позволь!
Старость боится смерти.
Жизнь я изменой куплю,
Выдам заветную тайну!
— Карлик сказал королю.
Голос его воробьиный
Резко и четко звучал:
— Тайну давно бы я выдал,
Если бы сын не мешал!
Мальчику жизни не жалко,
Гибель ему нипочем.
Мне продавать свою совесть
Совестно будет при нем.
Пускай его крепко свяжут
И бросят в пучину вод —
И я научу шотландцев
Готовить старинный мёд!..
Сильный шотландский воин
Мальчика крепко связал
И бросил в открытое море
С прибрежных отвесных скал.
Волны над ним сомкнулись.
Замер последний крик…
И эхом ему ответил
С обрыва отец-старик:
— Правду сказал я, шотландцы
От сына я ждал беды.
Не верил я в стойкость юных,
Не бреющих бороды.
А мне костер не страшен.
Пускай со мной умрет
Моя святая тайна
— Мой вересковый мёд!
Перевод С. Маршака
Шотландский вересковый эль (часть статьи)
Брюс Уиллиамс
Вересковый эль без сомнения самое старое пивоваренное наследие Шотландии. Напиток готовили следующим образом: сначала делали затор из шотландского элевого солода, варили сусло с цветущими верхушками вереска, затем засыпали его поверхность свежими цветами вереска[1], оставляли охладиться и сбраживали 12 дней, пока вереск не почернеет. Эль пили прямо из емкости, называвшейся сгап (бочка), в которой на четверть по высоте проделывалось отверстие для крана. Это янтарный, слегка газированный эль с мягкой горечью, крепким маслянистым телом и виноподобным финалом — поинтересуйтесь у Майкла Джексона. Этот напиток во времена Старого Альянса (Auld Alliance) в 18-ом веке французы называли шотландским бургундским, а англичане шотландской мальвазией. История приготовления верескового эля имеет древнюю традицию. Во время археологических раскопок на шотландском острове Rhum были обнаружены черепки эпохи неолита, датированные приблизительно 2000 лет до н. э., на которых были найдены следы напитка брожения, содержащего вереск. Намного позже, приблизительно 100 лет до н. э. было известно, что совершенно определенный европейский народ (кельты) готовили опьяняющий отвар из цветов вереска и меда, но самое богатое наследие по изготовлению верескового эля относится к временам, когда Шотландия носила название Piсtland (земля пиктов). Известный в четвертом веке мореплаватель Пифей (Pytheas) отмечал, что пикты (Piets) были искусными пивоварами, и в Шотландском словаре говорилось, что "пикты варят некий великий и ужасный напиток, называемый вересковым элем, из вереска и некоего неведомого fogg'а[2]". Короли Пиктланда отразили многие вторжения европейцев, англосаксов и ирландцев, и они даже прогнали восвояси силы могущественной Римской империи. За это они обрели славу свирепого народа, обладающего секретным волшебным зельем, которое называлось вересковым элем. Одна легенда, которая жива и по сей день, была записана Нейлом Мунро, сэром Гербертом Максвеллом, и она послужила темой для поэмы Роберта Льюиса Стивенсона — ее изложение отличается лишь в деталях. В 400 году нашей эры ирландский король вторгся в Далриаду (Dalriada) на юго-западе Шотландии и начал истреблять местных пиктов. Считается, что охваченный диким желанием выиграть сражение, он убил их всех прежде, чем вспомнил о существовании верескового эля. Он послал свою армию, чтобы найти выживших, и они вернулись с вождем пиктов и его сыном. Ирландский король собирался уже пытать их, чтобы выведать тайну верескового эля, когда вождь вдруг согласился сообщить тайну при условии, что они убьют его сына быстро. Как только его сын был умерщвлен, старый вождь привел ирландского короля к утесу, где рос урожай вереска, и, согласно Роберту Льису Стивенсону, промолвил: "Но теперь напрасна пытка, огонь тут не поможет, вот умирает в моей груди тайна верескового эля." (В переводе Андрея Кузнецова это звучит так:
«Плевать мне на ваши пытки,
Со мною исчезнет в огне,
Напитка из вереска тайна,
Известная только мне»).
БЕСПОКОЙСТВО
«Блеск и нищета» высшего образования в России
Кастуева-Жан Т.
Россия унаследовала от советского периода репутацию страны с высокообразованным населением и эффективной системой образования. На самом деле ситуация менее однозначна. За большой популярностью высшего образования и успехами некоторых крупных университетов и институтов скрываются снижение качества образования и некоторое обесценивание дипломов. Попытки реформирования наталкиваются на структурные проблемы образовательной системы, а также на коррупцию в педагогической среде. Такая ситуация затрудняет реальную оценку российских дипломов. Кроме того, «приватизация» большей части образовательных услуг остро ставит проблему равенства шансов.
Введение
Высокий уровень образованности населения обусловил научные и в технические достижения страны в советский период. Для советских людей образовательная и научно-исследовательская система страны, полностью финансируемая из государственного бюджета, долгое время оставалась наиболее ощутимым достижением социалистического строя. После распада СССР, экономический кризис и резкое сокращение государственного финансирования вынудили учебные заведения искать стратегии выживания. В результате реформ, начатых в начале постсоветского периода с целью модернизации и децентрализации образовательной системы, государственные учебные заведения получили большую автономию. Были также созданы условия для развития частных образовательных учреждений. За очень короткий срок возник рынок образовательных услуг, который с тех пор неуклонно растёт. Несмотря на призывы педагогического сообщества к «спасению» российского образования, которому угрожают недостаток государственного финансирования и последствия сокрушительного распада системы, уровень образования в России продолжает по инерции восприниматься — как внутри страны, так и за рубежом — как в целом высокий. Этот потенциал часто упоминается среди главных козырей, которые могут стать залогом мощи и возрождения России. Представители российской политический элиты постоянно подчёркивают значение этого предполагаемого потенциала для экономического роста в условиях жёсткой международной конкуренции. Так, в одном из своих выступлений на заседании Госсовета в августе 2001 года Владимир Путин подчеркнул, что образование «стало дорогим и самым ценным товаром, а устойчивое развитие стран уже давно определяется не столько их ресурсами, сколько общим уровнем образования нации». Образование фигурирует в числе четырёх национальных проектов, заявленных в сентябре 2005 г., и вошло в повестку дня встречи большой восьмёрки в Санкт-Петербурге в июле 2006 г. Между тем, за этими заявлениями и общим впечатлением качества образовательной системы скрываются противоречия и контрасты. Повышенный спрос на высшее образование и успехи отдельных учебных заведений не могут заслонить собой общего снижения качества образования, усиливающегося неравенства возможностей и обесценивания дипломов.
Количество или качество?
Повышенный спрос на высшее образование По количественным показателям Россия входит в группу самых образованных стран мира[3]. Что касается сферы высшего образования, в 1995–1996 гг. в 762 российских вузах обучалось 2,791 миллиона студентов; а в 2005–2006 гг. уже 1068 вузов (655 государственных и 413 частных) ведут подготовку 7,064 миллионов студентов[4]. Этот стремительный рост происходит на фоне демографического спада в стране, который уже начинает проявляться в снижении количества дошкольных и средних учебных заведений. Сегодня Россия насчитывает больше студентов, чем за всю свою предыдущую историю, включая и советский период. Количество студентов государственных вузов в 2004–2005 гг. составило 419 человек на 10 000 населения — цифра тем более значительная, что на то же количество населения выделяется лишь 170 бюджетных мест. И этот показатель постоянно увеличивается с 1996 г., после окончания периода явного спада спроса на высшее образование между 1990 и 1995 гг. Сегодня больше, чем когда-либо, в глазах россиян образование является приоритетом, оправдывающим вложение средств. По некоторым опросам общественного мнения образовательные услуги стали первой статьей расходов российских семей, обгоняя семейные сбережения и расходы на медицинское обслуживание[5]. Речь идёт о рынке будущего, поскольку наблюдается настоящий бум в сфере высшего образования, которое стремятся дать своим детям как минимум 85 % российских семей. Так же высока тяга к обучению в аспирантуре и получению учёных степеней. В 2003 году количество аспирантов превосходило в 2,5 раза их количество в 1995 году (140 741 против 62317)[6]. В 2005 году Российская государственная библиотека зарегистрировала рекордное количество диссертаций — 31000 (для сравнения — 27000 в 2004 г.)[7]. Наличие диссертации в России уже давно негласно рассматривается как необходимое условие для исполнения ответственных политических или управленческих должностей. Политики, чиновники высшего уровня и бизнесмены стремятся к приобретению учёных степеней. В апреле 2006 г. Независимая газета опубликовала сведения о количестве «кандидатов VIР» в действительные члены или члены-корреспонденты Российской академии наук[8]. Среди депутатов Думы (450 человек), например, насчитывается 143 кандидата наук, 71 доктор наук, а 14 депутатов имеют две учёные степени в разных областях науки.
О качестве дипломов
По инерции или в силу национальной гордости большая часть россиян продолжает считать свою систему образования одной из лучших в мире. Так, после недавнего визита в Оксфорд губернатор Свердловской области Э. Россель заявил, что не увидел там «ничего особенного», а «с точки зрения среднего и высшего образования мы далеко впереди Оксфорда[9]». Опросы общественного мнения далеко не так однозначны[10], а международные исследования на эту тему даже бьют тревогу. Так, если говорить о среднем образовании, Россия находится на последних позициях в рейтинге, составленном по результатам тестов PISA (Program for International Student Assessment), проведённых Организацией экономического сотрудничества и развития (ОЭСР) в 2000 и 2003 гг. среди пятнадцатилетних школьников[11]. В 2000 г. из 32-х стран-участниц российские подростки заняли лишь 27-е место по чтению (интерпретация текста, способность его анализировать, делать выводы, формулировать собственную точку зрения), 26-е по естественным наукам и 21-е по математике. В 2003 году Россия оказалась соответственно на 32-ом, 24-ом и 29-ом местах среди 41 страны. Тесты свидетельствуют о трудностях, возникающих у российских учеников при необходимости применить полученные знания вне школьных уроков, использовать противоречивую или неполную информацию, а также о нехватке критического мышления. Эти результаты, совершенно не соответствующие имеющимся у россиян представлениям об уровне среднего образования в стране, вызвали шок в педагогической среде. Многие педагоги выступают отныне за подход, нацеленный не на приобретение знаний, а на развитие компетенций (так называемый компетентностный подход), определяемых как способность эффективно реагировать на незнакомые ситуации[12]. Что касается сравнений в области высшего образования, можно указать международный рейтинг, опубликованный в августе 2006 года Шанхайским институтом высшего образования. Среди 500 лучших мировых университетов в нём фигурируют лишь два российских: Московский государственный университет — МГУ на 72-ом месте (21-е место в рейтинге европейских университетов) и Санкт-Петербургский государственный университет на 340-ом[13]. Для сравнения, рейтинг включает 22 французских университета, 20 китайских, 4 бразильских, 2 польских, и большое количество американских университетов, занимающих бесспорные лидирующие позиции. Этот рейтинг демонстрирует своего рода международную маргинализацию большинства российских вузов, подтверждающуюся и относительно небольшим числом иностранных студентов, проходящих обучение в России. Например, в 2003 году Россия приняла 54 200 иностранных студентов (для сравнения, в том же году в США обучались 558 000 иностранных студентов, в Германии — 240 619, в Великобритании — 255 233 и во Франции — 221 567)[14], из которых значительная часть обучалась на языковых курсах. Доля России на мировом рынке образовательных услуг не превышает 0,5 %[15]. Такую ситуацию объясняет целый ряд различных факторов: относительная трудность русского языка, слабая привлекательность политического и экономического имиджа страны, условия жизни и т. д. Однако, не последней причиной является и недостаточная конкурентоспособность российского образования, в целом ещё слабо интегрированного в европейские и мировые схемы, несмотря на недавнее (2003) присоединение России к болонскому процессу. Основным критерием оценки молодого специалиста является его способность найти работу по профилю на рынке труда и удовлетворять требованиям работодателя. Между тем, исследования, проведённые агентством РейтОР, специализирующимся на рейтинговании российских образовательных ресурсов, в одном из семи федеральных округов России (Приволжском), подчёркивают неудовлетворённость работодателей качеством принятых на работу новоиспечённых выпускников вузов[16]. Помимо завышенной оценки своих знаний и требований высокой зарплаты, работодатели отмечают нехватку как практических, так и теоретических знаний у молодых специалистов[17]. Более половины опрошенных в 2004–2005 гг. работодателей считают, что молодые специалисты нуждаются в дополнительной подготовке[18]. По свидетельству многих руководителей кадровых агентств, лишь 13–20 % предприятий готовы принять на работу молодых специалистов[19]. Впрочем, студенты, по всей видимости, и сами осознают недостатки своих дипломов. То же исследование в Приволжском округе показало, что 59 % студентов желали продолжить образование по завершению первично пройденного пятилетнего цикла, из них 52 % заявили о готовности самим финансировать его и 26 % — о желании приобрести другую специальность[20]. При прочих равных условиях (без учёта опыта работы), решающим фактором при подборе кадров на сегодняшнем российском рынке труда является второе высшее образование. В ходе различных встреч между представителями деловых кругов, ответственными лицами из министерства образования и ректорами вузов[21], первые регулярно указывают на архаизм содержания учебных курсов и методов обучения, цель которых заключается в накоплении знаний в ущерб самообразованию и развитию критического мышления. Они ратуют за обучение в форме проектов и case-studies. Эти предложения не вызывают одобрения большей части профессуры (в частности, ректоров, объединённых в могущественный Союз ректоров России), приверженной сохранению «фундаментального характера» российского образования. Другая проблема: согласно работодателям, профиль дипломированных выпускников не в полной мере удовлетворяет потребностям экономики страны. Так, например, рынок переполнен экономистами и юристами[22], в то время как в промышленности существует реальная нехватка руководителей среднего звена, инженеров и технических менеджеров (в отличие, например, от Индии, которая сделала особую ставку на подготовку инженеров). Интерес промышленных групп к образовательной сфере проявляется в стремлении ассоциироваться с вузами или создать собственные учебные заведения или факультеты, например, в виде «корпоративных университетов»[23], начавших развиваться с 2000 г. Среди первых предприятий, профинансировавших подобные инициативы — Северсталь, владелец которой, А. Мордашов, защитил свой диплом MBA по этой теме. Другие предприятия последовали примеру Северстали, чтобы обеспечить своему персоналу надлежащий уровень обучения: Сухой (авиация), Билайн (телекоммуникации), РусАл (алюминий) и др. Другим примером сотрудничества между вузами и предприятиями является подписание в апреле 2006 г. соглашения между РуссНефтью и МГУ с целью создания на базе Московского университета «Высшей школы инновационного бизнеса МГУ» со статусом факультета. МГУ должен осуществлять подбор профессорско-преподавательского состава, а РуссНефть может принимать участие в разработке учебных программ и планов, которые будут удовлетворять потребностям компании. Стоит также упомянуть соглашение между О. Дерипаской, владельцем РусАла, и Экономической академией им. Плеханова, воспринимаемое корпусом ректоров как своего рода революцию. Оно послужит пробным шаром для нового статуса автономной некоммерческой организации (АНО), предложенного правительством, и даст предпринимателю реальное droit de regard над программой. Этот проект вызывает обеспокоенность многих ректоров, видящих в нём начало скрытой «приватизации» вузов.
Причины «нищеты»
Б целом, политические элиты и другие общественные круги осознают снижение качества высшего образования в стране: В. Путин и министр образования А. Фурсенко не раз публично признавали этот факт. Но союз ректоров, деловые круги, эксперты и министерство образования расходятся во мнениях о причинах снижения качества. Для ректоров оно связано, с одной стороны, с недостатком государственного финансирования, что приводит к нехватке оборудования и низкому уровню оплаты труда преподавателей. С другой стороны, ответственность за это несёт быстрый рост сектора платных образовательных услуг и появление многих частных вузов или филиалов сомнительного качества. По мнению ректоров, ультралиберальная ориентация в сочетании с попытками правительства «привить» западные модели на российскую почву разрушают традиционные основы образовательной системы страны[24]. В свою очередь, чиновники министерства образования и деловые круги говорят о консерватизме ректоров и их сопротивлении реформам под влиянием корпоративных интересов и стремления защитить принцип финансовой автономии вуза без внешнего контроля.
Финансирование Между 1992 и 1998 гг., сумма бюджетных средств, выделяемая на образование, сократилась в три раза. В этот период учебные заведения с трудом могли покрыть свои текущие расходы и обеспечить выплату зарплат. С 1999 г. наблюдается рост государственного финансирования: в 2004 году его объём увеличился на треть по сравнению с серединой девяностых годов. Но, если доля образования в государственных расходах России сравнима со средним значением стран ОЭСР (12,7 %), уровень финансирования образования в процентном соотношении к ВВП намного ниже: в течение последних трёх лет он варьировался между 3,5 и 3,8 % от ВВП (из которых 0,5 % приходится на высшее образование), в то время как средний показатель по ОЭСР составляет 6,2 %[25]. Эти данные далеки от поставленной российскими властями цели — 10 % от ВВП. Расходы на одного студента составляют 23000-25000 рублей в год (около 700–800 евро). В январе 2006 года во время встречи со студентами сельскохозяйственной академии им. Тимирязева, министр Фурсенко признал эту сумму недостаточной и предложил увеличить её до 75000 рублей с тем, чтобы приблизиться по этому показателю к западным странам (во Франции, например, в 2003 году годовые расходы на одного студента вуза составили 8 370 евро[26]). Закон об образовании от 1992 года позволил учебным заведениям полу чить статус юридических лиц и, соответственно, распоряжаться своими ресурсами. Многие школы и вузы бросились изыскивать средства для выживания разнообразными способами: сдавать часть своих помещений в аренду, создавать подсобные хозяйства, автошколы, привлекать спонсоров, взимать плату за многие услуги и создавать факультеты и отделения в соответствии с запросами родителей учеников и студентов. По официальным данным, объём платных услуг в сфере образования возрос от 41 млрд. руб. (1,17 млрд. евро) в 2000 г. до 147,5 млрд. руб. в 2005 г[27]. Около 56 % студентов сегодня сами финансируют своё обучение[28]. Опросы общественного мнения показывают, что идея оплаты высшего образования практически прижилась в менталитете россиян, готовых отныне инвестировать в эту сферу. Однако, в нынешней системе явно наблюдается перекос в сторону платных услуг: платить иногда приходится и за пропуски, и за пересдачу экзаменов и зачетов и т. д. В начале обучения детей родители зачастую не имеют чёткого представления о конечной сумме, которую им придётся затратить. Кроме того, к «платным студентам» нередко наблюдается иное отношение, нежели к «бюджетникам».
Педагогические кадры Объём государственного финансирования не позволяет обеспечить привлекательный уровень зарплат в секторе образования. Средняя зарплата преподавателя в средней школе составляет 9300 рублей (270 евро) в Москве и 3900 рублей (113 евро) в регионах, а в высшем учебном заведении соответственно 7500 и 5000 рублей (217 и 145 евро)[29]. Для сравнения, в 2004 году средняя зарплата в образовании составляла 62 % от средней зарплаты в России по всем секторам и 53 % от средней зарплаты в промышленности[30]. Зарплата молодого преподавателя вуза ниже прожиточного минимума. Как и в секторе научных исследований, преподаватели вузов вынуждены искать другие источники дохода, такие как репетиторство и т. п. Эти дополнительные доходы могут достигать официальной зарплаты и даже превышать её. Эта ситуация приводит иногда к практически фиктивной принадлежности преподавателя к вузу. В таких условиях неудивительно, что выпускники вузов не стремятся к работе в системе образования, а также то, что профессия преподавателя стареет и феминизируется, особенно в средней школе, где женщины составляют 85,3 %. Говоря о качестве преподавательского состава нужно также отметить разрыв между высшим образованием и наукой. Процент преподавателей, занимающихся исследовательской работой, относительно невелик. По словам ректора Высшей школы экономики Я. Кузьминова, 4/5 вузовских программ осуществляются преподавателями, не участвующими в научных исследованиях[31]. Среди 3797 российских научно-исследовательских заведений, вузы представляют лишь скромную часть (393).
Искажения в отношении общества к высшему образованию В российском обществе наблюдается некое искажение отношения к высшему образованию. В декабре 2005 г. А. Фурсенко, с сожалением отмечая утрату качества образования, объяснил его тем, что студенты прежде всего стремятся получить «бумажку», не заботясь о знаниях, предполагающих получение диплома[32]. Не говоря о тех, для кого поступление в вуз является способом избежать службы в армии[33] (по различным опросам эти лица составляют от 5 до 10 % абитуриентов) или преследует другие, не связанные с образованием цели, высшее образование стало жертвой своей популярности. Социологи констатируют, что оно всё в большей степени играет «социализирующую» роль: диплом служит своего рода социальным знаком качества. Студенты часто не связывают напрямую получаемое ими образование с будущей профессиональной карьерой. Они ведут себя так, как если бы целью проведения пяти лет в вузе являлось получение диплома, а профессии они реально смогут обучиться во время первого профессионального опыта. Такой подход имеет множественное объяснение. На выбор профессионального пути молодого россиянина влияют многие противоречивые факторы. С одной стороны, социальное давление и условия на рынке труда укрепляют привлекательность высшего образования, что, в сочетании с фактически негативным имиджем других типов учебных заведений (таких как профессиональные школы — бывшие ПТУ), подталкивает к выбору высшего образования в ущерб иному профессиональному. С другой стороны, это попытка адаптироваться к переходу к «экономике знаний»[34]. К тому же, рынку труда в России незнакомо понятие избытка квалификации для той или иной должности. Диплом о высшем образовании нередко требуется работодателем независимо от рабочего места как некая гарантия серьёзности кандидата. Рост числа вузов и их относительная доступность для студентов, «проваливших» вступительные экзамены на бюджетные места, способствуют укреплению тенденции к массовизации высшего образования. Кроме того, молодёжь в России заканчивает среднюю школу несколько раньше, чем на Западе (в 16–17 лет), и родители считают 5 лет учёбы в вузе периодом «дозревания», позволяющим создать некий «интеллектуальный капитал» до осуществления настоящего профессионального выбора. По окончанию этого периода приходит время второго высшего образования, — в некотором смысле, «настоящего» — получение которого и родители, и студенты готовы сами финансировать. Такое специфическое отношение к высшему образованию порождает практику разного рода мошенничества. В некотором смысле, это явление даже институционализировано. Многочисленные интернет-сайты предлагают уже готовые или на заказ в кратчайшие сроки всевозможные рефераты, дипломы или готовые ответы на экзаменационные билеты, содержание которых, по российской традиции, известно студентам заранее. Сайты предлагают даже такие технологические новинки, как ручки с невидимыми чернилами или программы, загружаемые на мобильные телефоны[35]. Процветает рынок дипломных и курсовых работ, выполненных на заказ. Можно найти подставных лиц, готовых сдать экзамены за абитуриента. Дипломные работы редко представляют собой настоящее исследование, в лучшем случае, речь идет об удачной компиляции уже существующих работ (по известному выражению «студенты делают вид, что пишут, а преподаватели — что проверяют»). Эта система естественным образом ведёт и к простой покупке дипломов, продажа которых в переходах московского метро продолжает шокировать иностранцев. Согласно недавнему опросу ФОМ, 76 % опрошенных (91 % среди москвичей) утверждают, что достать «липовый» диплом не составляет ни малейшей проблемы, 23 % уже встречали людей, купивших диплом, и 61 % считает, что фальшивый диплом открывает такие же возможности для профессиональной карьеры, что и настоящий[36].
Образование или взяточничество? Губительные последствия коррупции в образовании признаются на самом высоком уровне. С одной стороны, речь идёт об «откатах» — суммах, которые университеты оставляют чиновникам с целью добиться бюджетного финансирования, которые иногда оцениваются в 50 % от размера самих субсидий[37]. С другой стороны, — о взятках, которые родители студентов выплачивают вузам или непосредственно не слишком щепетильным преподавателям, чтобы заручиться особым подходом. Некоторые эксперты считают, что «чёрный оборот» некоторых институтов и университетов достигает оборота крупных нефтяных компаний[38]. В 2002–2003 гг. общая сумма взяток в сфере образования достигла, по некоторым сведениям, 26,4 млрд. рублей (754 млн. евро), из которых половина была выплачена за поступление в вуз и половина за само обучение[39]. Согласно одному из исследований, проведённых Высшей школой экономики, одна семья из пяти (и одна из четырёх в Москве) готова заплатить дополнительные деньги за поступление в престижную школу, а 18 % (24 % в Москве) готовы платить за поступление в престижный вуз[40]. Таким образом, спрос на подобный вид услуг высок. Объявления с предложением помощи для поступления в вуз встречаются в изобилии при общем толерантном отношении общества к ним. Исследовательская среда также не избежала коррупции. Некоторые аспиранты доверяют целиком или частично написание своих диссертаций «субподрядчикам». Клиенты VIP покупают свои диссертации «под ключ» (что может включать выбор темы, подбор аспирантуры и состав «доброжелательной» комиссии и т. п.)[41]. За определённую плату аспиранты могут опубликовать свои статьи (обязательные для защиты) в журналах, порой псевдонаучных[42]. Высшая аттестационная комиссия министерства образования и науки озабочена подобным явлением, поскольку, по некоторым данным, до трети диссертаций защищается лицами, не являющимися их авторами. Попытки контроля натыкаются на глубоко укоренившуюся практику. Например, правительственное решение о назначении ректоров (должность, которая стала выборной после распада СССР) с целью большего контроля встретило сильнейшее сопротивление со стороны ректоров, заговоривших о возврате к советской системе. В любом случае, эта мера вряд ли привела бы к уменьшению коррупции, настолько образовательная система изобилует возможностями приобретения символических и финансовых льгот.
Сохранение элитного образования
На фоне общего снижения качества образования сохраняется его элитная часть. Элитизм проявляется на всё более ранних стадиях обучения и во всё более яркой форме. На уровне среднего образования, наряду с обычными школами появились гимназии и лицеи: они предполагают строгий вступительный отбор (что может вызвать необходимость интенсивной подготовки шестилетнего ребёнка с родителями или репетитором). Уверенные в своих финансовых возможностях, эти школы могут позволить себе пригласить на работу лучших преподавателей и предложить качественное обучение. Чтобы набрать лучших выпускников, вузы заключают с ними соглашения; в старших классах обучение может вестись преподавателями вузов. Таким образом, их ученики оказываются в привилегированном положении на вступительных экзаменах. Так образуются своего рода герметические «коридоры», идущие иногда от детского сада до университета, в зависимости от уровня доходов семей[43]. Сомнения относительно качества диплома приводят к тому, что работодатели выбирают целевую группу для своих предложений: кадровые агентства всё чаще сталкиваются с тем, что многие предприятия требуют дипломы конкретных вузов. Среди этих вузов, в большинстве случаев, встречаются те, которые уже и в советскую эпоху имели репутацию элитных. Пользуясь своим имиджем, они попали в «положительную спираль», недоступную для «непосвящённых»: их имя и репутация привлекают молодёжь из семей с высоким достатком, что способствует дальнейшему финансированию. Эти вузы часто развивают партнёрские отношения с зарубежными высшими школами и университетами и создают уникальные совместные программы. Многочисленные рейтинги, зачастую противоречащие друг другу в силу избранных критериев, подпитывают социальное давление в погоне за поступлением в тот или иной престижный вуз. Но разброс между местами, занимаемыми в различных рейтингах одним и тем же вузом (см. приложение) доказывает всю сложность объективной оценки. Например, Санкт-Петербургский государственный университет, фигурирующий на первых местах рейтинга министерства образования, занимает лишь 15-е в рейтинге Коммерсанта; Финансовая академия, качество образования в которой не вызывает сомнения, получила лишь 28-е место в рейтинге Фонда Потанина; Московский институт стали и сплавов, не занявший высоких мест в рейтингах, был отобран как инновационный вуз для финансирования в рамках национального проекта. В целом, около 20 вузов, в подавляющем большинстве московских, составляют сердцевину образовательной системы России: среди них Московский государственный университет им. Ломоносова, Московский государственный институт международных отношений (МГИМО-Университет), Московский государственный технический университет им. Баумана, Экономическая академия им. Плеханова, Финансовая академия, Московский физико-технический институт, Высшая школа экономики, Российский государственный гуманитарный университет, Московская юридическая академия и некоторые другие. За пределами Москвы имеют хорошую репутацию Санкт-Петербургский государственный университет, Казанский государственный университет и некоторые другие. Эти заведения известны не только качеством образования, но и социальным статусом своих выпускников. Исследование, проведённое в 2005 г. агентством РейтОР, касающееся первого высшего образования 505 высокопоставленных представителей российской политической элиты (правительство, президентская администрация, полномочные представители президента в федеральных округах, Совет безопасности, губернаторы), показало, что 40 % из них получили образование в московских вузах. За ними следуют центральная и северноевропейская часть России (17 %), Санкт-Петербург (16 %), Дальний Восток (16 %), СНГ (6 %) и юг страны (4 %). Тройка лидеров включает МГУ (37 человек), СПГУ (16 человек) и МГИМО (15человек). За ними следуют Московская академия им. Плеханова, Финансовая и Юридическая академии. Российское государство поддерживает лидирующие вузы, которыми руководят близкие к политическим кругам влиятельные ректоры. МГУ финансируется отдельной строкой бюджета. Ректор СПГУ Л. Вербицкая недавно выразила удовлетворение значительным ростом с 2000 г. государственного финансирования, выделяемого своему вузу. В итоге, две трети финансирования alma mater президента Путина и его двух дочерей обеспечивается за счёт государственного бюджета. Эти два университета, представляющие гордость и достояние России, должны, по словам ректора СПГУ, обязательно финансироваться из государственного бюджета[44]. Самые престижные вузы наиболее сдержанно относятся к идее замены вступительных экзаменов Единым государственным экзаменом (ЕГЭ), введённым в большинстве регионов. Они выступают за сохранение возможности поступления на особых условиях для победителей региональных и федеральных Олимпиад, что является способом привлечения лучших выпускников. Национальный проект Образование, заявленный В. Путиным в сентябре 2005 г., укрепляет эту тенденцию к элитизму, ставя акцент на поддержке лидеров. Вместо распыления кредитов было решено выделить их наиболее успешным. В мае 2006 года министерство опубликовало список 17 инновационных вузов, которые получат финансирование (на общую сумму в 10 млрд. рублей или 285 млн. евро). Наиболее крупные суммы (9950 млн. руб. каждому) будут выделены. МГУ и СПГУ[45].
Перспективы
В российском высшем образовании сложилась разнородная картина: успехи элитных вузов выделяются на фоне снижения среднего уровня российских дипломов. Эта ситуация является результатом пятнадцати лет приспособления и выживания образовательных структур в условиях недостаточного финансирования. Большой заслугой российской образовательной системы является уже то, что она сумела выжить, сохранить свою целостность, удержать своих лидеров, и даже найти определённое, хотя и хрупкое, равновесие. Боязнью нарушить его объясняется сегодняшняя сдержанность ректоров и профессорско-преподавательского состава по отношению к предлагаемым правительством реформам. Сегодняшний выбор политики российского правительства очевиден: государство окажет поддержку только лучшим. Совершенно очевидно, что не будет распыления финансирования для спасения аутсайдеров или уменьшения разрыва между лидерами и всеми остальными. Национальный проект недвусмысленно ставит акцент на поддержке ограниченного числа «точек роста». Он ратует также за создание двух государственных университетов и бизнес-школы, конкурентоспособных на международном уровне. Осуществление реформы ГИФО (система государственных именных финансовых обязательств) также приведет к тому, что деньги пойдут за студентом в вузы, пользующиеся наибольшим спросом. Таким образом, смогут выжить только самые сильные или самые новаторские вузы. На первый взгляд эта политика не только противоречит устремлениям профессорско-преподавательского состава, но и массовому спросу на высшее образование среди населения. Однако, демографические прогнозы тоже говорят в пользу подобного «естественного отбора». К 2015 году количество детей в России уменьшится на 38 % в возрастной группе от 5 до 14 лет и на 47 % среди подростков 15–19 лет по сравнению с 2000 годом[46]. Возникнет избыточность школ и вузов (количество мест в вузах будет практически совпадать с количеством выпускников средних школ). Демографический спад приведёт к естественному количественному выравниванию системы (закрытие или объединение некоторых школ и вузов; массовый выход на пенсию пожилых преподавателей; большее финансирование на одного учащегося при том же общем объёме финансирования и т. д.) Это выравнивание может создать благоприятные условия для повышения качества образования. Подобная ситуация может стать уникальной возможностью для России, которая стремится диверсифицировать свою экономику, не довольствоваться положением поставщика сырьевых ресурсов на международных рынках. Настоящий качественный прорыв представляется возможным при условии открытия как минимум двух больших строительных площадок. Во-первых, постепенное устранение коррупции. Эта практика, со всей очевидностью, не может быть истреблена в одночасье. Коррупция стала составной частью равновесия системы и поэтому с ней очень трудно бороться. Стоит, по-видимому, решать проблемы поочередно: условия приема, вступительные экзамены, наказание в наиболее вопиющих случаях… В соседней Украине с целью борьбы с коррупцией было решено полностью отменить вступительные экзамены в вузах. В России подобное решение было бы — во всяком случае на сегодняшний день — отвергнуто ректорами. Во-вторых, повышение престижа профессии преподавателя. Речь идет не о разовых подарках, как это делается в рамках национального проекта для преподавателей в сфере среднего образования, предоставляющего премии 10000 лучших, а об обеспечении уровня зарплат, гарантирующих преподавателям достойную жизнь и привлекательных для молодёжи. Воссоздание стабильной связи между преподаванием и научно-исследовательской деятельностью позволило бы преподавателям поддерживать высокий уровень своих знаний. Эти две меры послужили бы первыми шагами к продвинутой системе высшего образования, которая вновь могла бы стать козырем России, основой её национальной инновационной системы, а также инструментом soft power на международном уровне.
ПРИЛОЖЕНИЕ
Сопоставление рейтингов некоторых государственных вузов России[47]
Источники: <www.mon.gov.ru/proekt/shkola/2474/>, <www.korochka.ru/universities/rating/detail.php?ID=1064>, <www.reitor.ru>, <http://fund.potanin.ru/>.Приведённые рейтинги основываются на различных критериях: Министерство образования (2005) учитывает самые разнообразные критерии, среди которых много количественных (количество студентов, кв. метров аудиторий на одного студента, преподавателей с научным званием, иностранных студентов, научных исследований, публикаций, библиотек, общежитий и т. п.). Рейтинг проводился отдельно по каждому типу вузов (университет, технический вуз и т. д.), что объясняет наличие нескольких первых мест в таблице. Министр Фурсенко призвал принять во внимание и другие параметры. Следующий рейтинг министерства должен основываться на списке из 17 инновационных вузов, отобранных в мае 2006 года в рамках национального проекта. Фонд Потанина (2005–2006) оценивает индивидуальный и профессиональный потенциал студентов и молодых преподавателей соответствующих вузов. Агентство РейтОР (июнь 2005) объединяет результаты опросов среди педагогов, работодателей и СМИ по таким параметрам, как уровень зарплаты выпускников, быстрота карьерного роста. Газета Коммерсант (2006) базируется на количестве дипломированных специалистов, которые были приняты на работу (или запрошены) крупными предприятиями в 2005 г.



ЧИТАЛЬНЯ
Профессия
Азимов А.
Джордж Плейтен сказал с плохо скрытой тоской в голосе: — Завтра первое мая. Начало Олимпиады! Он перевернулся на живот и через спинку кровати пристально посмотрел на своего товарища по комнате. Неужели он не чувствует того же? Неужели мысль об Олимпиаде совсем его не трогает? У Джорджа было худое лицо, черты которого еще более обострились за те полтора года, которые он провел в приюте. Он был худощав, но в его синих глазах горел прежний неуемный огонь, а в том, как он сейчас вцепился пальцами в одеяло, было что-то от затравленного зверя. Его сосед по комнате на мгновение оторвался от книги и заодно отрегулировал силу свечения стены, у которой сидел. Его звали Хали Смани, он был нигерийцем. Темно-коричневая кожа и крупные черты лица Хали Смани, казалось, были созданы для того, чтобы выражать только одно спокойствие, и упоминание об Олимпиаде нисколько его не взволновало. — Я знаю, Джордж, — произнес он. Джордж многим был обязан терпению и доброте Хали; бывали минуты, когда он очень в них нуждался, но даже доброта и терпение могут стать поперек глотки. Разве сейчас можно сидеть с невозмутимым видом идола, вырезанного из дерева теплого, сочного цвета? Джордж подумал, не станет ли он сам таким же через десять лет жизни в этом месте, и с негодованием отогнал эту мысль. Нет! — По-моему, ты забыл, что значит май, — вызывающе сказал он. — Я очень хорошо помню, что он значит, — отозвался его собеседник. — Ровным счетом ничего! Ты забыл об этом, а не я. Май ничего не значит для тебя, Джорджа Плейтена… и для меня, Хали Смани, — негромко добавил он. — Сейчас на Землю за новыми специалистами прилетают космические корабли, — произнес Джордж. — К июню тысячи и тысячи этих кораблей, неся на борту миллионы мужчин и женщин, отправятся к другим мирам, и все это, по-твоему, ничего не значит? — Абсолютно ничего. И вообще, какое мне дело до того, что завтра первое мая? Беззвучно шевеля губами. Смани стал водить пальцем по строчкам книги, которую он читал, — видимо, ему попалось трудное место. Джордж молча наблюдал за ним. "К черту! — подумал он. — Закричи, завизжи! Это-то ты можешь? Ударь меня, ну, сделай хоть что-нибудь!" Лишь бы не быть одиноким в своем гневе. Лишь бы разделить с кем-нибудь переполнявшее его возмущение, отделаться от мучительного чувства, что только он, он один умирает медленной смертью! В те первые недели, когда весь мир представлялся ему тесной оболочкой, сотканной из какого-то смутного света и неясных звуков, — тогда было лучше. А потом появился Омани и вернул его к жизни, которая того не стоила. Омани! Он-то стар! Ему уже по крайней мере тридцать. "Неужели и я в этом возрасте буду таким же? — подумал Джордж. — Стану таким, как он, через каких-нибудь двенадцать лет?" И оттого, что эта мысль вселила в него панический страх, он заорал на Омани: — Брось читать эту идиотскую книгу! Омани перевернул страницу и, прочитав еще несколько слов, поднял голову, покрытую шапкой жестких курчавых волос. — А? — спросил он. — Какой толк от твоего чтения? — Джордж решительно шагнул к Омани, презрительно фыркнул: — Опять электроника! — и вышиб книгу из его рук. Омани неторопливо встал и поднял книгу. Без всякого раздражения он разгладил смятую страницу. — Можешь считать, что я удовлетворяю свое любопытство, — произнес он. — Сегодня я пойму кое-что, а завтра, быть может, пойму немного больше. Это тоже своего рода победа. — Победа! Какая там победа? И больше тебе ничего не нужно от жизни? К шестидесяти пяти годам приобрести четверть знаний, которыми располагает дипломированный инженер-электронщик? — А может быть, не к шестидесяти пяти годам, а к тридцати пяти? — Кому ты будешь нужен? Кто тебя возьмет? Куда ты пойдешь с этими знаниями? — Никому. Никто. Никуда. Я останусь здесь и буду читать другие книги. — Но этого тебе достаточно? Рассказывай! Ты заманил меня на занятия. Ты заставил меня читать и заучивать прочитанное. А зачем? Это не приносит мне никакого удовлетворения. — Что толку в том, что ты лишаешь себя возможности получать удовлетворение? — Я решил наконец покончить с этим фарсом. Я сделаю то, что собирался сделать с самого начала, до того как ты умаслил меня и лишил воли к сопротивлению. Я заставлю их… заставлю… Омани отложил книгу, а когда Джордж, не договорив, умолк, задал вопрос: — Заставишь, Джордж? — Заставлю исправить эту вопиющую несправедливость. Все было подстроено. Я доберусь до этого Антонелли и заставлю его признаться, что он… он… Омани покачал головой. — Каждый, кто попадает сюда, настаивает на том, что произошла ошибка. Мне казалось, что у тебя этот период уже позади. — Не называй это периодом, — злобно сказал Джордж. — В отношении меня действительно была допущена ошибка. Я ведь говорил тебе… — Да, ты говорил, но в глубине души ты прекрасно сознаешь, что в отношении тебя никто не совершил никакой ошибки. — Не потому ли, что никто не желает в этом сознаваться? Неужели ты думаешь, что кто-нибудь из них добровольно признает свою ошибку?.. Но я заставлю их сделать это. Во всем виноват был май, месяц Олимпиады. Это он возродил в Джордже былую ярость, и он ничего не мог с собой поделать. Да и не хотел: ведь ему грозила опасность все забыть. — Я собирался стать программистом вычислительных машин, и я действительно могу им быть, что бы они там ни говорили, ссылаясь на результаты анализа. — Он стукнул кулаком по матрасу. — Они не правы. И не могут они быть правы. — В анализах ошибки исключены. — Значит, не исключены. Ведь ты же не сомневаешься в моих способностях? — Способности не имеют к этому ровно никакого отношения. Мне кажется, что тебе достаточно часто это объясняли. Почему ты никак не можешь понять? Джордж отодвинулся от него, лег на спину и угрюмо уставился в потолок. — А кем ты хотел стать, Хали? — У меня не было определенных планов. Думаю, что меня вполне устроила бы профессия гидропониста. — И ты считал, что тебе это удастся? — Я не был в этом уверен. Никогда раньше Джордж не расспрашивал Омани о его жизни. Мысль о том, что у других обитателей приюта тоже были свои стремления и надежды, показалась ему не только странной, но даже почти противоестественной. Он был потрясен. Подумать только — гидропонист! — А тебе не приходило в голову, что ты попадешь сюда? — Нет, но, как видишь, я все-таки здесь. — И тебя это удовлетворяет. Ты на самом деле всем доволен. Ты счастлив. Тебе здесь нравится, и ничего другого ты не хочешь. Смани медленно встал и аккуратно начал разбирать постель. — Джордж, ты неисправим, — произнес он. — Ты терзаешь себя, потому что отказываешься признать очевидные факты. Ты находишься в заведении, которое называешь приютом, но я ни разу не слышал, чтобы ты произнес его название полностью. Так сделай это теперь, Джордж, сделай! А потом ложись в кровать и проспись. Джордж скрипнул зубами и ощерился. — Нет! — сказал он сдавленно. — Тогда это сделаю я, — сказал Омали, и, отчеканивая каждый слог, он произнес роковые слова. Джордж слушал, испытывая глубочайший стыд и горечь. Он отвернулся. В восемнадцать лет Джордж Плейтен твердо знал, что станет дипломированным программистом, — он стремился к этому с тех пор, как себя помнил. Среди его приятелей одни отстаивали космонавтику, другие — холодильную технику, третьи — организацию перевозок и даже административную деятельность. Но Джордж не колебался. Он с таким же жаром, как и все остальные, обсуждал преимущества облюбованной профессии. Это было вполне естественно. Впереди их всех ждал День образования — поворотный день их жизни. Он приближался, неизбежный и неотвратимый, — первое ноября того года, когда им исполнится восемнадцать лет. Когда День образования оставался позади, появлялись новые темы для разговоров: можно было обсуждать различные профессиональные вопросы, хвалить свою жену и детей, рассуждать о шансах любимой космобольной команды или вспоминать Олимпиаду. Но до наступления Дня образования лишь одна тема неизменно вызывала всеобщий интерес — и это был День образования…. "Кем ты хочешь быть? Думаешь, тебе это удастся? Ничегошеньки у тебя не выйдет. Справься в ведомостях — квоту же урезали. А вот логистика…" Или "а вот гипермеханика…", или "а вот связь…", или "а вот гравитика…" Гравитика была тогда самой модной профессией. За несколько лет до того, как Джорджу исполнилось восемнадцать лет, появился гравитационный двигатель, и все только и говорили, что о гравитике. Любая планета в радиусе десяти световых лет от звезды-карлика отдала бы правую руку, лишь бы заполучить хоть одного дипломированного инженера-гравитационника. Но Джорджа это не прельщало. Да, конечно, такая планета отдаст все свои правые руки, какие только сумеет наскрести. Однако Джордж слышал и о том, что случалось в других, только что возникших областях техники. Немедленно начнутся рационализация и упрощение. Каждый год будут появляться новые модели, новые типы гравитационных двигателей, новые принципы. А потом все эти баловни судьбы в один прекрасный день обнаружат, что они устарели, их заменят новые специалисты, получившие образование позже, и им придется заняться неквалифицированным трудом или отправиться на какую-нибудь захудалую планету, которая пока еще не догнала другие миры. Между тем спрос на программистов оставался неизменным из года в год, из столетия в столетие. Он никогда не возрастал стремительно, не взвинчивался до небес, а просто медленно и неуклонно увеличивался в связи с освоением новых миров и усложнением старых. Эта тема была постоянным предметом споров между Джорджем и Коротышкой Тревельяном. Как все закадычные друзья, они спорили до бесконечности, не скупясь на язвительные насмешки, и в результате оба оставались при своем мнении. Дело в том, что отец Тревельяна, дипломированный металлург, в свое время работал на одной из дальних планет, а его дед тоже был дипломированным металлургом. Естественно, что сам Коротышка не колеблясь остановил свой выбор на этой профессии, которую считал чуть ли не неотъемлемым правом своей семьи, и был твердо убежден, что все другие специальности не слишком-то респектабельны. — Металл будет существовать всегда, — заявил он, — и когда ты создаешь сплав с заданными свойствами и наблюдаешь, как слагается его кристаллическая решетка, ты видишь результат своего труда. А что делает программист? Целый день сидит за кодирующим устройством, пичкая информацией какую-нибудь дурацкую электронную машину длиной в милю. Но Джордж уже в шестнадцать лет отличался практичностью. — Между прочим, вместе с тобой будет выпущен еще миллион металлургов, — спокойно указал он. — Потому что это прекрасная профессия. Самая лучшая. — Но ведь ты попросту затеряешься в их массе, Коротышка, и можешь оказаться где-то в хвосте. Каждая планета может сама зарядить нужных ей металлургов, а спрос на усовершенствованные земные модели не так уж велик, да и нуждаются в них главным образом малые планеты. Ты ведь знаешь, какой процент общего выпуска дипломированных металлургов получает направление на планеты класса А. Я поинтересовался — всего лишь 13,3 процента. А это означает семь шансов из восьми, что тебя засунут на какую-нибудь третьесортную планету, где в лучшем случае есть водопровод. А то и вовсе можешь застрять на Земле — такие составляют 2,3 процента. — Не вижу в этом ничего позорного, — вызывающе заявил Тревельян. Земле тоже нужны специалисты. И хорошие. Мой дед был земным металлургом. — Подняв руку, Тревельян небрежно провел пальцем по еще не существующим усам. Джордж знал про дедушку Тревельяна, и, памятуя, что его собственные предки тоже работали на Земле, не стал ехидничать, а, наоборот, дипломатично согласился: — В этом, безусловно, нет ничего позорного. Конечно, нет. Однако попасть на планету класса А — это вещь, скажешь нет? Теперь возьмем программиста. Только на планетах класса А есть такие вычислительные машины, для которых действительно нужны высококвалифицированные программисты, и поэтому только эти планеты и берут их. К тому же ленты по программированию очень сложны и для них годится далеко не всякий. Планетам класса А нужно больше программистов, чем может дать их собственное население. Это же чистая статистика. На миллион человек приходится в среднем, скажем, один первоклассный программист. И если на планете живет десять миллионов, а им там требуется двадцать программистов, они вынуждены обращаться к Земле, чтобы получить еще пять, а то и пятнадцать специалистов. Верно? А знаешь, сколько дипломированных программистов отправилось в прошлом году на планеты класса А? Не знаешь? Могу тебе сказать. Все до единого! Если ты программист, можешь считать, что ты уже там. Так-то! Тревельян нахмурился. — Если только один человек из миллиона годится в программисты, почему ты думаешь, что у тебя это выйдет? — Выйдет, можешь быть спокоен, — сдержанно ответил Джордж. Он никогда не осмелился бы рассказать ни Тревельяну, ни даже своим родителям, что именно он делает и почему так уверен в себе. Он был абсолютно спокоен за свое будущее. (Впоследствии, в дни безнадежности и отчаяния, именно это воспоминание стало самым мучительным.) Он был так же непоколебимо уверен в себе, как любой восьмилетний ребенок накануне Дня чтения, этого преддверия следующего за ним через десять лет Дня образования. Ну, конечно. День чтения во многом отличался от Дня образования. Во-первых, следует учитывать особенности детской психологии. Ведь восьмилетний ребенок легко воспринимает многие самые необычные явления. И то, что вчера он не умел читать, а сегодня уже умеет, кажется ему само собой разумеющимся. Как солнечный свет, например. А во-вторых, от этого дня зависело не так уж много. После него толпы вербовщиков не теснились перед списками, с нетерпением ожидая, когда будут объявлены результаты ближайшей Олимпиады. День чтения практически ничего не менял в жизни детей, и они еще десять лет оставались под родительской кровлей, как и все их сверстники. Просто после этого дня они уже умели читать. И Джордж, готовясь к Дню образования, почти не помнил подробностей того, что произошло с ним в День чтения, десять лет назад. Он, правда, не забыл, что день выдался пасмурный и моросил сентябрьский дождь. (День чтения — в сентябре. День образования — в ноябре, Олимпиада — в мае. На эту тему сочиняли даже детские стишки.) Было еще темно, и Джордж одевался при стенном свете. Родители его волновались гораздо больше, чем он сам. Отец Джорджа был дипломированным трубопрокладчиком и работал на Земле, чего втайне стыдился, хотя все понимали, что большая часть каждого поколения неизбежно должна остаться на Земле. Сама Земля нуждалась в фермерах, шахтерах и даже в инженерах. Для работы на других планетах требовались только самые последние модели высококвалифицированных специалистов, и из восьми миллиардов земного населения туда ежегодно отправлялось всего лишь несколько миллионов человек. Естественно, не каждый житель Земли мог попасть в их число. Но каждый мог надеяться, что по крайней мере кому-нибудь из его детей доведется работать на другой планете, и Плейтен старший, конечно, не был исключением. Он видел (как, впрочем, видели и совершенно посторонние люди), что Джордж отличается незаурядными способностями и большой сообразительностью. Значит, его ждет блестящая будущность, тем более, что он единственный ребенок в семье. Если Джордж не попадет на другую планету, то его родителям придется возложить все надежды на внуков, а когда-то еще у них появятся внуки! Сам по себе День чтения, конечно, мало что значил, но в то же время только он мог показать хоть что-нибудь до наступления того, другого, знаменательного дня. Когда дети возвращались домой, все родители Земли внимательно слушали, как они читают, стараясь уловить особенную беглость, чтобы истолковать ее как счастливое предзнаменование. Почти в любой семье подрастал такой многообещающий ребенок, на которого со Дня чтения возлагались огромные надежды только потому, что он легко справлялся с трехсложными словами. Джордж смутно сознавал, отчего так волнуются его родители, и в то дождливое утро его безмятежный детский покой смущал только страх, что радостное выражение на лице отца может угаснуть, когда он вернется домой и покажет, как он научился читать. Детей собрали в просторном зале городского Дома образования. В этот месяц во всех уголках Земли в миллионах местных Домов образования собирались такие же группы детей. Серые стены и напряженность, с которой держались дети, стеснявшиеся непривычной нарядной одежды, нагнали на Джорджа тоску. Он инстинктивно поступил так же, как другие: отыскав кучку ребят, живших с ним на одном этаже, он присоединился к ним. Тревельян, мальчик из соседней квартиры, все еще разгуливал в длинных локонах, а от маленьких бачков и жидких рыжеватых усов, которые ему предстояло отрастить, едва он станет к этому физиологически способен, его отделяли еще многие годы. Тревельян (для которого Джордж тогда был еще Джорджи) воскликнул: — Ага! Струсил, струсил! — Вот и нет! — возразил Джордж и затем доверительно сообщил: — А папа с мамой положили печатный лист на мою тумбочку и, когда я вернусь домой, я прочту им все до последнего словечка. Вот! (В тот момент наибольшее мучение Джорджу причиняли его собственные руки, которые он не знал куда девать. Ему строго-настрого приказали не чесать голову, не тереть уши, не ковырять в носу и не засовывать руки в карманы. Так что же ему было с ними делать?) Зато Тревельян как ни в чем не бывало сунул руки в карманы и заявил: — А вот мой папа ничуточки не беспокоится. Тревельян старший почти семь лет работал металлургом на Динарии, и, хотя теперь он вышел на пенсию и жил опять на Земле, соседи смотрели на него снизу вверх. Возвращение на Землю не очень поощрялось из-за проблемы перенаселенности, но все же кое-кому удавалось вернуться. Прежде всего, жизнь на Земле была дешевле, и пенсия, мизерная в условиях Дипории, на Земле выглядела весьма солидно. Кроме того, некоторым людям особенно приятно демонстрировать свои успехи именно перед друзьями детства и знакомыми, а не перед всей остальной Вселенной. Свое возвращение Тревельян старший объяснил еще и тем, что, останься он на Дипории, там пришлось бы остаться и его детям, а Дипория имела сообщение только с Землей. Живя же на Земле, его дети смогут в будущем попасть на любой из миров, даже на Новию. Коротышка Тревельян рано усвоил эту истину. Еще до Дня чтения он беззаботно верил, что в конце концов будет жить на Новия, и говорил об этом как о деле решенном. Джордж, подавленный мыслью о будущем величии Тревельяна и сознанием собственного ничтожества, немедленно в целях самозащиты перешел в наступление. — Мой папа тоже не беспокоится. Ему просто хочется послушать, как я читаю! Ведь он знает, что читать я буду очень хорошо. А твой отец просто не хочет тебя слушать: он знает, что у тебя ничего не выйдет. — Нет, выйдет! А чтение — это ерунда. Когда я буду жить на Новик, я найму людей, чтобы они мне читали. — Потому что сам ты читать не научишься! Потому что ты дурак! — А как же я тогда попаду на Новик)? И Джордж, окончательно выведенный из себя, посягнул на основу основ: — А кто это тебе сказал, что ты попадешь на Новик)? Никуда ты не попадешь. Вот! Коротышка Тревельян покраснел. — Ну, уж трубопрокладчиком, как твой папаша, я не буду! — Возьми назад, что сказал, дурак) — Сам возьми! Они были готовы броситься друг на друга. Драться им, правда, совсем не хотелось, но возможность заняться чем-то привычным в этом чужом месте сама по себе была уже облегчением. А к тому же Джордж сжал кулаки и встал в боксерскую стойку, так что мучительная проблема — куда девать руки — временно разрешилась. Остальные дети возбужденно обступили их. Но тут же все кончилось: по залу внезапно разнесся усиленный громкоговорителями женский голос — и сразу наступила тишина. Джордж разжал кулаки и забыл о Тревельяне. — Дети, — произнес голос, — сейчас мы будем называть ваши фамилии. Тот, кто услышит свою фамилию, должен тут же подойти к одному из служителей, которые стоят у стен. Вы видите их? Они одеты в красную форму, и вы легко их заметите. Девочки пойдут направо, мальчики — налево. А теперь посмотрите, какой человек в красном стоит к вам ближе всего… Джордж сразу же увидел своего служителя и стал ждать, когда его вы зовут. Он еще побыл посвящен в тайну алфавита, и к тому времени, когда дошла очередь до его фамилии, уже начал волноваться. Толпа детей редела, ручейками растекаясь к красным фигурам. Когда наконец было произнесено имя "Джордж Плейтен", он испытал невыразимое облегчение и упоительную радость: его уже вызвали, а Коротишку — нет! Уходя, Джордж бросил ему через плечо: — Ага, Коротышка! А может, ты им вовсе и не нужен? Но его приподнятое настроение быстро улетучилось. Его поставили рядом с незнакомыми детьми и всех повели по коридорам. Они испуганно переглядывались, но заговорить никто не осмеливался, и слышалось только сопение да иногда сдавленный шепот; "Не толкайся!" и "Эй, ты, поосторожней!" Им раздали картонные карточки и велели их не терять. Джордж стал с любопытством рассматривать свою карточку. Он увидел маленькие черные значки разной формы. Он знал, что это называется печатными буквами, но был не в состоянии представить себе, как из них получаются слова. Его и еще четверых мальчиков отвели в отдельную комнату и велели им раздеться. Они быстро сбросили свою новую одежду и стояли теперь голые и маленькие, дрожа скорее от волнения, чем от холода. Лаборанты быстро, по очереди ощупывали и исследовали их с помощью каких-то странных инструментов, кололи им пальцы, чтобы взять кровь для анализа, а потом каждый брал карточки и черной палочкой торопливо выводил на них аккуратные ряды каких-то значков. Джордж Пристально вглядывался в эти новые значки, но они оставались такими же непонятными, как и старые. Затем детям велели одеться. Они сели на маленькие стулья и снова стали ждать. Их опять начали вызывать по фамилиям, и Джорджа Плейтена вызвали третьим. Он вошел в большую комнату, заполненную страшными аппаратами с множеством кнопок и прозрачных панелей. В самом Центре комнаты стоял письменный стол, за которым, устремив взгляд на кипу лежавших перед ним бумаг, сидел какой-то мужчина. — Джордж Плейтен? — спросил он. — Да, сэр, — дрожащим шепотом ответил Джордж, который в результате длительного ожидания и бесконечных переходов из комнаты в комнату начал волноваться. Он уже мечтал о том, чтобы все это поскорее кончилось. Человек за письменным столом сказал: — Меня зовут доктор Ллойд. Как ты себя чувствуешь, Джордж? Произнося эту фразу, доктор не поднял головы. Казалось, он повторял эти слова так часто, что ему уже не нужно было смотреть на того, к кому он обращался. — Хорошо. — Ты боишься, Джордж? — Н-нет, сэр, — ответил Джордж, и даже от него самого не укрылось, как испуганно прозвучал его голос. — Вот и прекрасно, — произнес доктор, — Ты же знаешь, что бояться нечего. Ну-ка, Джордж, посмотрим! На твоей карточке написано, что твоего отца зовут Питер и что по профессии он дипломированный трубопрокладчик. Имя твоей матери Эми, и она дипломированный специалист по домоведению. Правильно? — Д-да, сэр. — А ты родился 13 февраля и год назад перенес инфекционное заболевание уха. Так? — Да, сэр. — А ты знаешь, откуда мне это известно? — Я думаю, все это есть на карточке. — Совершенно верно, — доктор в первый раз взглянул на Джорджа и улыбнулся, показав ровные зубы. На вид он был гораздо моложе отца Джорджа — и Джордж несколько успокоился. Доктор протянул ему карточку. — Ты знаешь, что означают эти значки? И хотя Джорджу было отлично известно, что этого он не знает, от неожиданности он взглянул на карточку с таким вниманием, словно по велению судьбы внезапно научился читать. Но значки по-прежнему оставались непонятными, и он вернул карточку доктору. — Нет, сэр. — А почему? У Джорджа вдруг мелькнуло подозрение: а не сошел ли с ума этот док тор? Разве он этого не знает сам? — Потому что я не умею читать, сэр. — А тебе хотелось бы научиться читать? — Да, сэр. — А зачем, Джордж?! Джордж в недоумении вытаращил глаза. Никто никогда не задавал ему такого вопроса, и он растерялся. — Я не знаю, сэр, — запинаясь, произнес он. — Печатная информация будет руководить тобой всю твою жизнь. Даже после Дня образования тебе предстоит узнать еще очень многое. И эти знания ты будешь получать из таких вот карточек, из книг, с телевизионных экранов. Печатные тексты расскажут тебе столько полезного и интересного, что не уметь читать было бы так же ужасно, как быть слепым. Тебе это понятно? — Да, сэр. — Ты боишься, Джордж? — Нет, сэр. — Отлично. Теперь я объясню тебе, с чего мы начнем. Я приложу вот эти провода к твоему лбу над уголками глаз. Они приклеятся к коже, но не причинят тебе никакой боли. Потом я включу аппарат и раздастся жужжание. Оно покажется тебе непривычным, и, возможно, тебе будет немного щекотно, но это тоже совершенно безболезненно. Впрочем, если тебе все-таки станет больно, ты мне скажешь, и я тут же выключу аппарат. Но больно не будет. Ну, как, договорились? Судорожно глотнув, Джордж кивнул. — Ты готов? Джордж снова кивнул. С закрытыми глазами он ждал, пока доктор готовил аппаратуру. Родители не раз рассказывали ему про все это. Они тоже говорили, что ему не будет больно. Но зато ребята постарше, которым исполнилось десять, а то и двенадцать лет, всегда дразнили ожидавших своего Дня чтения восьмилеток и кричали: "Берегитесь иглы!" А другие, отозвав малыша в какой-нибудь укромный уголок, по секрету сообщали: "Они разрежут тебе голову вот таким большущим ножом с крючком на конце" — и добавляли множество жутких подробностей. Джордж никогда не принимал это за чистую монету, но тем не менее по ночам его мучили кошмары. И теперь, испытывая непередаваемый ужас, он закрыл глаза. Он не почувствовал прикосновения проводов к вискам. Жужжание доносилось откуда-то издалека, и его заглушал звук стучавшей в ушах крови, такой гулкий, словно все происходило в большой пустой пещере. Джордж рискнул медленно открыть глаза. Доктор стоял к нему спиной. Из одного аппарата ползла узкая лента бумаги, на которой виднелась волнистая фиолетовая линия. Доктор отрывал кусочки этой ленты и вкладывал их в прорезь другой машины. Он снова и снова повторял это, и каждый раз машина выбрасывала небольшой кусочек пленки, который доктор внимательно рассматривал. Наконец, он повернулся к Джорджу, как-то странно нахмурив брови. Жужжание прекратилось. — Уже все? — прошептал Джордж. — Да, — не переставая хмуриться, произнес доктор. — И я уже умею читать? — Джордж не чувствовал в себе никаких изменений. — Что? — переспросил доктор, и на его губах мелькнула неожиданная улыбка. — Все идет, как надо, Джордж. Читать ты будешь через пятнадцать минут. А теперь мы воспользуемся другой машиной, и это уже будет немного дольше. Я закрою тебе всю голову, и, когда я включу аппарат, ты на некоторое время перестанешь видеть и слышать, но тебе не будет больно. На всякий случай я дам тебе в руку выключатель. Если ты все-таки почувствуешь боль, нажми вот эту маленькую кнопку, и все прекратится. Хорошо? Позже Джорджу довелось услышать, что это был не настоящий выключатель и его давали ребенку только для того, чтобы он чувствовал себя спокойнее. Однако он не знал твердо, так ли это, поскольку сам кнопки не нажимал. Ему надели на голову большой шлем обтекаемой формы, выложенный изнутри резиной. Три-четыре небольшие выпуклости присосались к его черепу, но он ощутил лишь легкое давление, которое тут же исчезло. Боли не было. Откуда-то глухо донесся голос доктора: — Ну, как, Джордж, все в порядке? И тогда, без всякого предупреждения, его как будто окутал толстый слой войлока. Он перестал ощущать собственное тело, исчезли чувства, весь мир, вся Вселенная. Остался лишь он сам и доносившийся из бездонных глубин небытия голос, который что-то шептал ему… шептал… шептал… Он напряженно старался услышать и понять хоть что-нибудь, но между ним и шепотом лежал толстый войлок. Потом с него сняли шлем. Яркий свет ударил ему в глаза, а голос доктора отдавался в ушах барабанной дробью. — Вот твоя карточка, Джордж. Скажи, что на ней написано? Джордж снова взглянул на карточку — и вскрикнул. Значки обрели смысл! Они слагались в слова, которые он понимал так отчетливо, будто кто-то подсказывал их ему на ухо. Он был уверен, что именно слышал их. — Так что же на ней написано, Джордж? — На ней написано… написано… "Плейтен Джордж. Родился 13 февраля 64 92 года, родители Питер и Эми Плейтен, место…" — от волнения он не мог продолжать. — Ты умеешь читать, Джордж, — сказал доктор. — Все уже позади. — И я никогда не разучусь? Никогда? — Ну конечно же, нет. — Доктор наклонился и серьезно пожал ему руку. — А сейчас тебя отправят домой. Прошел не один день, прежде чем Джордж освоился со своей новой, замечательной способностью. Он так бегло читал отцу вслух, что Плейтен старший не смог сдержать слез умиления и поспешил поделиться этой радостной новостью с родственниками. Джордж бродил по городу, читая все попадавшиеся ему по пути надписи, и не переставал удивляться тому, что было время, когда он их не понимал. Он пытался вспомнить, что это такое — не уметь читать, и не мог. Ему казалось, будто он всегда умел читать. Всегда. К восемнадцати годам Джордж превратился в смуглого юношу среднего роста, но благодаря худобе он выглядел выше, чем был на самом деле. А коренастый, широкоплечий Тревельян, который был ниже его разве что на дюйм, по-прежнему выглядел настоящим коротышкой. Однако за последний год он стал очень самолюбив и никому не позволял безнаказанно употреблять это прозвище. Впрочем, настоящее имя нравилось ему еще меньше, и его называли просто Тревельяном или каким-нибудь прилично звучавшим сокращением фамилии. А чтобы еще более подчеркнуть свое возмужание, он упорно отращивал баки и жесткие, как щетина, усики. Сейчас он вспотел от волнения, и Джордж, к тому времени тоже сменивший картавое "Джооджи" на односложное гортанное "Джордж", глядел на него, посмеиваясь. Они находились в том же огромном зале, где их однажды уже собирали десять лет назад (и куда они с тех пор ни разу не заходили). Казалось, внезапно воплотилось в действительность туманное сновидение из далекого прошлого. В первые минуты Джордж был очень удивлен, обнаружив, что все здесь как будто стало меньше и теснее, но потом он сообразил, что это вырос он сам. Собралось их здесь меньше, чем в тот, первый раз, и одни юноши. Для девушек был назначен другой день. — Не понимаю, почему нас заставляют ждать так долго, — вполголоса сказал Тревельян. — Обычная волокита, — заметил Джордж. — Вез нее не обойдешься. — И откуда в тебе это идиотское спокойствие? — раздраженно поинтересовался Тревельян. — А мне не из-за чего волноваться. — Послушать тебя, так уши вянут! Надеюсь, ты станешь дипломированным возчиком навоза, вот тогда-то я на тебя погляжу. — Он окинул толпу угрюмым, тревожным взглядом. Джордж тоже посмотрел по сторонам. На этот раз система была иной, чем в День чтения. Все шло гораздо медленнее, а инструкции были розданы сразу в печатном виде — значительное преимущество перед устными инструкциями еще не умеющим читать детям. Фамилии "Плейтен" и "Тревельян" по-прежнему стояли в конце списка, но теперь они уже знали, в чем дело. Юноши один за другим выходили из проверочных комнат. Нахмурившись и явно испытывая неловкость, они забирали свою одежду и вещи и отправлялись узнавать результаты. Каждого окружала с каждым разом все более редевшая кучка тех, кто еще ждал своей очереди. "Ну, как?", "Очень трудно было?", "Как по-твоему, что тебе дали?", "Чувствуешь разницу?" — раздавалось со всех сторон. Ответы были туманными и уклончивыми. Джордж, напрягая всю волю, держался в стороне. Такие разговоры — лучший способ вывести человека из равновесия. Все единогласно утверждали, что больше всего шансов у тех, кто сохраняет спокойствие. Но, несмотря ни на что, он чувствовал, как у него постепенно холодеют руки. Забавно, как с годами приходят новые заботы. Например, высококвалифицированные специалисты отправляются работать на другие планеты только с женами (или мужьями). Ведь на всех планетах необходимо поддерживать правильное соотношение числа мужчин и женщин. А какая девушка откажется выйти за человека, которого посылают на планету класса А? У Джорджа не было на примете никакой определенной девушки, да он и не интересовался ими. Еще не время. Вот когда его мечта осуществится и он получит право добавлять к своему имени слова "дипломированный программист", вот тогда он, как султан в гареме, сможет выбрать любую. Эта мысль взволновала его, и он постарался тут же выкинуть ее из головы. Необходимо сохранять спокойствие. — Что же это все-таки может значить? — пробормотал Тревельян. — Сначала тебе советуют сохранять спокойствие и хладнокровие, а потом тебя ставят в такое вот положение — тут только и сохранять спокойствие! — Может быть, это нарочно? Чтобы с самого начала отделить мужчин от мальчиков? Легче, легче, Трев! — Заткнись! Наконец вызвали Джорджа, но не по радио, как в тот раз, — его фамилия вспыхнула на световом табло. Джордж помахал Тревельяну рукой. — Держись, Трев! Не волнуйся. Когда он входил в проверочную комнату, он был счастлив. Да, счастлив! — Джордж Плейтен? — спросил человек, сидевший за столом. На миг в сознании Джорджа с необыкновенной четкостью возник образ другого человека, который десять лет назад задал такой же вопрос, и ему вдруг показалось, что перед ним тот же доктор, а он, Джордж, переступив порог, снова превратился в восьмилетнего мальчугана. Сидевший за столом поднял голову — его лицо конечно, не имело ничего общего с образом, всплывшим из глубин памяти Джорджа. У этого нос был картошкой, волосы жидкие и спутанные, а под подбородком висела складка, словно прежде он был очень толстым, а потом вдруг сразу похудел. — Ну? — раздраженно произнес он. Джордж очнулся. — Да, я Джодж Плейтен, сэр. — Так и говорите. Я — доктор Зэкери Антонелли. Сейчас мы с вами познакомимся поближе. Он пристально, по-совиному, разглядывал на свет маленькие кусочки пленки. Джордж внутренне содрогнулся. Он смутно вспомнил, что тот, другой доктор (он забыл, как его звали) тоже рассматривал такую же пленку. Неужели это та самая? Тот хмурился, а этотвзглянул на него сейчас так, как будто его что-то рассердило. Джордж уже не чувствовал себя счастливым. Доктор Антонелли раскрыл довольно пухлую папку и осторожно отложил в сторону пленки. — Тут сказано, что вы хотите стать программистом вычислительных машин. — Да, доктор. — Вы не передумали? — Нет, сэр. — Это очень ответственная и сложная профессия. Вы уверены, что она вам по силам? — Да, сэр. — Большинство людей, еще не получивших образования, не называют никакой конкретной профессии. Видимо, они боятся повредить себе. — Наверное так, сэр. — А вас это не пугает? — Я полагаю, что лучше быть откровенным, сэр. Доктор Антонелли кивнул, но выражение его лица осталось прежним. — Почему вы хотите стать программистом? — Как вы только что сказали, сэр, это ответственная и сложная профессия. Программисты выполняют важную и интересную работу. Мне она нравится, и я думаю, что справлюсь с ней. Доктор Антонелли отодвинул папку и кисло взглянул на Джорджа. — Откуда вы знаете, что она вам понравится? Вы, наверное, думаете, что вас тут же подхватит какая-нибудь планета класса А? "Он пробует запугать меня, — с тревогой подумал Джордж. — Спокойно, Джордж, говори правду". — Мне кажется, что у программиста на это большие шансы, — произнес он, — но, даже если бы меня оставили на Земле, работа эта мне все равно нравилась бы, я знаю. ("Во всяком случае, это так и я не лгу", — подумал Джордж.) — Пусть так, но откуда вы это знаете? Вопрос был задан таким тоном, словно на него нельзя было ответить разумно, и Джордж еле сдержал улыбку. У него-то имелся ответ! — Я читал о программировании, сэр, — сказал он. — Что? На лице доктора отразилось неподдельное изумление, и это доставило Джорджу удовольствие. — Я читал о программировании, сэр, — повторил он. — Я купил книгу на эту тему и изучал ее. — Книгу, предназначенную для дипломированных программистов? — Да, сэр. — Но ведь вы могли не понять то, что там написано. — Да, вначале. Но я достал другие книги по математике и электронике и разобрался в них, насколько мог. Я, конечно, знаю не так уж много, но все-таки достаточно, чтобы понять, что мне нравится эта профессия и что я могу быть программистом. (Даже его родители ничего не знали о тайнике, где он хранил эти книги, и не догадывались, почему он проводит так много времени в своей комнате и почему не высыпается.) Доктор оттянул пальцами дряблую складку под подбородком. — А зачем вы это делали, друг мой? — Мне хотелось проверить, действительно ли эта профессия интересна. — Но ведь вам известно, что это не имеет ни малейшего значения. Как бы вас ни привлекала та или иная профессия, вы не получите ее, если физическое устройство вашего мозга делает вас более пригодным для занятий иного рода. Вам ведь это известно? — Мне говорили об этом, — осторожно ответил Джордж. — Так поверьте, что это правда. Джордж промолчал. — Или вы думаете, что изучение какого-нибудь предмета перестроит мозговые клетки в нужном направлении? А еще одна теорийка рекомендует беременной женщине чаще слушать прекрасную музыку, если она хочет, чтобы ребенок стал композитором. Вы, значит, верите в это? Джордж покраснел. Конечно, он думал и об этом. Он полагал, что, постоянно упражняя свой интеллект в избранной области, он получит таким образом дополнительное преимущество. Его уверенность в значительной мере объяснялась именно этим. — Я никогда… — начал было он, но не нашел, что сказать. — Ну, так это неверно, молодой человек! К моменту рождения ваш мозг уже окончательно сформирован. Он может быть изменен ударом, достаточно сильным, чтобы повредить его клетки, или разрывом кровеносного сосуда, или опухолью, или тяжелым инфекционным заболеванием — в любом случае обязательно к худшему. Но повлиять на строение мозга, упорно о чем-то думая, попросту невозможно. — Он задумчиво посмотрел на Джорджа и добавил: Кто вам посоветовал делать это? Окончательно расстроившись, Джордж судорожно глотнул и ответил: — Никто, доктор. Это моя собственная идея. — А кто знал об этих ваших занятиях? — Никто. Доктор, я не хотел ничего плохого. — Кто сказал, что это плохо? Бесполезно, только и всего. А почему вы скрывали свои занятия? — Я… я думал, что надо мной будут смеяться. (Он вдруг вспомнил о недавнем споре с Тревельяном. Очень осторожно, как будто эта мысль только зародилась в самом отдаленном уголке его сознания, Джордж высказал предположение, что, пожалуй, можно кое-чему научиться, черпая знания, так сказать, вручную, постепенно и понемногу. Тревельян оглушительно расхохотался: "Джордж, не хватает еще, чтобы ты начал дубить кожу для своих башмаков и сам ткать материю для своих рубашек". И тогда он обрадовался, что сумел хорошо сохранить свою тайну.) Погрузившись в мрачное раздумье, доктор Антонелли перекладывал с места на место кусочки пленки, которые рассматривал в начале их разговора. Наконец он произнес: — Займемся-ка вашим анализом. Так мы ничего не добьемся. К вискам Джорджа приложили провода. Раздалось жужжание, и снова в его мозгу возникло ясное воспоминание о том, что происходило с ним в этом здании десять лет назад. Руки Джорджа были липкими от пота, его сердце отчаянно колотилось. Зачем, зачем он сказал доктору, что тайком читает книги? Всему виной было его проклятое тщеславие. Ему захотелось щегольнуть своей предприимчивостью и самостоятельностью, а вместо этого он продемонстрировал свое суеверие и невежество и восстановил доктора против себя. (Он чувствовал, что Антонелли возненавидел его за самодовольную развязность.) А теперь он довел себя до такого возбуждения, что анализатор, конечно, покажет полную бессмыслицу. Он не заметил, когда именно с его висков сняли провода. Он только вдруг осознал, что доктор стоит перед ним и задумчиво смотрит на него. А проводов уже не было. Отчаянным усилием воли Джордж взял себя в руки. Он полностью распростился с мечтой стать программистом. За каких-нибудь десять минут она окончательно рассеялась. — Наверно, нет? — уныло спросил он. — Что нет? — Из меня не выйдет программиста? Доктор потер нос и сказал: — Одевайтесь, заберите свои вещи и идите в комнату 3.5-С. Там вас будут ждать ваши бумаги и мое заключение. — Разве я уже получил образование? — изумленно спросил Джордж. — Мне казалось, что это только… Доктор Антонелли внимательно посмотрел на письменный стол. — Вам все объяснят. Делайте, как я сказал. Джордж почувствовал смятение. Что от него утаивают? Он годится только для профессии дипломированного чернорабочего, и его хотят подготовить, заставить смириться с этой судьбой? Он сразу полностью уверовал в правильность своей догадки, и ему пришлось напрячь все силы, чтобы не закричать. Спотыкаясь, он побрел к своему месту в зале. Тревельяна там не было, и, если бы Джордж в тот момент был способен осмысленно воспринимать окружающее, он обрадовался бы этому обстоятельству. В зале вообще уже почти никого не осталось, а те немногие, которые, судя по их виду, как будто намеревались его порасспросить, были слишком измучены ожиданием своей очереди в конце алфавита, чтобы выдержать его свирепый, полный злобной ненависти взгляд. По какому праву они будут квалифицированными специалистами, а он — чернорабочим? Чернорабочим! Он был в этом уверен. Служитель в красной форме повел его по коридорам, полным деловой суеты, мимо комнат, в каждой из которых помещалась та или иная группа специалистов — где два, а где пять человек: механики-мотористы, инженеры-строители, агрономы… Существовали сотни различных профессий, и значительная их часть будет представлена в этом году по крайней мере одним или двумя жителями его родного городка. В эту минуту Джордж ненавидел их всех: статистиков, бухгалтеров, тех, кто поважнее, и тех, кто помельче. Он ненавидел их за то, что они уже получили свои знания и им была ясна их дальнейшая судьба, а его самого, все еще не обученного, ждала какая-то новая волокита. Наконец он добрался до двери с номером 3.5-С. Его ввели в пустую комнату и оставили одного. На какой-то миг он воспрянул духом. Ведь, если бы эта комната предназначалась для чернорабочих, тут уже сидело бы много его сверстников. Дверь в невысокой, в половику человеческого роста, перегородке скользнула в паз, и в комнату вошел пожилой седовласый мужчина. Он улыбнулся, показав ровные, явно фальшивые зубы, однако на его румяном лице не было морщин, а голос сохранил звучность. — Добрый вечер, Джордж, — сказал он. — Я вижу, что на этот раз к нам в сектор попал только один из вас. — Только один? — с недоумением повторил Джордж. — Ну, по всей Земле таких, как вы, наберется несколько тысяч. Да, тысяч. Вы не одиноки. Джордж почувствовал раздражение. — Я ничего не понимаю, сэр, — сказал он. — Какова моя классификация? Что происходит? — Полегче, друг мой. Ничего страшного. Это могло бы случиться с кем угодно. — Он протянул руку, и Джордж, машинально взяв ее, почувствовал крепкое пожатие. — Садитесь. Меня зовут Сэм Элленфорд. Джордж нетерпеливо кивнул. — Я хочу знать, в чем дело, сэр. — Естественно. Во-первых, Джордж, вы не можете быть программистом. Я думаю, что вы и сами об этом догадались. — Да, — с горечью согласился Джордж. — Но кем же я тогда буду? — Это очень трудно объяснить, Джордж. — Он помолчал и затем отчетливо произнес: Никем. — Что?! — Никем! — Что это значит? Почему вы не можете дать мне профессию? — Мы тут бессильны, Джордж, у нас нет выбора. За нас решает устройство вашего мозга. Лицо Джорджа стало землистым, глаза выпучились. — Мой мозг никуда не годится? — Да как сказать. Но в отношении профессиональной классификации — можете считать, что он действительно не годится. — Но почему? Элленфорд пожал плечами. — Вам, конечно, известно, как осуществляется на Земле программа образования. Практически любой человек может усвоить любые знания, но каждый индивидуальный мозг лучше подходит для одних видов знаний, чем для других. Мы стараемся по возможности сочетать устройство мозга с соответствующими знаниями в пределах квоты на специалистов каждой профессии. Джордж кивнул. — Да, я знаю. — Но иногда, Джордж, нам попадается молодой человек, чей интеллект не подходит для наложения на него каких бы то ни было знаний. — Другими словами, я не способен получить образование? — Вот именно. — Но это безумие. Ведь я умен. Я могу понять… — Он беспомощно оглянулся, как бы отыскивая какую-нибудь возможность доказать, что его мозг работает нормально. — Вы неправильно — меня поняли, — очень серьезно произнес Элленфорд. — Вы умны. Об этом вопроса не встает. Более того, ваш интеллект даже выше среднего. К сожалению, это не имеет никакого отношения к тому, подходит ли он для наложения знаний. Вообще сюда почти всегда попадают умные люди. — Вы хотите сказать, что я не могу стать даже дипломированным черно рабочим? — пролепетал Джордж. Внезапно ему показалось, что даже это лучше, чем открывшаяся перед ним пустота. — Что особенного нужно знать, чтобы быть чернорабочим? — Вы недооцениваете чернорабочих, молодой человек. Существует множество разновидностей этой профессии, и каждая из этих разновидностей имеет свой комплекс довольно сложных знаний. Вы думаете, не требуется никакого искусства, чтобы правильно поднимать тяжести? Кроме того, для профессии чернорабочего мы должны отбирать не только подходящий тип мозга, но и подходящий тип тела. Вы не годитесь для этого, Джордж. Если бы вы стали чернорабочим, вас хватило бы ненадолго. Джордж знал, что не обладает крепким телосложением. — Но я никогда не слышал ни об одном человеке без профессии, — возразил он. — Таких людей немного, — ответил Элленфорд. — И мы оберегаем их. — Оберегаете? — Джордж почувствовал, как в его душе растут смятение и страх. — Вы находитесь под опекой планеты, Джордж. С того момента, как вы вошли в эту дверь, мы приступили к своим обязанностям. — И он улыбнулся. Это была добрая улыбка. Джорджу она показалась улыбкой собственника, улыбкой взрослого, обращенной к беспомощному ребенку. — Значит, я попаду в тюрьму? — спросил он. — Ни в коем случае. Вы просто будете жить с себе подобными. "С себе подобными!" Эти слова громом отдались в ушах Джорджа. — Вам нужны особые условия. Мы позаботимся о вас, — сказал Элленфорд. К своему ужасу, Джордж вдруг залился слезами. Элленфорд отошел в другой конец комнаты и, как бы задумавшись о чем-то, отвернулся. Джордж напрягал все силы, и судорожные рыдания сменились всхлипываниями, а потом ему удалось подавить и их. Он думал о своем отце, о матери, о друзьях, о Тревельяне, о своем позоре… — Я же научился читать! — возмущенно сказал он. — Каждый нормальный человек может научиться этому. Нам никогда не приходилось сталкиваться с исключениями. Но именно на этом этапе мы обнаруживаем… э… э… исключения. Когда вы научились читать, Джордж, мы узнали об особенностях вашего мышления. Дежурный врач уже тогда сообщил о некотором его своеобразии. — Неужели вы не можете попробовать дать мне образование? Ведь вы даже не пытались сделать это. Весь риск я возьму на себя. — Закон запрещает нам это, Джордж. Но, послушайте, все будет хорошо. Вашим родителям мы представим дело в таком свете, что это не огорчит их. А там, куда вас поместят, вы будете пользоваться некоторыми привилегиями. Мы дадим вам книги, и вы сможете изучать все, что пожелаете. — Собирать знания по зернышку, — горько произнес Джордж. — И к концу жизни я буду знать как раз достаточно, чтобы стать дипломированным младшим клерком в отделе скрепок. — Однако, если не ошибаюсь, вы уже пробовали учиться по книгам. Джордж замер. Его мозг пронзила страшная догадка. — Так вот оно что… — Что? — Этот ваш Антонелли! Он хочет погубить меня! — Нет, Джордж, вы ошибаетесь. — Не разуверяйте меня. — Джорджа охватила безумная ярость. — Этот поганый ублюдок решил расквитаться со мной за то, что я оказался для него слишком умным. Я читал книги и пытался подготовиться к профессии программиста. Ладно, какого отступного вы хотите? Деньги? Так вы их не получите. Я ухожу, и когда я объявлю об этом… Он перешел на крик. Элленфорд покачал головой и нажал кнопку. В комнату на цыпочках вошли двое мужчин и с двух сторон приблизились к Джорджу. Они прижали его руки к бокам, и один из них поднес к локтевой впадине его правой руки воздушный шприц. Снотворное проникло в вену и подействовало почти мгновенно. Крики Джорджа тут же оборвались, голова поникла, колени подогнулись, и только служители, поддерживавшие его с обеих сторон, не дали ему, спящему, рухнуть на пол. Как и было обещано, о Джордже позаботились. Его окружили вниманием и были к нему неизменно добры — Джорджу казалось, что он сам точно так же обращался бы с больным котенком. Ему сказали, что он должен взять себя в руки и найти для себя какой-нибудь интерес в жизни. Потом ему объяснили, что большинство тех, кто попадает сюда, вначале всегда предается отчаянию и что со временем у него это пройдет. Но он даже не слышал их. Джорджа посетил сам доктор Элленфорд и рассказал, что его родителям сообщили, будто он получил особое назначение. — А они знают?.. — пробормотал Джордж. Элленфорд поспешил успокоить его: — Мы не вдавались в подробности. Первое время Джордж отказывался есть и ему вводили питание внутривенно. От него спрятали острые предметы и приставили к нему охрану. В его комнате поселился Хали Омани, и флегматичность нигерийца подействовала на Джорджа успокаивающе. Однажды, снедаемый отчаянной скукой, Джордж попросил какую-нибудь книгу. Смани, который сам постоянно что-то читал, поднял голову и широко улыбнулся. Не желая доставлять им удовольствия, Джордж чуть было не взял назад свою просьбу, но потом подумал: "А не все ли равно?" Он не уточнил, о чем именно хочет читать, и Смани принес ему книгу по химии. Текст был напечатан крупными буквами, составлен из коротких слов и пояснялся множеством картинок. Это была книга для подростков, и Джордж с яростью швырнул ее об стену. Таким он будет всегда. На всю жизнь он останется подростком, человеком, не получившим образования, и для него будут писать специальные книги. Изнывая от ненависти, он лежал в кровати и глядел в потолок, однако через час, угрюмо насупившись, встал, поднял книгу и принялся за чтение. Через неделю он кончил ее и попросил другую. — А первую отнести обратно? — спросил Омани. Джордж нахмурился. Кое-чего он не понял, но он еще не настолько потерял чувство собственного достоинства, чтобы сознаться в этом. — Впрочем, пусть остается, — сказал Смани. — Книги предназначены для того, чтобы их читали и перечитывали. Это произошло в тот самый день, когда он наконец принял приглашение Смани посмотреть заведение, в котором они находились. Он плелся за нигерийцем, бросая вокруг быстрые злобные взгляды. О да, это место не было тюрьмой! Ни запертых дверей, ни стен, ни охраны. Оно напоминало тюрьму только тем, что его обитатели не могли его покинуть. И все-таки было приятно увидеть десятки подобных себе людей. Ведь слишком легко могло показаться, что во всем мире только ты один так… искалечен. — А сколько здесь живет человек? — пробормотал он. — Двести пять, Джордж, и это не единственное в мире заведение такого рода. Их тысячи. Где бы он ни проходил, люди поворачивались в его сторону и провожали его глазами — и в гимнастическом зале, и на теннисных кортах, и в библиотеке. (Никогда в жизни он не представлял себе, что может существовать такое количество книг; ими были битком — именно битком — набиты длинные полки.) Все с любопытством рассматривали его, а он бросал в ответ яростные взгляды. Уж они-то ничем не лучше его, как же они смеют глазеть на него, словно на какую-нибудь диковинку! Всем им, по-видимому, было лет двадцать — двадцать пять. — А что происходит с теми, кто постарше? — неожиданно спросил Джордж. — Здесь живут только более молодые, — ответил Омани, а затем, словно вдруг поняв скрытый смысл вопроса Джорджа, укоризненно покачал головой и добавил: Их не уничтожают, если ты это имеешь в виду. Для старшего возраста существуют другие приюты. — А впрочем, мне наплевать… — пробормотал Джордж, почувствовав, что проявил к этому слишком большой интерес и ему угрожает опасность сдаться. — Но почему? Когда ты станешь старше, тебя переведут в приют, предназначенный для лиц обоего пола. Это почему-то удивило Джорджа. — И женщин тоже? — Конечно. Неужели ты считаешь, что женщины не подвержены этому? И Джордж поймал себя на том, что испытывает такой интерес и волнение, каких не замечал в себе с того дня, когда… Он заставил себя не думать об этом. Омани остановился на пороге комнаты, где стояли небольшая телевизионная установка и настольная счетная машина. Перед экраном сидели пять-шесть человек. — Классная комната, — пояснил. Омани. — А что это такое? — спросил Джордж. — Эти юноши получают образование, — ответил Омани. — Но не обычным способом, — быстро добавил он. — То есть они получают знания по капле? — Да. Именно так учились в старину. С тех пор как он появился в приюте, ему все время твердили об этом. Но что толку? Предположим, было время, когда человечество не знало диатермической печи. Разве из этого следует, что он должен довольствоваться сырым мясом в мире, где все остальные едят его вареным или жареным? — Что толку от этого крохоборства? — спросил он. — Нужно же чем-то занять время, Джордж, а кроме того, им интересно. — А какая им от этого польза? — Они чувствуют себя счастливее. Джордж размышлял над этим, даже ложась спать. На другой день он буркнул, обращаясь к Смани: — Ты покажешь мне класс, где я смогу узнать что-нибудь о программировании? — Ну конечно, — охотно согласился Омани. Дело продвигалось медленно, и это возмущало Джорджа. Почему кто-то снова и снова объясняет одно и то же? Почему он читает и перечитывает какой-нибудь абзац, а потом, глядя на математическую формулу, не сразу ее понимает? Ведь людям за стенами приюта все это дается в один присест, Несколько раз он бросал занятия. Однажды он не посещал классов целую неделю. Но всегда возвращался обратно. Дежурный наставник, который советовал им, что читать, вел телевизионные сеансы и даже объяснял трудные места и понятия, казалось, не замечал его поведения. В конце концов Джорджу поручили постоянную работу в парке, а кроме того, когда наступала его очередь, он занимался уборкой и помогал на кухне. Ему представили это как шаг вперед, но им не удалось его провести. Ведь тут можно было бы завести куда больше всяческих бытовых приборов, но юношам нарочно давали работу, чтобы создать для них иллюзию полезного существования. Только его, Джорджа, им провести не удалось. Им даже платили небольшое жалованье. Эти деньги они могли тратить на кое-какие дополнительные блага из числа указанных в списке либо откладывать их для сомнительного использования в столь же сомнительной старости. Джордж держал свои деньги в открытой жестянке, стоявшей на полке стенного шкафа. Он не имел ни малейшего представления, сколько там накопилось. Ему это было совершенно безразлично. Он ни с кем по-настоящему не подружился, хотя к этому времени уже вежливо здоровался с обитателями приюта. Он даже перестал (вернее, почти перестал) терзать себя мыслями о роковой ошибке, из-за которой попал сюда. По целым неделям ему уже не снился Антонелли, его толстый нос и дряблая шея, его злобная усмешка, с которой он заталкивал Джорджа в раскаленный зыбучий песок и держал его там до тех пор, пока тот не просыпался с криком, встречая участливый взгляд склонившегося над ним Смани. Как-то раз в снежный февральский день Смани сказал ему: — Просто удивительно, как ты приспособился. Но это было в феврале, точнее, тринадцатого февраля, в день его рождения. Пришел март, за ним апрель, а когда уже не за горами был май, Джордж понял, что ничуть не приспособился. Год назад он не заметил мая. Тогда, впав в апатию и, потеряв цель в жизни, он целыми днями валялся в постели. Но этот май был иным. Джордж знал, что повсюду на Земле вскоре начнется Олимпиада и молодые люди будут состязаться друг с другом в профессиональном искусстве, борясь за места на новых планетах. На всей Земле будет праздничная атмосфера, волнение, нетерпеливое ожидание последних новостей о результатах состязаний. Прибудут важные агенты-вербовщики с далеких планет. Победители будут увенчаны славой, но и потерпевшие поражение найдут чем утешиться. Сколько было об этом написано книг! Как он сам мальчишкой из года в год увлеченно следил за олимпийскими состязаниями! И сколько с этим было связано его личных планов… Джордж Плейтен сказал с плохо скрытой тоской в голосе: — Завтра первое мая. Начало Олимпиады! И это вызвало его первую ссору с Смани, так что тот, сурово отчеканивая каждое слово, произнес полное название заведения, где находился Джордж. Пристально глядя на Джорджа, Смани сказал раздельно: — Приют для слабоумных. Джордж Плейтен покраснел. Для слабоумных! Он в отчаянии отогнал эту мысль и глухо сказал: — Я ухожу. Он сказал это не думая, и смысл этих слов достиг его сознания, лишь когда они уже сорвались с языка. Смани, который снова принялся за чтение, поднял голову. — Что ты сказал? Но теперь Джордж понимал, что говорит. — Я ухожу! — яростно повторил он. — Что за нелепость! Садись, Джордж, и успокойся. — Ну, нет! Сколько раз повторять тебе, что со мной попросту расправились. Я не понравился этому доктору, Антонелли, а все эти мелкие бюрократишки любят покуражиться. Стоит только с ними не поладить, и они одним росчерком пера на какой-нибудь карточке стирают тебя в порошок. — Ты опять за старое? — Да, и не отступлю, пока все не выяснится. Я доберусь до Антонелли и выжму из него правду. — Джордж тяжело дышал, его била нервная дрожь. Наступал месяц Олимпиады, и он не мог допустить, чтобы этот месяц прошел для него безрезультатно. Если он сейчас ничего не предпримет, он окончательно капитулирует и погибнет навсегда. Омани спустил ноги с кровати и встал. Он был почти шести футов ростом, и выражение лица придавало ему сходство с озабоченным сенбернаром. Он обнял Джорджа за плечи. — Если я обидел тебя… Джордж сбросил его руку. — Ты просто сказал то, что считаешь правдой, а я докажу, что это ложь, вот и все. А почему бы мне не уйти? Дверь открыта, замков здесь никаких нет. Никто никогда не говорил, что мне запрещено выходить. Я просто возьму и уйду. — Допустим. Но куда ты отправишься? — В ближайший аэропорт, а оттуда — в ближайший большой город, где проводится какая-нибудь Олимпиада. У меня есть деньги. — Он схватил жестянку, в которую складывал свой заработок. Несколько монет со звоном упало на пол. — Этого тебе, возможно, хватит на неделю. А потом? — К этому времени я все улажу. — К этому времени ты приползешь обратно, — сказал Смани с силой, — и тебе придется начинать все сначала. Ты сошел с ума, Джордж. — Только что ты назвал меня слабоумным. — Ну, извини меня. Останься, хорошо? — Ты что, попытаешься удержать меня? Смани сжал толстые губы. — Нет, не попытаюсь. Это твое личное дело. Если ты образумишься только после того, как столкнешься с внешним миром и вернешься сюда с окровавленной физиономией, так уходи… Да, уходи! Джордж уже стоял в дверях. Он оглянулся через плечо. — Я ухожу. Он вернулся, чтобы взять свой карманный несессер. — Надеюсь, ты ничего не имеешь против, если я заберу кое-что из моих вещей? Омани пожал плечами. Он опять улегся в постель и с безразличным видом погрузился в чтение. Джордж снова помедлил на пороге, но Омани даже не взглянул в его сторону. Скрипнув зубами, Джордж повернулся, быстро зашагал по безлюдному коридору и вышел в окутанный ночной мглой парк. Он ждал, что его задержат еще в парке, но его никто не остановил. Он зашел в ночную закусочную, чтобы спросить дорогу к аэропорту, и думал, что хозяин тут же вызовет полицию, но этого не случилось. Джордж вызвал скиммер, и водитель повез его в аэропорт, не задав ни одного вопроса. Однако все это его не радовало. Когда он прибыл в аэропорт, на душе у него было на редкость скверно. Прежде он как-то не задумывался над тем, что его ожидает во внешнем мире. И вот он оказался среди людей, обладающих профессиями. В закусочной над кассой была укреплена пластмассовая пластинка с именем хозяина. Такой-то, дипломированный повар. У человека, управляющего скиммером, были права дипломированного водителя. Джордж остро почувствовал незаконченность своей фамилии и из-за этого ощущал себя как будто голым, даже хуже — ему казалось, что с него содрали кожу. Но он не поймал на себе ни одного подозрительного взгляда. Никто не остановил его, не потребовал у него профессионального удостоверения. "Кому придет в голову, что есть люди без профессии?" — с горечью подумал Джордж. Он купил билет до Сан-Франциско на стратоплан, улетавший в 3 часа ночи. Другие стратопланы в крупные центры Олимпиады вылетали только утром, а Джордж боялся ждать. Даже и теперь он устроился в самом укромном уголке зала ожидания и стал высматривать полицейских. Но они не явились. В Сан-Франциско он прилетел еще до полудня, и городской шум обрушился на него, подобно удару. Он никогда еще не видел такого большого города, а за последние полтора года привык к тишине и спокойствию. Да и к тому же это был месяц Олимпиады. Когда Джордж вдруг сообразил, что именно этим объясняются особый шум, возбуждение и суматоха, он почти забыл о собственном отчаянном положении. Для удобства приезжающих пассажиров в аэропорту были установлены Олимпийские стенды, перед которыми собирались большие толпы. Для каждой основной профессии был отведен особый стенд, на котором значился адрес того Олимпийского зала, где в данный день проводилось состязание по этой профессии, затем перечислялись его участники с указанием места их рождения и называлась планета-заказчик (если таковая была). Все полностью соответствовало традициям. Джордж достаточно часто читал в газетах описания Олимпийских состязаний, видел их по телевизору и даже однажды присутствовал на небольшой Олимпиаде дипломированных мясников в главном городе своего округа. Даже это состязание, не имевшее никакого отношения к другим мирам (на нем, конечно, не присутствовало ни одного представителя иной планеты), привлекло множество зрителей. Отчасти это объяснялось просто самим фактом состязания, отчасти — местным патриотизмом (о, что творилось, когда среди участников оказывался земляк, пусть даже совершенно незнакомый, но за которого можно было болеть!) и, конечно, до некоторой степени — возможностью заключать пари. Бороться с этим было невозможно. Джордж убедился, что подойти поближе к стенду не так-то просто. Он поймал себя на том, что как-то по-новому смотрит на оживленные лица вокруг. Ведь было же время, когда эти люди сами участвовали в Олимпиадах. А чего они достигли? Ничего! Если бы они были победителями, то не сидели бы на Земле, а находились бы где-нибудь далеко в глубинах Галактики. Кем бы они ни были, их профессии с самого начала сделали их добычей Земли. Или, если у них были высокоспециализированные профессии, они стали добычей Земли из-за собственной бездарности. Теперь эти неудачники, собравшись здесь, взвешивали шансы новых, молодых участников состязаний. Стервятники! Как страстно он желал, чтобы они прикидывали сейчас его шансы! Он машинально шел мимо стендов, держась у самого края толпы. В стратоплане он позавтракал, и ему не хотелось есть. Зато ему было страшно. Он находился в большом городе в самый разгар суматохи, которая сопутствует началу Олимпийских состязаний. Это, конечно, для него выгодно. Город наводнен приезжими. Никто не станет расспрашивать Джорджа. Никому не будет до него никакого дела. "Никому, даже приюту", — с болью подумал Джордж. Там за ним ухаживали, как за больным котенком, но если больной котенок возьмет да убежит? Что поделаешь — тем хуже для него! А теперь, добравшись до Сан-Франциско, что он предпримет? На этот вопрос у него не было ответа. Обратиться к кому-нибудь? К кому именно? Как? Он не знал даже, где ему остановиться. Деньги, которые у него остались, казались жалкими грошами. Он вдруг со стыдом прикинул, не лучше ли будет вернуться в приют. Ведь можно пойти в полицию… Но тут же яростно замотал головой, словно споря с реальным противником. Его взгляд упал на слово "Металлурги", которое ярко светилось на одном из стендов. Рядом, помельче — "Цветные металлы". А под длинным списком фамилий завивались прихотливые буквы: "Заказчик — Новия". На Джорджа нахлынули мучительные воспоминания: его спор с Тревельяном, когда он был так уверен, что станет программистом, так уверен, что программист намного выше металлурга, так уверен, что идет по правильному пути, так уверен в своих способностях… И вот он расхвастался перед этим мелочным, мстительным Антонелли! Он же был так уверен в себе, когда его вызвали, и он еще постарался ободрить нервничавшего Тревельяна. В себе-то он был уверен! У Джорджа вырвался всхлипывающий вздох. Какой-то человек обернулся и, взглянув на него, поспешил дальше. Мимо торопливо проходили люди, поминутно толкая его то в одну, то в другую сторону, а он не мог оторвать изумленных глаз от стенда. Ведь стенд словно откликнулся на его мысли! Он настойчиво думал о Тревельяне, и на мгновение ему показалось, что на стенде в ответ обязательно возникнет слово "Тревельян". Но там и в самом деле значился Тревельян. Арманд Тревельян (имя, которое так ненавидел Коротышка, ярко светилось на стенде для всеобщего обозрения), а рядом — название их родного города. Да и к тому же Трев всегда мечтал о Новии, стремился на Новию, ни о чем не думал, кроме Новии. А заказчиком этого состязания была Новия. Значит, это действительно Трев, старина Трев! Почти машинально он запомнил адрес зала, где проводилось состязание, и стал в очередь на скиммер. "Трев-таки добился своего! — вдруг угрюмо подумал он. — Он хотел стать металлургом и стал!" Джорджу стало холодно и одиноко, как никогда. У входа в зал стояла очередь. Очевидно, Олимпиада металлургов обещала захватывающую и напряженную борьбу. По крайней мере так утверждала горевшая высоко в небе надпись, и так же, казалось, думали теснившиеся у входа люди. Джордж решил, что, судя по цвету неба, день выдался дождливый, но над всем Сан-Франциско, от залива до океана, натянули прозрачный защитный купол. Это, конечно, стоило немалых денег, но, когда дело касается удобства представителей других миров, все расходы оправдываются. А на Олимпиаду их съедется сюда немало. Они швыряют деньги направо и налево, а за каждого нанятого специалиста планета-заказчик платила не только Земле, но и местным властям. Так что город, в котором представители других планет проводили месяц Олимпиады с удовольствием, внакладе не оставался. Сан-Франциско знал, что делает. Джордж так глубоко задумался, что вздрогнул, когда его плеча мягко коснулась чья-то рука и вежливый голос произнес: — Вы стоите в очереди, молодой человек? Очередь продвинулась, и теперь Джордж наконец обнаружил, что перед ним образовалось пустое пространство. Он поспешно шагнул вперед и пробормотал: — Извините, сэр. Два пальца осторожно взяли его за локоть. Он испуганно оглянулся. Стоявший за ним человек весело кивнул. В его волосах пробивалась обильная седина, а под пиджаком он носил старомодный свитер, застегивавшийся спереди на пуговицы. — Я не хотел вас обидеть, — сказал он. — Ничего. — Вот и хорошо! — Он, казалось, был расположен благодушно поболтать. — Я подумал, что вы, может быть, случайно остановились тут; задержавшись из-за очереди, и что вы, может быть… — Кто? — резко спросил Джордж. — Участник состязания, конечно. Вы же очень молоды. Джордж отвернулся. Он был не в настроении для благодушной болтовни и испытывал злость ко всем любителям совать нос в чужие дела. Его внезапно осенила новая мысль. Не разыскивают ли его? Вдруг сюда уже сообщили его приметы или прислали фотографию? Вдруг этот Седой позади него ищет предлога получше рассмотреть его лицо? Надо бы все-таки узнать последние известия. Задрав голову, Джордж взглянул на движущуюся полосу заголовков и кратких сообщений, которые бежали по одной из секций прозрачного купола, непривычно тусклые на сером фоне затянутого облаками предвечернего неба. Но тут же решил, что это бесполезно, и отвернулся. Для этих сообщений его персона слишком ничтожна. Во время Олимпиады только одни новости заслуживают упоминания: количество очков, набранных победителями, и призы, завоеванные континентами, нациями и городами. И так будет продолжаться до конца месяца. Очки будут подсчитываться на душу населения, и каждый город будет изыскивать способ подсчета, который дал бы ему возможность занять почетное место. Его собственный город однажды занял третье место на Олимпиаде электротехников, третье место во всем штате! В ратуше до сих пор висит мемориальная доска, увековечившая это событие. Джордж втянул голову в плечи и засунул руки в карманы, но тут же решил, что так скорее привлечет к себе внимание. Он расслабил мышцы и попытался принять безразличный вид, но от страха не избавился. Теперь он находился уже в вестибюле, и до сих пор на его плечо не опустилась властная рука. Наконец он вошел в зал и постарался пробраться как можно дальше вперед. Вдруг он заметил рядом Седого и опять почувствовал страх. Он быстро отвел взгляд и попытался внушить себе, что это вполне естественно. В конце концов. Седой стоял в очереди прямо за ним. К тому же Седой, поглядев на него с приветливой вопросительной улыбкой, отвернулся. Олимпиада вот-вот должна была начаться. Джордж приподнялся, высматривая Тревельяна, и на время забыл обо всем остальном. Зал был не очень велик и имел форму классического вытянутого овала. Зрители располагались на двух галереях, опоясывавших зал, а участники состязания — внизу, в длинном и узком углублении. Приборы были уже установлены, а на табло над каждым рабочим местом пока светились только фамилии и номера состязающихся. Сами участники были на сцене. Одни читали, другие разговаривали. Кто-то внимательно разглядывал свои ногти. (Хороший тон требовал, чтобы они проявляли полное равнодушие к предстоящему испытанию, пока не будет подан сигнал к началу состязания.) Джордж просмотрел программу, которая появилась из ручки его кресла, когда он нажал кнопку, и отыскал фамилию Тревельяна. Она значилась под номером двенадцать, и, к огорчению Джорджа, место его приятеля находи лось в другом конце зала. Он нашел участника под двенадцатым номером: тот, засунув руки в карманы, стоял спиной к своему прибору и смотрел на галереи, словно пересчитывал зрителей, но лица его Джордж не видел. И все-таки он сразу узнал Трева. Джордж откинулся в кресле. Добьется ли Трев успеха? Из чувства долга он желал ему самых лучших результатов, однако в глубине его души нарастал бунт. Он, Джордж, человек без профессии, сидит здесь простым зрителем, а Тревельян, дипломированный металлург, специалист по цветным металлам, участвует в Олимпиаде. Джордж не знал, выступал ли Тревельян олимпийским соискателем в первый год после получения профессии. Такие смельчаки находились: либо человек был очень уверен в себе, либо очень торопился. В этом крылся определенный риск. Как ни эффективен был процесс обучения, год, проведенный на Земле после получения образования ("чтобы смазать неразработавшиеся знания", согласно поговорке), обеспечивал более высокие результаты. Если Тревельян выступает в состязаниях вторично, он, быть может, не так уж и преуспел. Джордж со стыдом заметил, что эта мысль ему скорее приятна. Он поглядел по сторонам. Почти все места были заняты. Олимпиада соберет много зрителей, а значит, участники будут больше нервничать, а может быть, и работать с большей энергией — в зависимости от характера. "Но почему это называется Олимпиадой?" — подумал он вдруг в недоумении. Он не знал. Почему хлеб называют хлебом? В детстве он как-то спросил отца: — Папа, а что такое Олимпиада? И отец ответил: — Олимпиада — значит состязание. — А когда мы с Коротышкой боремся, это тоже Олимпиада? — поинтересовался Джордж. — Нет, — ответил Плейтен старший. — Олимпиада — это особенное состязание. Не задавай глупых вопросов. Когда получишь образование, узнаешь все, что тебе положено знать. Джордж глубоко вздохнул, вернулся к действительности и уселся поглубже в кресло. Все, что тебе положено знать! Странно, как хорошо он помнит эти слова! "Когда ты получишь образование…" Никто никогда не говорил: "Если ты получишь образование…" Теперь ему казалось, что он всегда задавал глупые вопросы. Как будто его разум инстинктивно предвидел свою неспособность к образованию и придумывал всяческие расспросы, чтобы хоть по обрывкам собрать побольше знаний. А в приюте поощряли его любознательность, проповедуя то же, к чему инстинктивно стремился его разум. Единственный открытый ему путь! Внезапно он выпрямился. Черт возьми, что это он? Чуть было не попался на их удочку! Неужели он сдастся только потому, что там перед ним Трев, получивший образование и участвующий в Олимпиаде? Нет, он не слабоумный! Нет!! И, словно в ответ на этот мысленный вопль протеста, зрители вокруг зашумели. Все встали. В ложу, расположенную в самом центре длинной дуги овала, входили люди, одетые в цвета планеты Новия, и на главном табло над их головами вспыхнуло слово "Новия". Новия была планетой класса А с большим населением и высокоразвитой цивилизацией, быть может самой лучшей во всей Галактике. Каждый землянин мечтал когда-нибудь поселиться в таком мире, как Новия, или, если ему это не удастся, увидеть там своих детей. Джордж вспомнил, с какой настойчивостью стремился к Новии Тревельян, и вот теперь он состязается за право уехать туда. Лампы над головами зрителей погасли, потухли и стены. Поток яркого света хлынул вниз, туда, где находились участники состязания. Джордж снова попытался рассмотреть Тревельяна. Но тот был слишком далеко. — Уважаемые новианские заказчики, уважаемые дамы и господа! — раздался отчетливый, хорошо поставленный голос диктора. — Сейчас начнутся Олимпийские состязания металлургов, специалистов по цветным металлам. В состязании принимают участие… С добросовестной аккуратностью диктор прочитал список, приведенный в программе. Фамилии, названия городов, откуда прибыли участники, год получения образования. Зрители встречали каждое имя аплодисментами, и самые громкие доставались на долю жителей Сан-Франциско. Когда очередь дошла до Тревельяна, Джордж неожиданно для самого себя бешено завопил и замахал руками. Но еще больше его удивило то, что сидевший рядом с ним седой мужчина повел себя точно так же. Джордж не мог скрыть своего изумления, и его сосед, наклонившись к нему и напрягая голос, чтобы перекричать шум, произнес: — У меня здесь нет земляков. Я буду болеть за ваш город. Это ваш знакомый? Джордж отодвинулся, насколько мог. — Нет. — Я заметил, что вы все время смотрите в том направлении. Не хотите ли воспользоваться моим биноклем? — Нет, благодарю вас. (Почему этот старый дурак сует нос не в свое дело?) Диктор продолжал сообщать другие официальные сведения, касавшиеся номера состязания, метода хронометрирования и подсчитывания очков. Наконец он дошел до самого главного, и публика замерла, обратившись в слух. — Каждый участник состязания получит по бруску сплава неизвестного для него состава. От него потребуется произвести количественный анализ этого сплава и сообщить все результаты с точностью до четырех десятых процента. Для этого все соревнующиеся будут пользоваться микроспектрографами Бимена, модель ЛЮ-2, каждый из которых в настоящее время неисправен. Зрители одобрительно зашумели. — Каждый участник должен будет определить неисправность своего прибора и ликвидировать ее. Для этого им даны инструменты и запасные детали. Среди них может не оказаться нужной детали, и ее надо будет затребовать. Время, которое займет доставка, вычитается из общего времени,затраченного на выполнение задания. Все участники готовы? На табло над пятым номером вспыхнул тревожный красный сигнал. Номер пять бегом бросился из зала и быстро вернулся. Зрители добродушно рассмеялись. — Все готовы? Табло остались темными. — Есть какие-нибудь вопросы? По-прежнему ничего. — Можете начинать! Разумеется, ни один из зрителей не имел возможности непосредственно определить, как продвигается работа у каждого участника. Некоторое представление об этом могли дать только надписи, вспыхивавшие на табло. Впрочем, это не имело ни малейшего значения. Среди зрителей только металлург, окажись он здесь, мог бы разобраться в сущности состязания. Но важно было, кто победит, кто займет второе, а кто — третье место. Для тех, кто ставил на участников (а этому не могли помешать никакие законы), только это было важно. Все прочее их не интересовало. Джордж следил за состязанием так же жадно, как и остальные, поглядывая то на одного участника, то на другого. Он видел, как вот этот, ловко орудуя каким-то маленьким инструментом, снял крышку со своего микроспектрографа; как тот всматривался в экран аппарата; как спокойно вставлял третий свой брусок сплава в зажим и как четвертый накручивал верньер, причем настолько осторожно, что, казалось, на мгновение застыл в полной неподвижности. Тревельян, как и все остальные участники, был целиком поглощен своей работой. А как идут его дела, Джордж определить не мог. На табло над семнадцатым номером вспыхнула надпись: "Сдвинута фокусная пластинка". Зрители бешено зааплодировали. Семнадцатый номер мог быть прав, но мог, конечно, и ошибиться. В этом случае ему пришлось бы позже исправить свой вывод, потеряв на этом время. А может быть, он не заметил бы ошибки и не сумел бы сделать анализ. Или же, еще хуже, он мог получить совершенно неверные результаты. Неважно. А пока зрители ликовали. Зажглись и другие табло. Джордж не спускал глаз с табло номер двенадцать. Наконец оно тоже засветилось: "Держатель децентрирован. Требуется новый зажим". К Тревельяну подбежал служитель с новой деталью. Если Трев ошибся, это означает бесполезную задержку, а время, потраченное на ожидание детали, не будет вычтено из общего времени. Джордж невольно затаил дыхание. На семнадцатом табло начали появляться светящиеся буквы результата анализа: "Алюминий — 41,2649, магний — 22,1914, медь — 10.1001". И на других табло все чаще вспыхивали цифры. Зрители бесновались. Джордж недоумевал, как участники могли работать в таком бедламе, потом ему пришло в голову, что, может быть, это даже хорошо: ведь первоклассный специалист лучше всего работает в напряженной обстановке. На семнадцатом табло вспыхнула красная рамка, знаменующая окончание работы, и семнадцатый номер поднялся со своего места. Четвертый отстал от него всего лишь на две секунды. Затем кончил еще один и еще. Тревельян продолжал работать, определяя последние компоненты своего сплава. Он поднялся, когда почти все состязающиеся уже стояли. Последним встал пятый номер, и публика приветствовала его ироническими возгласами. Однако это был еще не конец. Заключение жюри, естественно, задерживалось. Время, затраченное на всю операцию, имело определенное значение, но не менее важна была точность результатов. И не все задачи были одинаково трудны. Необходимо было учесть множество факторов. Наконец раздался голос диктора: — Победителем состязания, выполнившим задание за четыре минуты двенадцать секунд, правильно определившим неисправность и получившим правильный результат с точностью до семи десятитысячных процента, является участник под номером… семнадцать, Генрих Антон Шмидт из… Остальное потонуло в бешеном реве. Второе место занял восьмой номер, третье — четвертый, хороший показатель времени которого был испорчен ошибкой в пять сотых процента при определении количественного содержания ниобия. Двенадцатый номер даже не был упомянут, если не считать фразы"… а остальные участники…" Джордж протолкался к служебному выходу и обнаружил, что здесь уже собралось множество людей — плачущие (кто от радости, кто от горя) родственники, репортеры, намеренные взять интервью у победителей, земляки, охотники за автографами, любители рекламы и просто любопытные. Были здесь и девушки, надеявшиеся обратить на себя внимание победителя, который почти наверняка отправится на Новик) (а может быть, и потерпевшего поражение, который нуждается в утешении и имеет деньги, чтобы позволить себе такую роскошь). Джордж остановился в сторонке. Он не увидел ни одного знакомого лица. Сан-Франциско был так далеко от их родного города, что вряд ли Трев приехал сюда в сопровождении близких, которые теперь печально поджидали бы его у двери. Смущенно улыбаясь и кланяясь в ответ на приветствия, появились участники соревнования. Полицейские сдерживали толпу, освобождая им проход. Каждый из набравших большое количество очков увлекал за собой часть людей, подобно магниту, двигающемуся по кучке железных опилок. Когда вышел Тревельян, у входа уже почти никого не было. (Джордж решил, что он долго выжидал этой минуты.) В его сурово сжатых губах была сигарета. Глядя в землю, он повернулся, чтобы уйти. Это было первое напоминание о родном доме за без малого полтора года, которые показались Джорджу в десять раз дольше. И он даже удивился, что Тревельян нисколько не постарел и остался все тем же Тревом, каким он видел его в последний раз. Джордж рванулся вперед. — Трев! Тревельян в изумлении обернулся. Он с недоумением взглянул на Джорджа и сразу же протянул ему руку. — Джордж Плейтен! Вот черт… Появившееся на его лице радостное выражение тут же угасло, а рука опустилась, прежде чем Джордж успел пожать ее. — Ты был там? — Тревельян мотнул головой в сторону зала. — Был. — Чтобы посмотреть на меня? — Да. — Я не слишком блеснул, а? Он бросил сигарету, раздавил ее ногой, глядя в сторону улицы, где медленно рассасывавшаяся толпа окружала скиммеры и уже стояли новые очереди желающих попасть на следующие состязания. — Ну и что? — угрюмо буркнул Тревельян. — Я проиграл всего во второй раз. А после сегодняшнего Новия может катиться ко всем чертям. Есть планеты, которые просто вцепятся в меня… Но послушай-ка, ведь я не видел тебя со Дня образования. Где ты пропадал? Твои родные сказали, что ты уехал по специальному заданию, но ничего не объяснили подробно. И ты ни разу мне не написал. А мог бы. — Да, пожалуй, — неловко произнес Джордж. — Но я пришел сказать, как мне жаль, что сейчас все так обернулось. — Не жалей, — возразил Тревельян. — Я ведь уже говорил тебе, что Новия может убираться к черту… Да я мог бы знать заранее! Все только и говорили, что использован будет прибор Бимена. Никто и не сомневался. А в проклятых лентах, которыми меня зарядили, был предусмотрен спектрограф Хенслера! Кто же теперь пользуется Хенслером? Разве что планеты вроде Гомона, если их вообще можно назвать планетами. Ловко это было подстроено, а? — Но ты ведь можешь подать жалобу в… — Не будь дураком. Мне скажут, что мой мозг лучше всего подходил для Хенслера. Пойди поспорь. Да и вообще мне не везло. Ты заметил, что мне одному пришлось послать за запасной частью? — Но потраченное на это время вычиталось? — Конечно! Только я, когда заметил, что среди запасных частей нет зажима, подумал, что напутал, и не сразу потребовал его. Это-то время не вычиталось! А будь у меня микроспектрограф Хенслера, я бы знал, что не ошибаюсь. Где мне было тягаться с ними? Первое место занял житель Сан-Франциско, и следующие три тоже. А пятое занял парень из Лос-Анджелеса. Они получили образование с лент, которыми снабжают большие города. С самых лучших, которые только есть. Там и спектрограф Бимена и все, что хочешь! Куда же мне было до них! Я отправился в такую даль потому, что только эту Олимпиаду по моей профессии заказала Новия, и с тем же успехом мог бы остаться дома. Я заранее знал, что так получится! Но теперь все. На Новии космос клином не сошелся. Из всех проклятых… Он говорил это не для Джорджа. Он вообще ни к кому не обращался. Джордж понял, что он просто отводит душу. — Если ты заранее знал, что вам дадут спектрограф Бимена, разве ты не мог ознакомиться с ним? — спросил Джордж. — Я же говорю тебе, что его не было в моих лентах. — Ты мог почитать… книги. Тревельян вдруг так пронзительно взглянул на него, что он еле выговорил последнее слово. — Ты что, смеешься? — сказал Тревельян. — Остришь? Неужели ты думаешь, что, прочитав какую-то книгу, я запомню достаточно, чтобы сравняться с теми, кто действительно знает? — Я считал… — А ты попробуй. Попробуй… Кстати, а ты какую получил профессию? — вдруг злобно спросил Тревельян. — Видишь ли… — Ну, выкладывай. Раз уже ты тут передо мной строишь такого умника, давай-ка посмотрим, кем стал ты. Раз ты все еще на Земле, значит, ты не программист и твое специальное задание не такое уж важное. — Послушай, Трев, — сказал Джордж, — я опаздываю на свидание… Он попятился, пытаясь улыбнуться. — Нет, ты не уйдешь. — Тревельян в бешенстве бросился к Джорджу и вцепился в его пиджак. — Отвечай! Почему ты боишься сказать мне? Кто ты такой? Ты мне тычешь в нос мое поражение, а сам? Это у тебя не выйдет, слышишь? Он неистово тряс Джорджа, тот вырывался. Так, отчаянно борясь и чуть не падая, они двигались через зал, но тут Джордж услышал глас Рока — суровый голос полицейского: — Довольно! Довольно! Прекратите это! Сердце Джорджа мучительно сжалось и превратилось в кусок свинца. Сейчас полицейский спросит их имена и потребует удостоверения личности, а у Джорджа нет никаких документов. После первых же вопросов выяснится, что у него нет и профессии. А Тревельян, озлобленный своей неудачей, конечно, поспешит рассказать об этом всем знакомым в родном городке, чтобы утешить собственное уязвленное самолюбие. Этого Джордж не мог вынести. Он вырвался и бросился было бежать, но ему на плечо легла тяжелая рука полицейского. — Эй, постойте. Покажите-ка ваше удостоверение. Тревельян шарил в карманах и говорил отрывисто и зло: — Я Арманд Тревельян, металлург по цветным-металлам. Я участвовал в Олимпиаде. А вот его проверьте хорошенько, сержант. Джордж стоял перед ними, не в силах вымолвить ни слова. Губы его пересохли, горло сжалось. Вдруг раздался еще один голос, спокойный и вежливый: — Можно вас на минутку, сержант? Полицейский шагнул назад. — Что вам угодно, сэр? — Этот молодой человек — мой гость. Что случилось? Джордж оглянулся вне себя от изумления. Это был тот самый седой мужчина, который сидел рядом с ним на Олимпиаде. Седой добродушно кивнул Джорджу. Его гость? Он что, сошел с ума? — Эти двое затеяли драку, сэр, — объяснил полицейский. — Вы предъявляете им какое-нибудь обвинение? Нанесен ущерб? — Нет, сэр. — В таком случае всю ответственность я беру на себя. Он показал полицейскому небольшую карточку, и тот сразу отступил. — Постойте… — возмущенно начал Тревельян, но полицейский свирепо перебил его: — Ну? У вас есть какие-нибудь претензии? — Я только… — Проходите! И вы тоже… Расходитесь, расходитесь! И собравшаяся вокруг толпа начала с неохотой расходиться. Джордж покорно пошел с Седым к скиммеру, но тут решительно остановился. — Благодарю вас, — сказал он, — но ведь я не ваш гость. (Может быть, по нелепой случайности его приняли за кого-то другого?) Но Седой улыбнулся и сказал: — Теперь вы уже мой гость. Разрешите представиться. Я — Ладислас Индженеску, дипломированный историк. — Но… — С вами ничего дурного не случится, уверяю вас. Я ведь просто хотел избавить вас от неприятного разговора с полицейским. — А почему? — Вы хотите знать причину? Ну, ведь мы с вами, так сказать, почетные земляки. Мы же дружно болели за одного человека. А земляки должны держаться друг друга, даже если они только почетные земляки. Не правда ли? И Джордж, не доверяя ни Индженеску, ни самому себе, все-таки вошел в скиммер. Они поднялись в воздух, прежде чем он успел передумать. "Это, наверное, важная птица, — вдруг сообразил он. — Полицейский говорил с ним очень почтительно". Только теперь он вспомнил, что приехал в Сан-Франциско вовсе не ради Тревельяна, а с целью найти достаточно влиятельного человека, который мог бы добиться переоценки его способностей. А вдруг этот Индженеску именно тот, кто ему нужен? И его даже не придется искать! Как знать, не сложилось ли все на редкость удачно… удачно… Но Джордж напрасно убеждал себя. На душе у него было по-прежнему тревожно. Во время недолгого полета на скиммере Индженеску поддерживал разговор, любезно указывая на достопримечательности города и рассказывая о других Олимпиадах, на которых ему доводилось бывать. Джордж слушал его рассеянно, издавал невнятное хмыканье, когда Индженеску замолкал, а сам с волнением следил за направлением полета. Вдруг они поднимутся к отверстию в защитном куполе и покинут город? Но скиммер снижался, и Джордж тихонько вздохнул с облегчением. В городе он чувствовал себя в большей безопасности. Скиммер опустился на крышу какого-то отеля, прямо у верхней двери, и, когда они вышли, Индженеску спросил: — Вы не откажетесь пообедать со мной в моем номере? — С удовольствием, — ответил Джордж и улыбнулся вполне искренне. Время второго завтрака давно прошло, и у него начало сосать под ложечкой. Они ели молча. Наступили сумерки, и автоматически засветились стены. ("Вот уже почти сутки, как я на свободе", — подумал Джордж.) За кофе Индженеску наконец заговорил. — Вы вели себя так, словно подозревали меня в дурных намерениях, сказал он. Джордж покраснел и, поставив чашку, попытался что-то возразить, но его собеседник рассмеялся и покачал головой. — Это так. Я внимательно наблюдал за вами с того момента, как впервые вас увидел, и, мне кажется, теперь я знаю о вас очень многое. Джордж в ужасе приподнялся с места. — Сядьте, — сказал Индженеску. — Я ведь только хочу помочь вам. Джордж сел, но в его голове вихрем неслись мысли. Если старик знал, кто он, то почему он помешал полицейскому? Да и вообще, с какой стати он решил ему помогать? — Вам хочется знать, почему я захотел помочь вам? — спросил Индженеску. — О, не пугайтесь, я не умею читать мысли. Видите ли, просто моя профессия позволяет мне по самой незначительной внешней реакции судить о мыслях человека. Вам это понятно? Джордж отрицательно покачал головой. — Представьте себе, каким я увидел вас, — сказал Индженеску. — Вы стояли в очереди, чтобы посмотреть Олимпиаду, но ваши микрореакции не соответствовали тому, что вы делали. У вас было не то выражение лица, не те движения рук. Отсюда следовало, что у вас какая-то беда, но, что самое интересное, необычная, не лежащая на поверхности. Быть может, вы сами не сознаете, что с вами, решил я. И, не удержавшись, последовал за вами, даже сел рядом. После окончания состязания я опять пошел за вами и подслушал ваш разговор с вашим знакомым. Ну, а уж к этому времени вы превратились в такой интересный объект для изучения — простите, если это звучит бессердечно, — что я просто не мог допустить, чтобы вас забрали в полицию… Скажите же, что вас тревожит? Джордж мучился, не зная, на что решиться. Если это ловушка, то зачем нужно действовать таким окольным путем? Ему же действительно нужна помощь. Ради этого он сюда и приехал. А тут помощь ему прямо предлагают. Пожалуй, именно это его и смущало. Что-то все получается уж очень просто. — Разумеется, то, что вы сообщите мне как социологу, становится профессиональной тайной, — сказал Индженеску. — Вы понимаете, что это значит? — Нет, сэр. — Это значит, что с моей стороны будет бесчестным, если я расскажу о том, что узнаю от вас, с какой бы целью я это ни сделал. Более того, никто не имеет права заставить меня рассказать об этом. — А я думал, вы историк, — подозрительно сказал Джордж. — Это верно. — Но вы же только сейчас сказали, что вы социолог. Индженеску расхохотался. — Не сердитесь, молодой человек, — извинился он, когда был в состоянии говорить. — Но право же, я смеялся не над вами. Я смеялся над Землей, над тем, какое большое значение она придает точным наукам, и над некоторыми практическими следствиями этого увлечения. Держу пари, что вы можете перечислить все разделы строительной технологии или прикладной механики и в то же время даже не слышали о социологии. — Ну, а что же такое социология? — Социология — это наука, которая занимается изучением человеческого общества и отдельных его ячеек и делится на множество специализированных отраслей, так же как, например, зоология. Так, существуют культурологи, изучающие культуру, ее рост, развитие и упадок. Культура, — добавил он, предупреждая вопрос Джорджа, — это совокупность всех сторон жизни. К культуре относится, например, то, каким путем мы зарабатываем себе на жизнь, от чего получаем удовольствие, во что верим, наши представления о хорошем и плохом и так далее. Вам это понятно? — Кажется, да. — Экономист — не специалист по экономической статистике, а именно экономист — специализируется на изучении того, каким образом культура удовлетворяет физические потребности каждого члена общества. Психолог изучает отдельных членов общества и то влияние, которое это общество на них "Заказывает. Прогнозист планирует будущий путь развития общества, а историк… Это уже по моей части. — Да, сэр? — Историк специализируется на изучении развития нашего общества в прошлом, а также обществ с другими культурами. Джорджу стало интересно. — А разве в прошлом что-то было по-другому? — Еще бы! Тысячу лет назад не было образования, то есть образования, как мы понимаем его теперь. — Знак — произнес Джордж. — Люди учились по книгам, собирая знания по крупицам. — Откуда вы это знаете? — Слыхал, — осторожно ответил Джордж и добавил: А какой смысл думать о том, что происходило в далеком прошлом? Я хочу сказать, что ведь со всем этим уже покончено, не правда ли? — С прошлым никогда не бывает покончено, мой друг. Оно объясняет настоящее. Почему, например, у нас существует именно такая система образования? Джордж беспокойно заерзал. Слишком уж настойчиво его собеседник возвращался к этой теме. — Потому что она самая лучшая, — отрезал он. — Да. Но почему она самая лучшая? Послушайте меня минутку, и я попы таюсь объяснить. А потом вы мне скажете, есть ли смысл в изучении истории. Даже до того, как начались межзвездные полеты… — Он внезапно умолк, заметив на лице Джорджа выражение глубочайшего изумления. — Неужели вы считали, что так было всегда? — Я никогда не задумывался над этим, сэр. — Вполне естественно. Однако четыре-пять тысяч лет назад человечество было приковано к Земле. Но и тогда уже техника достигла высокого уровня развития, а численность населения увеличилась настолько, но любое торможение техники привело бы к массовому голоду и эпидемиям. Для того чтобы уровень техники не снижался и соответствовал росту населения, нужно было готовить все больше инженеров и ученых. Однако по мере развития науки на их обучение требовалось все больше и больше времени. Когда же впервые были открыты способы межпланетных, а затем и межзвездных полетов, эта проблема стала еще острее. Собственно говоря, из-за недостатка специалистов человечество в течение почти полутора тысяч лет не могло по-настоящему колонизировать планеты, находящиеся за пределами Солнечной системы. Перелом наступил, когда был установлен механизм хранения знаний в человеческом мозгу. Как только это было сделано, появилась возможность создать образовательные ленты на основе этого механизма таким образом, чтобы сразу вкладывать в мозг определенное количество, так сказать, готовых знаний. Впрочем, это-то вы знаете. Это позволило выпускать тысячи и миллионы специалистов, и мы смогли приступить к тому, что впоследствии назвали "заполнением Вселенной". Сейчас в Галактике уже существует полторы тысячи населенных планет, и число их будет возрастать до бесконечности. Вы понимаете, что из этого следует? Земля экспортирует образовательные ленты, предназначенные для подготовки специалистов низкой квалификации, и это обеспечивает единство культуры для всей Галактики. Так, например, благодаря лентам чтения мы все говорим на одном языке… Не удивляйтесь. Могут быть и иные языки, и в прошлом люди на них говорили. Их были сотни. Земля, кроме того, экспортирует высококвалифицированных специалистов, и численность ее населения не превышает допустимого уровня. Поскольку при вывозе специалистов соблюдается равновесие полов, они образуют самовоспроизводящиеся ячейки, и это способствует росту населения на тех планетах, где в этом есть необходимость. Более того, за ленты и специалистов платят сырьем, в котором мы очень нуждаемся и от которого зависит наша экономика. Теперь вы поняли, почему наша система образования действительно самая лучшая? — Да, сэр. — И вам легче понять это, зная, что без нее в течение полутора тысяч лет было невозможно колонизировать планеты других солнечных систем? — Да, сэр. — Значит, вы видите, в чем польза истории? — Историк улыбнулся. — А теперь скажите, догадались ли вы, почему я вами интересуюсь? Джордж мгновенно вернулся из пространства и времени назад к действительности. Видимо, Индженеску неспроста завел этот разговор. Вся его лекция была направлена на то, чтобы атаковать его неожиданно. — Почему же? — неуверенно спросил он, снова насторожившись. — Социологи изучают общество, а общество состоит из людей. — Ясно. — Но люди не машины. Специалисты в области точных наук работают с машинами. А машина требует строго определенного количества знаний, и эти специалисты знают о ней все. Более того, все машины данного рода почти одинаковы, так что индивидуальные особенности машины не представляют для них интереса. Но люди… О, они так сложны и так отличаются друг от друга, что социолог никогда не знает о них все или хотя бы значительную часть-того, что можно о них знать. Чтобы не утратить квалификации, он должен постоянно изучать людей, особенно необычные экземпляры. — Вроде меня, — глухо произнес Джордж. — Конечно, называть вас экземпляром невежливо, но вы человек необычный. Вы не стоите того, чтобы вами заняться, и, если вы разрешите мне это, я в свою очередь по мере моих возможностей помогу вам в вашей беде. В мозгу Джорджа кружился смерч. Весь этот разговор о людях и о колонизации, ставшей возможной благодаря образованию… Как будто кто-то разбивал и дробил заскорузлую, спекшуюся корку мыслей. — Дайте мне подумать, — произнес он, зажав руками уши. Потом он опустил руки и сказал историку: — Вы можете оказать мне услугу, сэр? — Если она в моих силах, — любезно ответил историк. — Все, что я говорю в этой комнате, — профессиональная тайна? Вы так сказали. — Так оно и есть. — Тогда устройте мне свидание с каким-нибудь должностным лицом другой планеты, например с… с новианином. Индженеску был, по-видимому, крайне удивлен. — Право же… — Вы можете сделать это, — убежденно произнес Джордж. — Вы ведь важное должностное лицо. Я видел, какой вид был у полицейского, когда вы показали ему свое удостоверение. Если вы откажетесь сделать это, я… я не позволю вам изучать меня. Самому Джорджу эта угроза показалась глупой и бессильной. Однако на Индженеску она, очевидно, произвела большое впечатление. — Ваше условие невыполнимо, — сказал он. — Новианин в месяц Олимпиады… — Ну, хорошо, тогда свяжите меня с каким-нибудь новианином по видеофону, и я сам договорюсь с ним о встрече. — Вы думаете, вам это удастся? — Я в этом уверен. Вот увидите. Индженеску задумчиво посмотрел на Джорджа и протянул руку к видеофону. Джордж ждал, опьяненный новым осмыслением всей проблемы и тем ощущением силы, которое оно давало. Он не может потерпеть неудачу. Не может. Он все-таки станет новианином. Он покинет Землю победителем вопреки Антонелли и всей компании дураков из приюта (он чуть было не расхохотался вслух) для слабоумных. Джордж впился взглядом в засветившийся экран, который должен был распахнуть окно в комнату новиан, окно в перенесенный на Землю уголок Новии. И он добился этого за какие-нибудь сутки! Когда экран прояснился, раздался взрыв смеха, но на нем не появилось ни одного лица, лишь быстро мелькали тени мужчин и женщин. Послышался чей-то голос, отчетливо прозвучавший на фоне общего гомона. — Индженеску? Спрашивает меня? И вот на экране появился он. Новианин. Настоящий новианин. (Джордж ни на секунду не усомнился. В нем было что-то совершенно внеземное, нечто такое, что невозможно было точно определить или хоть на миг спутать с чем-либо иным.) Он был смугл, и его темные волнистые волосы были зачесаны со лба. Он носил танине черные усики и остроконечную бородку, которая только-только закрывала узкий подбородок. Но его щеки были такими гладкими, словно с них навсегда была удалена растительность. Он улыбался. — Ладислас, это уже слишком. Мы не протестуем, чтобы за нами, пока мы на Земле, следили — в разумных пределах, конечно. Но чтение мыслей в условие не входит! — Чтение мыслей, достопочтенный? — Сознайтесь-ка! Вы ведь знали, что я собирался позвонить вам сегодня. Вы знали, что я думал только допить вот эту рюмку. — На экране появилась его рука, и он посмотрел сквозь рюмку, наполненную бледно-сиреневой жидкостью. — К сожалению, я не могу угостить вас. Новианин не видел Джорджа, находившегося вне поля зрения видеофона. И Джордж обрадовался передышке. Ему необходимо было время, чтобы прийти в себя. Он словно превратился в сплошные беспокойные пальцы, которые непрерывно отбивали нервную дробь… Но он все-таки был прав. Он не ошибся. Индженеску действительно занимает важное положение. Новианин называет его по имени. Отлично! Все устраивается наилучшим образом. То, что Джордж потерял из-за Антонелли, он возместит с лихвой, используя Индженеску. И когда-нибудь он, став наконец самостоятельным, вернется на Землю таким же могущественным новианином, как этот, что небрежно шутит с Индженеску, называя его по имени, а сам оставаясь "достопочтенным", — вот тогда он сведет счеты с Антонелли. Он отплатит ему за эти полтора года, и он… Увлекшись этими соблазнительными грезами, он чуть не забыл обо всем на свете, но, внезапно спохватившись, заметил, что перестал следить за происходящим, и вернулся к действительности. — …не убедительно, — говорил новианин. — Новианская цивилизация так же сложна и так же высокоразвита, как цивилизация Земли. Новия — это все-таки не Зестон. И нам приходится прилетать сюда за отдельными специалистами — это же просто смешно! — О, только за новыми моделями, — примирительным тоном сказал Индженеску. — А новые модели не всегда находят применение. На приобретение образовательных лент вы потратили бы столько же, сколько вам пришлось бы заплатить за тысячу специалистов, а откуда вы знаете, что вам будет нужно именно такое количество? Новианин залпом допил свое вино и расхохотался. (Джорджа покоробило легкомыслие новианина. Он смущенно подумал, что тому следовало бы обойтись без этой рюмки и даже без двух или трех предыдущих.) — Это же типичное ханжество, Ладислас, — сказал новианин. — Вы прекрасно знаете, что у нас найдется дело для всех последних моделей специалистов, которые нам удастся достать. Сегодня я раздобыл пять металлургов. — Знаю, — сказал Индженеску. — Я был там. — Следили за мной! Шпионили! — вскричал новианин. — Ну, так слушайте! Эта новая модель металлурга отличается от предыдущих только тем, что умеет обращаться со спектрографом Бимена. Ленты не были модифицированы ни на вот столечко (он показал самый кончик пальца) по сравнению с прошлогодними. Вы выпускаете новые модели только для того, чтобы мы приезжали сюда с протянутой рукой и тратились на их приобретение. — Мы не заставляем вас их приобретать. — О, конечно! Только вы продаете специалистов последней модели на Лондонум, а мы ведь не можем отставать. Вы втянули нас в заколдованный круг, вы лицемерные земляне. Но берегитесь, может быть, где-нибудь есть из него выход. — Его смех прозвучал не слишком естественно и резко оборвался. — От всей души надеюсь, что он существует, — сказал Индженеску. Ну, а позвонил я потому… — Да, конечно, ведь это вы мне позвонили. Что ж, я уже высказал свое мнение. Наверное, в будущем году все равно появится новая модель металлурга, чтобы нам было за что платить. И она будет отличаться от нынешней только умением обращаться с каким-нибудь новым приспособлением для анализа ниобия, а еще через год… Но продолжайте. Почему вы позвонили? — У меня здесь находится один молодой человек, и я бы хотел, чтобы вы с ним побеседовали. — Что? — Видимо, новианина это не слишком обрадовало. — На какую тему? — Не знаю. Он мне не сказал. По правде говоря, он даже не назвал мне ни своего имени, ни профессии. Новианин нахмурился. — Тогда зачем же отнимать у меня время? — Он, по-видимому, не сомневается, что вас заинтересует то, что он собирается сообщить вам. — О, конечно! — И этим вы сделаете одолжение мне, — сказал Индженеску. Новианин пожал плечами. — Давайте его сюда, но предупредите, чтобы он говорил покороче. Индженеску отступил в сторону и шепнул Джорджу: — Называйте его "достопочтенным". Джордж с трудом проглотил слюну. Вот оно! Джордж почувствовал, что весь вспотел. Хотя эта мысль пришла ему в голову совсем недавно, он был убежден в своей правоте. Она возникла во время разговора с Тревельяном, потом под болтовню Индженеску перебродила и оформилась, а теперь слова новианина, казалось, поставили все на свои места. — Достопочтенный, я хочу показать вам выход из заколдованного круга, — начал Джордж, используя метафору новианина. Новианин смерил его взглядом. — Из какого это заколдованного круга? — Вы сами упомянули о нем, достопочтенный. Из того заколдованного круга, в который попадает Новия, когда вы прилетаете на Землю за… за специалистами. (Он не в силах был справиться со своими зубами, которые стучали, но не от страха, а от волнения.) — Вы хотите сказать, что знаете способ, как нам обойтись без земного интеллектуального рынка? Я правильно вас понял? — Да, сэр. Вы можете создать свою собственную систему образования. — Гм. Без лент? — Д-да, достопочтенный. — Индженеску, подойдите, чтобы я видел и вас, — не спуская глаз с Джорджа, позвал новианин. Историк встал за плечом Джорджа. — В чем дело? — спросил новианин. — Не понимаю. — Даю вам слово, достопочтенный, что бы это ни было, молодой человек поступает так по собственной инициативе. Я ему ничего не поручал. Я не имею к этому никакого отношения. — Тогда кем он вам приходится? Почему вы звоните мне по его просьбе? — Я его изучаю, достопочтенный. Он представляет для меня определенную ценность, и я исполняю некоторые его прихоти. — В чем же его ценность? — Это трудно объяснить. Чисто профессиональный момент. Новианин усмехнулся. — Что ж, у каждого своя профессия. Он кивнул невидимому зрителю или зрителям за экраном. — Некий молодой человек, по-видимому, протеже Индженеску, собирается объяснить нам, как получать образование, не пользуясь лентами. Он щелкнул пальцами, и в его руке появилась новая рюмка с бледно-сиреневым напитком. — Ну, говорит, молодой человек. На экране теперь появилось множество лиц. Мужчины и женщины отталкивали друг друга, чтобы поглядеть на Джорджа. На их лицах отражались самые разнообразные оттенки веселья и любопытства. Джордж попытался принять независимый вид. Все они, и новиане, и землянин, каждый по-своему изучали его, словно жука, насажанного на булавку. Индженеску теперь сидел в углу и не спускал с него пристального взгляда. "Какие же вы все идиоты", — напряженно подумал он. Но они должны понять. Он заставит их понять. — Я был сегодня на Олимпиаде металлургов, — сказал он. — Как, и вы тоже? — вежливо спросил новианин. — По-видимому, там присутствовала вся Земля. — Нет, достопочтенный, но я там был. В состязании участвовал мой друг, и ему очень не повезло, потому что вы дали участникам состязания прибор Бимена, а он получил специализацию по Хенслеру, — очевидно, уже устаревшая модель. Вы же сами сказали, что различие очень незначительно. — Джордж показал кончик пальца, повторяя недавний жест своего собеседника. — И мой друг знал заранее, что потребуется знакомство с прибором Бимена. — И что же из этого следует? — Мой друг всю жизнь мечтал попасть на Новию. Он уже знал прибор Хенслера. Он знал, что ему нужно ознакомиться с прибором Бимена, чтобы попасть к вам. А для этого ему следовало усвоить всего лишь несколько дополнительных сведений и, быть может, чуточку попрактиковаться. Если учесть, что на чашу весов была поставлена цель всей его жизни, он мог бы с этим справиться… — А где бы он достал ленту с дополнительной информацией? Или образование здесь, на Земле, превратилось в частное домашнее обучение? Лица на заднем плане расплылись в улыбках, которых, по-видимому, от них и ожидали. — Поэтому-то он и не стал доучиваться, достопочтенный. Он считал, что ему для этого нужна лента. А без нее он и не пытался учиться, как ни заманчива была награда. Он и слышать не хотел, что без ленты можно чему-то научиться. — Да неужели? Так он, пожалуй, даже не захочет летать без скиммера? — Раздался новый взрыв хохота, и новианин слегка улыбнулся. — А он забавен, — сказал он. — Продолжайте. Даю вам еще несколько минут. — Не думайте, что это шутка, — сказал Джордж горячо. — Ленты попросту вредны. Они учат слишком многому и слишком легко. Человек, который получает знания с их помощью, не представляет, как можно учиться по-другому. Он способен заниматься только той профессией, которой его зарядили. А если бы, вместо того чтобы пичкать человека лентами, его заставили с самого начала учиться, так сказать вручную, он привык бы учиться самостоятельно и продолжал бы учиться дальше. Разве это не разумно? А когда эта привычка достаточно укрепится, человеку можно будет прививать небольшое количество знаний с помощью лент, чтобы заполнить пробелы или закрепить кое-какие детали. После этого он сможет учиться дальше самостоятельно. Таким способом вы могли бы научить металлургов, знающих спектрограф Хенслера, пользоваться спектрографом Бимена, и вам не пришлось бы прилетать на Землю за новыми моделями. Новианин кивнул и отхлебнул из рюмки. — А откуда можно получить знания помимо лент? Из межзвездного пространства? — Из книг. Непосредственно изучая приборы. Думая. — Из книг? Как же можно понять книги, не получив образования? — Книги состоят из слов, а большую часть слов можно понять. Специальные же термины могут объяснить специалисты, которых вы уже имеете. — А как быть с чтением? Для этого вы допускаете использование лент? — По-видимому, ими можно пользоваться, хотя не вижу причины, почему нельзя научиться читать и старым способом. По крайней мере частично. — Чтобы с самого начала выработать хорошие привычки? — спросил новианин. — Да, да, — подтвердил Джордж, радуясь, что собеседник уже начал понимать его. — А как быть с математикой? — Это легче всего, сэр… достопочтенный. Математика отличается от других технических дисциплин. Она начинается с некоторых простых принципов и лишь постепенно усложняется. Можно приступить к изучению математики, ничего о ней не зная. Она практически и предназначена для этого. А познакомившись с соответствующими разделами математики, уже нетрудно разобраться в книгах по технике. Особенно если начать с легких. — А разве есть легкие книги? — Безусловно. Но если бы их и не было, специалисты, которых вы уже имеете, могут написать их. Наверное, некоторые из них сумеют выразить свои знания с помощью слов и символов. — Боже мой! — сказал новианин, обращаясь к сгрудившимся вокруг него людям. — У этого чертенка на все есть ответ. — Да, да! — вскричал Джордж. — Спрашивайте! — А сами-то вы пробовали учиться по книгам? Или это только ваша теория? Джордж быстро оглянулся на Индженеску, но историк сохранял полную невозмутимость. Его лицо выражало только легкий интерес. — Да, — сказал Джордж. — И вы считаете, что из этого что-нибудь получается? — Да, достопочтенный, — заверил Джордж. — Возьмите меня с собой на Новию. Я могу составить программу и руководить… — Погодите, у меня есть еще несколько вопросов. Как вы думаете, сколько вам понадобится времени, чтобы стать металлургом, умеющим обращаться со спектрографом Бимена, если предположить, что вы начнете учиться, не имея никаких знаний, и не будете пользоваться образовательными лентами? Джордж заколебался. — Ну… может быть, несколько лет. — Два года? Пять? Десять? — Еще не знаю, достопочтенный. — Итак, на самый главный вопрос у вас не нашлось ответа. Ну, скажем, пять лет. Вас устраивает этот срок? — Думаю, что да. — Отлично. Итак, в течение пяти лет человек изучает металлургию по вашему методу. Вы не можете не согласиться, что все это время он для нас абсолютно бесполезен, но его нужно кормить, обеспечить жильем и платить ему. — Но… — Дайте мне кончить. К тому времени, когда он будет готов и сможет пользоваться спектрографом Бимена, пройдет пять лет. Вам не кажется, что тогда у нас уже появятся усовершенствованные модели этого прибора, с которыми он не сумеет обращаться? — Но ведь к тому времени он станет опытным учеником и усвоение новых деталей будет для него вопросом дней. — По-вашему, это так. Ладно, предположим, что этот ваш друг, например, сумел самостоятельно изучить прибор Бимена; сможет ли сравниться его умение с умением участника состязания, который получил его посредством лент? — Может быть, и нет… — начал Джордж. — То-то же, — сказал новианин. — Погодите, дайте кончить мне. Даже если он знает кое-что хуже, чем тот, другой, в данном случае важно то, что он может учиться дальше. Он сможет придумывать новое, на что не способен ни один человек, получивший образование с лент. У вас будет запас людей, способных к самостоятельному мышлению… — А вы в процессе своей учебы придумали что-нибудь новое? — спросил новианин. — Нет, но ведь я один, и я не так уж долго учился… — Да… Ну-с, дамы и господа, мы достаточно позабавились? — Постойте! — внезапно испугавшись, крикнул Джордж. — Я хочу договориться с вами о личной встрече. Есть вещи, которые я не могу объяснить по видеофону. Ряд деталей… Новианин уже не смотрел на Джорджа. — Индженеску! По-моему, я исполнил вашу просьбу. Право же, завтра у меня очень напряженный день. Всего хорошего. Экран погас. Руки Джорджа взметнулись к экрану в бессмысленной попытке вновь его оживить. — Он не поверил мне! Не поверил! — Да, Джордж, не поверил. Неужели вы серьезно думали, что он поверит? — сказал Индженеску. Но Джордж не слушал. — Почему же? Ведь это правда. Это так для него выгодно. Никакого риска. Только я и еще несколько… Обучение десятка людей в течение даже многих лет обошлось бы дешевле, чем один готовый специалист… Он был пьян! Пьян! Он не был способен понять. Задыхаясь, Джордж оглянулся. — Как мне с ним увидеться? Это необходимо. Все получилось не так, как нужно. Я не должен был говорить с ним по видеофону. Мне нужно время. И чтобы лично. Как мне… — Он откажется принять вас, Джордж, — сказал Индженеску. — А если и согласится, то все равно вам не поверит. — Нет, поверит, уверяю вас. Когда он будет трезв, он… — Джордж повернулся к историку, и глаза его широко раскрылись. — Почему вы называете меня Джорджем? — А разве это не ваше имя? Джордж Плейтен? — Вы знаете, кто я? — Я знаю о вас все. Джордж замер, и только его грудь тяжело вздымалась. — Я хочу помочь вам, Джордж, — сказал Индженеску. — Я уже говорил вам об этом. Я давно изучаю вас и хочу вам помочь. — Мне не нужна помощь! — крикнул Джордж. — Я не слабоумный! Весь мир выжил из ума, но не я! Он стремительно повернулся и бросился к двери. За ней стояли два полицейских, которые его немедленно схватили. Как Джордж ни вырывался, шприц коснулся его шеи под подбородком. И все кончилось. Последнее, что осталось в его памяти, было лицо Индженеску, который с легкой тревогой наблюдал за происходящим. Когда Джордж открыл глаза, он увидел белый потолок. Он помнил, что произошло. Но помнил, как сквозь туман, словно это произошло с кем-то другим. Он смотрел на потолок до тех пор, пока не наполнился его белизной, казалось, освобождавшей его мозг для новых идей, для иных путей мышления. Он не знал, как долго лежал так, прислушиваясь к течению своих мыслей. — Ты проснулся? — раздался чей-то голос. И Джордж впервые услышал свой собственный стон. Неужели он стонал? Он попытался повернуть голову. — Тебе больно, Джордж? — спросил голос. — Смешно, — прошептал Джордж. — Я так хотел покинуть Землю. Я же ничего не понимал. — Ты знаешь, где ты? — Снова в… в приюте. — Джорджу удалось повернуться. Голос принадлежал Омани. — Смешно, как я ничего не понимал, — сказал Джордж. Омани ласково улыбнулся. — Поспи еще… Джордж заснул. И снова проснулся. Сознание его прояснилось. У кровати сидел Омани и читал, но, как только Джордж открыл глаза, он отложил книгу. Джордж с трудом сел. — Привет, — сказал он. — Хочешь есть? — Еще бы! — Джордж с любопытством посмотрел на Омани. — За мной следили, когда я ушел отсюда, так? Омани кивнул. — Ты все время был под наблюдением. Мы считали, что тебе следует побывать у Антонелли, чтобы ты мог дать выход своим агрессивным потребностям. Нам казалось, что другого способа нет. Эмоции тормозили твое развитие. — Я был к нему очень несправедлив, — с легким смущением произнес Джордж. — Теперь это не имеет значения. Когда в аэропорту ты остановился у стенда металлургов, один из наших агентов сообщил нам список участников. Мы с тобой говорили о твоем прошлом достаточно, для того чтобы я мог понять, как подействует на тебя фамилия Тревельяна. Ты спросил, как попасть на эту Олимпиаду. Это могло привести к кризису, на который мы надеялись, и мы послали в зал ЛадисласаИндженеску, чтобы он занялся тобой сам. — Он ведь занимает важный пост в правительстве? — Да. — И вы послали его ко мне. Выходит, что я сам много значу. — Ты действительно много значишь, Джордж. Принесли дымящееся ароматное жаркое. Джордж улыбнулся и откинул простыню, чтобы освободить руки. Смани помог ему поставить поднос на тумбочку. Некоторое время Джордж молча ел. — Я уже один раз ненадолго просыпался, — заметил он. — Знаю, — сказал Смани. — Я был здесь. — Да, я помню. Ты знаешь, все изменилось. Как будто я так устал, что уже не мог больше чувствовать. Я больше не злился. Я мог только думать. Как будто мне дали наркотик, чтобы уничтожить эмоции. — Нет, — сказал Омами. — Это было просто успокоительное. И ты хорошо отдохнул. — Ну, во всяком случае, мне все стало ясно, словно я всегда знал это, но не хотел прислушаться к внутреннему голосу. "Чего я ждал от Новии?" — подумал я. Я хотел отправиться на Новию, чтобы собрать группу юношей, не получивших образования, и учить их по книгам. Я хотел открыть там приют для слабоумных… вроде этого… а на Земле уже есть такие приюты… и много. Смани улыбнулся, сверкнув зубами. — Институт высшего образования — вот как точно называются эти заведения. — Теперь-то я это понимаю, — сказал Джордж, — до того ясно, что только удивляюсь, каким я был слепым. В конце концов, кто изобретает новые модели механизмов, для которых нужны новые модели специалистов? Кто, например, изобрел спектрограф Бимена? По-видимому, человек по имени Бимен. Но он не мог получить образование через зарядку, иначе ему не удалось бы продвинуться вперед. — Совершенно верно. — А кто создает образовательные ленты? Специалисты по производству лент? А кто же тогда создает ленты для их обучения? Специалисты более высокой квалификации? А кто создает ленты… Ты понимаешь, что я хочу сказать. Где-то должен быть конец. Где-то должны быть мужчины и женщины, способные к самостоятельному мышлению. — Ты прав, Джордж. Джордж откинулся на подушки и устремил взгляд в пространство. На какой-то миг в его глазах мелькнула тень былого беспокойства. — Почему мне не сказали об этом с самого начала? — К сожалению, это невозможно, — ответил Смани. — А так мы были бы избавлены от множества хлопот. Мы умеем анализировать интеллект, Джордж, и определять, что вот этот человек может стать приличным архитектором, а тот — хорошим плотником. Но мы не умеем определять, способен ли человек к творческому мышлению. Это слишком тонкая вещь. У нас есть несколько простейших способов, позволяющих распознавать тех, кто, быть может, обладает такого рода талантом. Об этих индивидах сообщают сразу после Дня чтения, как, например, сообщили о тебе. Их приходится примерно один на десять тысяч. В День образования этих людей проверяют снова, и в девяти случаях из десяти оказывается, что произошла ошибка. Тех, кто остается, посылают в такие заведения, как это. — Но почему нельзя сказать людям, что один из… из ста тысяч попадает в такое заведение? — спросил Джордж. — Тогда тем, с кем это случается, было бы легче. — А как же остальные? Те девяносто девять тысяч девятьсот девяносто девять человек, которые никогда не попадут сюда? Нельзя, чтобы все эти люди чувствовали себя неудачниками. Они стремятся получить профессии и получают их. Каждый может прибавить к своему имени слова "дипломированный специалист по тому-то или тому-то". Так или иначе каждый индивид находит свое место в обществе. Это необходимо. — А мы? — спросил Джордж. — Мы, исключения? Один на десять тысяч? — Вам ничего нельзя объяснить. В том-то и дело. Ведь в этом заключается последнее испытание. Даже после отсева в День образования девять человек из десяти, попавших сюда, оказываются не совсем подходящими для творчества, и нет такого прибора, который помог бы нам выделить из этой десятки того единственного, кто нам нужен. Десятый должен доказать это сам. — Каким образом? — Мы помещаем вас сюда, в приют для слабоумных, и тот, кто не желает смириться с этим, и есть человек, которого мы ищем. Быть может, это жестокий метод, но он себя оправдывает. Нельзя же сказать человеку: "Ты можешь творить. Так давай, твори". Гораздо вернее подождать, пока он сам не скажет: "Я могу творить, и я буду творить, хотите вы этого или нет". Есть около десяти тысяч людей, подобных тебе, Джордж, и от них зависит технический прогресс полутора тысяч миров. Мы не можем позволить себе потерять хотя бы одного из них или тратить усилия на того, кто не вполне отвечает необходимым требованиям. Джордж отодвинул пустую тарелку и поднес к губам чашку с кофе. — А как же с теми, которые… не вполне отвечают требованиям? — В конце концов они проходят зарядку и становятся социологами. Индженеску — один из них. Сам я — дипломированный психолог. Мы, так сказать, составляем второй эшелон. Джордж допил кофе. — Мне все еще непонятно одно, — сказал он. — Что же? Джордж сбросил простыню и встал. — Почему состязания называются Олимпиадой?
ЛИКБЕЗ
Основы биотехнологии
Н. А. Кузьмина
ВВЕДЕНИЕ В БИОТЕХНОЛОГИЮ
Биотехнология, или технология биопроцессов, это производственное использование биологических агентов или их систем для получения ценных продуктов и осуществления целевых превращений. Биологические агенты в данном случае — микроорганизмы, растительные и животные клетки, клеточные компоненты: мембраны клеток, рибосомы, митохондрии, хлоропласты, а также биологические макромолекулы (ДНК, РНК, белки — чаще всего ферменты). Биотехнология использует также вирусную ДНК или РНК для переноса чужеродных генов в клетки. Человек использовал биотехнологию многие тысячи лет: люди пекли хлеб, варили пиво, делали сыр, другие молочнокислые продукты, используя различные микроорганизмы, при этом, даже не подозревая об их существовании. Собственно сам термин появился в нашем языке не так давно, вместо него употреблялись слова "промышленная микробиология", "техническая биохимия" и др. Вероятно, древнейшим биотехнологическим процессом было сбраживание с помощью микроорганизмов. В пользу этого свидетельствует описание процесса приготовления пива, обнаруженное в 1981 г. при раскопках Вавилона на дощечке, которая датируется примерно 6-м тысячелетием до н. э. В 3-м тысячелетии до н. э. шумеры изготовляли до двух десятков видов пива. Не менее древними биотехнологическими процессами являются виноделие, хлебопечение, и получение молочнокислых продуктов. В традиционном, классическом, понимании биотехнология — это наука о методах и технологиях производства различных веществ и продуктов с использованием природных биологических объектов и процессов Термин "новая" биотехнология в противоположность "старой" биотехнологии применяют для разделения биопроцессов, использующих методы генной инженерии, новую биопроцессорную технику, и более традиционные формы биопроцессов. Так, обычное производство спирта в процессе брожения — "старая" биотехнология, но использование в этом процессе дрожжей, улучшенных методами генной инженерии с целью увеличения выхода спирта — "новая" биотехнология. Биотехнология как наука является важнейшим разделом современной биологии, которая, как и физика, стала в конце XX в. одним из ведущих приоритетов в мировой науке и экономике. Всплеск исследований по биотехнологии в мировой науке произошел в 80-х годах, когда новые методологические и методические подходы обеспечили переход к эффективному их использованию в науке и практике и возникла реальная возможность извлечь из этого максимальный экономический эффект. По прогнозам, уже в начале 21 века биотехнологические товары будут составлять четверть всей мировой продукции. В нашей стране значительное расширение научно-исследовательских работ и внедрение их результатов в производство также было достигнуто в 80-е годы. В этот период в стране была разработана и активно осуществлялась первая общенациональная программа по биотехнологии, были созданы межведомственные биотехнологические центры, подготовлены квалифицированные кадры специалистов-биотехнологов, организованы биотехнологические лаборатории и кафедры в научно-исследовательских учреждениях и вузах. Однако в дальнейшем внимание к проблемам биотехнологии в стране ослабло, а их финансирование сокращено. В результате развитие биотехнологических исследований и их практическое использование в России замедлилось, что привело к отставанию от мирового уровня, особенно в области генетической инженерии. Что касается более современных биотехнологических процессов, то они основаны на методах рекомбинантных ДНК, а также на использовании иммобилизованных ферментов, клеток или клеточных органелл. Современная биотехнология — это наука о генно-инженерных и клеточных методах и технологиях создания и использования генетически трансформированных биологических объектов для интенсификации производства или получения новых видов продуктов различного назначения. В рамках биотехнологии можно выделить 3 основных части: 1. Промышленная биотехнология, где рассматриваются общие принципы осуществления биотехнологических процессов, происходит знакомство с основными объектами и сферами применения биотехнологии, рядом крупномасштабных промышленных биотехнологических производств, использующих микроорганизмы. 2. Клеточная инженерия. Основная цель этого раздела — знакомство с методами ведения культур клеток и практическим использованием этих объектов. В рамках этого раздела выделяют культивирование растительных клеток и методы культивирования животных клеток, так как подходы к культивированию этих объектов различаются в силу их принципиальных биологических различий. Клеточная биотехнология обеспечила ускоренное получение новых важных форм и линий растений и животных, используемых в селекции на устойчивость, продуктивность и качество; размножение ценных генотипов, получение ценных биологических препаратов пищевого, кормового и медицинского назначения. 3. Генная инженерия. Высшим достижением современной биотехнологии является генетическая трансформация, перенос чужеродных генов и других материальных носителей наследственности в клетки растений, животных и микроорганизмов, получение трансгенных организмов с новыми или усиленными свойствами и признаками. По своим целям и возможностям в перспективе это направление является стратегическим. Оно позволяет решать коренные задачи селекции биологических объектов на устойчивость, высокую продуктивность и качество продукции при оздоровлении экологической обстановки во всех видах производств. Однако для достижения этих целей предстоит преодолеть огромные трудности в повышении эффективности генетической трансформации и, прежде всего, в идентификации генов, создании их банков клонирования, расшифровке механизмов полигенной детерминации признаков и свойств биологических объектов, обеспечении высокой экспрессии генов и создании надежных векторных систем. Уже сегодня во многих лабораториях мира, в том числе и в России, с помощью методов генетической инженерии созданы принципиально новые трансгенные растения, животные и микроорганизмы, получившие коммерческое признание. Современная биотехнология тесно стыкуется с рядом научных дисциплин, осуществляя их практическое применение или же являясь их основным инструментом (рис. 1).

Рис. 1. Связь биотехнологии с другими науками (по В.И.Кефели, 1989)
В молекулярной биологии использование биотехнологических методов позволяет определить структуру генома, понять механизм экспрессии генов, смоделировать клеточные мембраны с целью изучения их функций и т. д. Конструирование нужных генов методами генной и клеточной инженерии позволяет управлять наследственностью и жизнедеятельностью животных, растений и микроорганизмов и создавать организмы с новыми полезными для человека свойствами, ранее не наблюдавшимися в природе. Микробиологическая промышленность в настоящее время использует тысячи штаммов различных микроорганизмов. В большинстве случаев они улучшены путем индуцированного мутагенеза и последующей селекции. Это позволяет вести широкомасштабный синтез различных веществ. Некоторые белки и вторичные метаболиты могут быть получены только путем культивирования клеток эукариот. Растительные клетки могут служить источником ряда соединений — атропин, никотин, алкалоиды, сапонины и др. Клетки животных и человека также продуцируют ряд биологически активным соединений. Например, клетки гипофиза — липотропин, стимулятор расщепления жиров, и соматотропин — гормон, регулирующий рост. Созданы перевиваемые культуры клеток животных, продуцирующие моноклональные антитела, широко применяемые для диагностики заболеваний. В биохимии, микробиологии, цитологии несомненный интерес вызывают методы иммобилизации как ферментов, так и целых клеток микроорганизмов, растений и животных. В ветеринарии широко используются такие биотехнологические методы, как культура клеток и зародышей, овогенез in vitro, искусственное оплодотворение. Все это свидетельствует о том, что биотехнология станет источником не только новых продуктов питания и медицинских препаратов, но и получения энергии и новых химических веществ, а также организмов с заданными свойствами.
Промышленная биотехнология
ОСНОВНЫЕ НАПРАВЛЕНИЯ БИОТЕХНОЛОГИИ
Условно можно выделить следующие основные направления биотехнологии: биотехнология пищевых продуктов, препаратов для сельского хозяйства, препаратов и продуктов для промышленного и бытового использования, лекарственных препаратов, средств диагностики и реактивов, биотехнология также включает выщелачивание и концентрирование металлов, защиту окружающей среды от загрязнения, деградацию токсических отходов и увеличение добычи нефти.
Биоэнергетика Растительный покров Земли составляет более 1800 млрд. т сухого вещества, что энергетически эквивалентно известным запасам энергии полезных ископаемых. Леса составляют около 68 % биомассы суши, травяные экосистемы — примерно 16 %, а возделываемые земли — только 8 %. Для сухого вещества простейший способ превращения биомассы в энергию заключается в сгорании — оно обеспечивает тепло, которое в свою очередь превращается в механическую или электрическую энергию. Что же касается сырого вещества, то в этом случае древнейшим и наиболее эффективным методом превращения биомассы в энергию является получение биогаза (метана). Метановое «брожение», или биометаногенез, — давно известный процесс превращения биомассы в энергию[48]. Он был открыт в 1776 г. Вольтой, который установил наличие метана в болотном газе. Биогаз, получающийся в ходе этого процесса, представляет собой смесь из 65 % метана, 30 % углекислого газа, 1 % сероводорода (H2S) и незначительных количеств азота, кислорода, водорода и закиси углерода. Болотный газ дает пламя синего цвета и не имеет запаха[49]. Его бездымное горение причиняет гораздо меньше неудобств людям по сравнению со сгоранием дров, навоза жвачных животных или кухонных отбросов. Энергия, заключенная в 28 м3 биогаза, эквивалентна энергии 16,8 м3 природного газа, 20,8 л нефти или 18,4 л дизельного топлива. Биометаногенез осуществляется в три этапа: растворение и гидролиз органических соединений, ацидогенез и метаногенез. В энергоконверсию вовлекается только половина органического материала — 1800 ккал/кг сухого вещества по сравнению с 4000 ккал при термохимических процессах, но остатки, или шлаки, метанового «брожения» используются в сельском хозяйстве как удобрения. В процессе биометаногенеза участвуют три группы бактерий. Первые превращают сложные органические субстраты в масляную, пропионовую и молочную кислоты; вторые превращают эти органические кислоты в уксусную кислоту, водород и углекислый газ, а затем метанообразующие бактерии восстанавливают углекислый газ в метан с поглощением водорода, который в противном случае может ингибировать уксуснокислые бактерии. В 1967 г. Брайант и др. установили, что уксуснокислые и метанообразующие микроорганизмы образуют симбиоз, который ранее считался одним микробом и назывался Methanobacillus omelianskii. Для всех метанобактерий характерна способность к росту в присутствии водорода и углекислого газа, а также высокая чувствительность к кислороду и ингибиторам производства метана. В природных условиях метанобактерии тесно связаны с водородобразующими бактериями: эта трофическая ассоциация выгодна для обоих типов бактерий. Первые используют газообразный водород, продуцируемый последними; в результате его концентрация снижается и становится безопасной для водородобразующих бактерий. Метановое «брожение» происходит в водонепроницаемых цилиндрических цистернах (дайджестерах) с боковым отверстием, через которое вводится ферментируемый материал. Над дайджестером находится стальной цилиндрический контейнер, который используется для сбора газа; нависая над бродящей смесью в виде купола, контейнер препятствует проникновению внутрь воздуха, так как весь процесс должен происходить в строго анаэробных условиях. Как правило, в газовом куполе имеется трубка для отвода биогаза. Дайджестеры изготовляют из глиняных кирпичей, бетона или стали. Купол для сбора газа может быть изготовлен из нейлона; в этом случае его легко прикреплять к дайджестеру, изготовленному из твердого пластического материала. Газ надувает нейлоновый мешок, который обычно соединен с компрессором для повышения давления газа. В тех случаях, когда используются отходы домашнего хозяйства или жидкий навоз, соотношение между твердыми компонентами и водой должно составлять 1:1 (100 кг отходов на 100 кг воды), что соответствует общей концентрации твердых веществ, составляющей 8-11 % по весу. Смесь сбраживаемых материалов обычно засевают ацетогенными и метаногенными бактериями или отстоем из другого дайджестера. Низкий pH подавляет рост метаногенных бактерий и снижает выход биогаза; такой же эффект вызывает перегрузка дайджестера. Против закисления используют известь. Оптимальное «переваривание» происходит в условиях, близких к нейтральным (pH 6,0–8,0). Максимальная температура процесса зависит от мезофильности или термофильности микроорганизмов (30–40 °C или 50–60 °C); резкие изменения температуры нежелательны. Обычно дайджестеры загружают в землю, чтобы использовать изоляционные свойства почвы. В странах с холодным климатом их нагревают при помощи устройств, которые применяют при компостировании сельскохозяйственных отходов. С точки зрения питательных потребностей бактерий избыток азота (например, в случае жидкого навоза) способствует накоплению аммиака, который подавляет рост бактерий. Для оптимальной переработки соотношение C/N должно быть порядка 30:1 (по весу). Это соотношение можно изменять, смешивая субстраты, богатые азотом, с субстратами, богатыми углеродом. Так, C/N навоза можно изменить добавлением соломы или жома сахарного тростника. Отходы пищевой промышленности и сельскохозяйственного производства характеризуются высоким содержанием углерода (в случае перегонки свеклы на 1 л отходов приходится до 50 г углерода), поэтому они лучше всего подходят для метанового «брожения», тем более, что некоторые из них получаются при температуре, наиболее благоприятной для этого процесса. Желательно перемешивать суспензию сбраживаемых веществ, чтобы воспрепятствовать расслаиванию, которое подавляет брожение. Твердый материал необходимо раздробить, так как наличие крупных комков препятствует образованию метана. Обычно длительность переработки навоза крупного рогатого скота составляет две — четыре недели. Двухнедельной переработки при температуре 35 °C достаточно, чтобы убить все патогенные энтеробактерии и энтеровирусы, а также 90 % популяции Ascaris lumbricoides и Ancylostoma. Конференция ООН по науке и технике для развивающихся стран (1979) и эксперты Экономической и социальной комиссии по странам Азии и Тихого океана подчеркнули достоинства интегрированных сельскохозяйственных программ, использующих биогаз. Такие программы направлены на разработку пищевых культур, а также на производство белка культурами водорослей, создание рыбных ферм, переработку отходов и превращение различных отбросов в удобрения и энергию в виде метана. Надо отметить, что 38 % от 95-миллионного поголовья крупного рогатого скота в мире, 72 % остатков сахарного тростника и 95 % отходов бананов, кофе и цитрусовых приходятся на долю стран Африки, Латинской Америки, Азии и Ближнего Востока. Не удивительно, что в этих регионах сосредоточены огромные количества сырья для метанового «брожения». Следствием этого явилась ориентация некоторых стран сельскохозяйственно ориентированной экономикой на биоэнергетику. Например, одним из основных принципов энергетической политики Индии является производство биогаза в сельских районах. В конце 1979 г. в Индии работало менее 100 ООО установок. В Китае в этот же период насчитывалось 10 млн. установок. Сырьем для загрузки установок в этих странах являются отходы животноводческих ферм и птицефабрик. В Центральной Америке построены установки, работающие на отходах производства кофе. В Масатенанго была построена фабрика, выпускающая 90 м3 биогаза в сутки и 900 т органических удобрений в год из отходов кофе. Биогаз обеспечивает работу двигателя мощностью 35 л. с., являющегося частью устройства, которое лущит кофе со скоростью 3 т/ч, вырабатывает 1500 В электроэнергии и обеспечивает работу компрессора. В Израиле с 1974 г. производством биогаза занимается «Ассоциация киббуци индастриз» (KIA). Проведены фундаментальные исследования процесса метаногенеза при активном участии нескольких университетов и промышленных исследовательских институтов под эгидой министерства энергетики. Анаэробное брожение происходит при 55 °C. Исследователям удалось добиться повышения выхода биогаза до 4–6,5 м3 в сутки на каждый кубометр объема цистерны дайджестера (что в десять раз превышает обычный выход). Биогаз состоит из 62 % метана и 38 % углекислого газа; последний предполагают использовать в теплицах для ускорения фотосинтеза культивируемых растений. Отходы переработки, содержащие только 12 % твердого вещества, скармливают рыбам. Это помогло сэкономить половину гранулированных кормов из злаков, которые обычно употребляют при разведении рыб. Как показали эксперименты, богатые белками, минеральными солями и витаминами отходы крупного рогатого скота и овец можно использовать в качестве корма для скота, заменяя ими до 25 % сухого вещества поглощаемой пищи. Производство биогаза путем метанового «брожения» отходов — одно из возможных решений энергетической проблемы в большинстве сельских районов развивающихся стран. И хотя при использовании коровьего навоза только четверть органического материала превращается в биогаз, последний выделяет тепла на 20 % больше, чем его можно получить при полном сгорании навоза. Производство биогаза имеет следующие достоинства: это источник энергии, доступный на семейном и общинном уровне; отходы процесса служат высококачественными удобрениями и в довершение сам процесс способствует поддержанию чистоты окружающей среды. Чтобы обеспечить крупномасштабное развитие и экономическую выгоду предприятий по производству биогаза, необходимо решить целый ряд биохимических, микробиологических и социальных проблем. Усовершенствования касаются следующих областей: сокращения числа стальных элементов в используемом оборудовании; создания оборудования с оптимальной конструкцией; разработки эффективных нагревателей; нагрева дайджестеров за счет солнечной энергии; объединения систем производства биогаза с другими нетрадиционными источниками энергии; конструирования крупномасштабных производственных единиц для сельских или городских общин; оптимального использования переработанных отходов и, наконец, усовершенствования процессов брожения и начальной деградации отходов. Биотехнология в состоянии внести крупный вклад в решение проблем энергетики посредством производства достаточно дешевого биосинтетического этанола, который кроме того является и важным сырьем для микробиологической промышленности при получении пищевых и кормовых белков, а также белково-липидных кормовых препаратов. Источником углеводородов также могут служить водоросли. У широко распространенной зеленой водоросли Botryococcus braunii (обитающей в пресной и солоноватой воде умеренных и тропических зон) углеводороды в зависимости от условий роста и разновидностей могут составлять до 75 % сухой массы. Они накапливаются внутри клеток, и водоросли, в которых их много, плавают на поверхности. После сбора водорослей эти углеводороды легко отделить экстракцией каким-нибудь растворителем или методом деструктивной отгонки. Таким путем может быть получено вещество, аналогичное дизельному топливу и керосину. Встречается несколько разновидностей В. braunii, отличающихся пигментацией и структурой синтезируемых углеводородов. Зеленая разновидность содержит линейные углеводороды с нечетным (25–31) числом атомов углерода, бедных двойными связями. Красная водоросль содержит углеводороды с 34–38 атомами углерода и несколькими двойными связями; это так называемые "ботриококкцены". Смысл существования двух разновидностей в настоящее время изучается. Углеводороды накапливаются в клеточной стенке, их синтез связан с метаболической активностью водоросли в фазе роста. Выход углеводородов при создании оптимальных условий культивирования может достигать 60 т/га*год для культуры водорослей, выращиваемой в толще воды в природных или искусственных условиях. Для определения перспективности использования В. braunii необходимо провести следующие исследования: — определить условия, обеспечивающие максимальную скорость роста и образования углеводов в лабораторных и полевых условиях; — выяснить, можно ли добиться скорости роста В. braunii, сопоставимой с известной для других водорослей; — разработать соответствующие методы выращивания, сбора и переработки; — оценить применимость получаемого продукта как альтернативного источника топлива и смазочных веществ. Исследования, связанные с выделением и возможностью утилизации углеводородов В. braunii, могут также способствовать лучшему пониманию вопроса о происхождении нефти.
Биотехнология обработки стоков и контроль загрязнения воды тяжелыми металлами Развитие промышленности ведет к образованию большого количества отходов, в том числе отходов, содержащих новые антропогенные компоненты. Методами биотехнологии эти отходы могут быть переработаны в полезные или безвредные продукты. Бытовые отходы делятся на 2 группы: твердые отходы и сточные воды. Твердые бытовые отходы состоят из целлюлозосодержащих материалов (до 40 % бумаги, 2.5 % дерева, 8 % текстиля) и пищевых отходов (40 %). Наиболее экономична и радикальна переработка их метановым брожением, в результате образуется легко транспортируемое топливо — метан. Сточные воды обычно содержат сложную смесь нерастворимых и растворимых компонентов различной природы и концентрации. Бытовые отходы, как правило, содержат почвенную и кишечную микрофлору, включая патогенные микроорганизмы. Сточные воды сахарных, крахмальных, пивных и дрожжевых заводов, мясокомбинатов содержат в больших количествах углеводы, белки и жиры, являющиеся источниками питательных веществ и энергии. Стоки химических и металлургических производств могут содержать значительное количество токсических и даже взрывчатых веществ. Серьезное загрязнение возникает при попадании в окружающую среду соединений тяжелых металлов, таких как железо, медь, олово и др. Цель очистки сточных вод — удаление растворимых и нерастворимых компонентов, элиминирование патогенных микроорганизмов и проведение детоксикации таким образом, чтобы компоненты стоков не вредили человеку, не загрязняли водоемы. Бактерии рода Pseudomonas практически всеядны. Например, P. putida могут утилизировать нафталин, толуол, алканы, камфару и др. соединения. Выделены чистые культуры микроорганизмов, способные разлагать специфические фенольные соединения, компоненты нефти в загрязненных водах и т. д. Микроорганизмы рода Pseudomonas могут утилизировать и необычные химические соединения — инсектициды, гербициды и другие ксенобиотики. Генетически сконструированные штаммы микроорганизмов в будущем смогут решить проблему очистки сточных вод и почв, загрязненных пестицидами и другими антропогенными веществами. Азотсодержащие соединения (белки, аминокислоты, мочевина) могут быть удалены в биологическом процессе денитрификации-нитрификации. Биологическое удаление азота и фосфора, являющихся причинами эвтрофикации (зарастания озер микроводорослями, которые бурно размножаются, затем отмирают, давая пищу аэробным бактериям, потребляющими кислород, что приводит к замору рыбы) озер и каналов, находится в стадии экспериментов. Тяжелые металлы затрудняют биологические процессы очистки стоков и отрицательно влияют на флору и фауну. Природные штаммы микроорганизмов не могут быть использованы для накопления этих металлов в силу их высоко токсичности. Однако, есть белок высших организмов — металлотионеин, который активно связывает различные тяжелые металлы. Ген, кодирующий синтез мышиного металлотионеина, клонирован в бактериях. Это открывает возможность получения белка в больших количествах с использование иммобилизованных бактерий и его использования для связывания и экстракции тяжелых металлов.
Сельскохозяйственная биотехнология Биологическая азотфиксация — процесс фиксации атмосферного азота бактериями, живущими в симбиозе с представителями семейства бобовых. Для ускорения заселения ризосферы обычно используют бактериальные удобрения, содержащие культуры азотфиксирующих микроорганизмов, например, клубеньковых бактерий. Методами генной инженерии выведены мутанты клубеньковых бактерий с повышенной способностью к азотфиксации. Ведутся работы по созданию азотфиксирующих растений, способных к симбиозу со злаковыми. Микробные инсектициды. В последнее время все чаще появляются данные о мутагенном и канцерогенном действии химических пестицидов, которые плохо разрушаются и накапливаются в окружающей среде. Для получения микробных инсектицидов используются вирусы, грибы, простейшие, наиболее удобны — спорообразующие бактерии. Микробные инсектициды высоко специфичны и действуют только на определенные вредные насекомые, оставляя невредимыми полезные. Патогенность микроорганизмов вызвана действием определенных токсинов, поэтому выработки устойчивости к биопрепаратам у насекомых не происходит. Микробные пестициды подвержены биодеградации. Микроорганизмы могут регулировать рост растений и животных, подавлять заболевания. Некоторые бактерии изменяют кислотность и соленость почвы, другие продуцируют соединения, связывающие железо, третьи — вырабатывают регуляторы роста. Как правило, микроорганизмами инокулируют семена и или растения перед посадкой. В животноводстве биотехнология также находит применение. Это диагностика, профилактика, лечение заболеваний с использованием техники моноклональных антител, генетическое улучшение пород животных. Некоторые вещества, полученные с помощью микроорганизмов могут использоваться в виде кормовых добавок, другие — подавляют вредную микрофлору в желудочно-кишечном тракте или стимулируют образование специфических микробных метаболитов. Подробнее вопросы бактериальных удобрений и энтомопатогенных препаратов для сельского хозяйства рассматриваются в главе: "Биотехнология препаратов для сельского хозяйства", а генно-инженерных модификаций сельскохозяйственных растений и животных в разделе "трансгенные животные".
Биогеотехнология Некоторые микроорганизмы могут катализировать определенные окислительно-восстановительные реакции — окисление Fe и Мn в воде, окисление серосодержащих соединений, окисление-восстановление азотсодержащих соединений. Аэробные бактерии могут выделять железо, медь, сульфаты. Биогеотехнология — использование геохимической деятельности микроорганизмов в горнодобывающей промышленности. Это экстракция и концентрирование металлов при биологической очистке сточных вод предприятий горнодобывающей промышленности и флотационных процессах: выщелачивание бедных и отработанных руд, десульфирование каменного угля, окисление пиритов и пиритсодержащих пород. Своими корнями биогеотехнология уходит в геологическую микробиологию. Микроорганизмы принимали и принимают активное участие в геологических процессах. Биологические свойства различных групп микроорганизмов и особенности их жизнедеятельности в месторождениях полезных ископаемых составляют научные основы биогеотехнологии. Биогеотехнология стихийно зародилась еще в XVI в. До нас дошли сведения о том, что в те далекие времена в Венгрии для получения меди груды добытой руды орошали водой. Этот нехитрый технологический прием оказался прообразом современного бактериально-химического метода кучного выщелачивания металлов из руд. Конечно, тогда еще не знали, что используемый процесс получения меди по своей природе является микробиологическим. Это стало известно только в 1922 г. благодаря работам немецких ученых Рудольфа и Хельброннера. По-видимому, 1922 г. следует считать официальной датой рождения биогеотехнологии. В дальнейшем биогеотехнология развивалась неровно и своего совершеннолетия достигла к началу 80-х годов нашего века. К этому времени наряду с бактериальным выщелачиванием металлов сформировались и другие разделы биогеотехнологии — удаление серы из углей, борьба с метаном в угольных шахтах, повышение нефтеотдачи пластов. Биогеотехнология выщелачивания металлов — использование главным образом тионовых (окисляющих серу и серосодержащие соединения) бактерий для извлечения металлов из руд, рудных концентратов и горных пород. При переработке бедных и сложных руд тысячи и даже миллионы тонн ценных металлов теряются в виде отходов, шлаков, «хвостов». Происходят также выбросы вредных газов в атмосферу. Бактериально-химическое выщелачивание металлов уменьшает эти потери. Основу этого процесса составляет окисление содержащихся в рудах сульфидных минералов тионовыми бактериями. Окисляются сульфиды меди, железа, цинка, олова, кадмия и т. д. При этом металлы из нерастворимой сульфидной формы переходят в сульфаты, хорошо растворимые в воде. Из сульфатных растворов металлы извлекаются путем осаждения, экстракции, сорбции. Одним из возможных путей извлечения металлов из растворов является адсорбция металлов клетками живых микроорганизмов, так называемая биосорбция металлов. Металлы включаются в состав специфических белков — металлотионеинов. Полезными для биогеотехнологии добычи металлов свойствами обладает целый ряд микроорганизмов. Но основным из них, безусловно, является открытый в 1947 г. Колмером и Кинкелем вид тионовых бактерий, названный Thiobacillus ferrooxidans. Необходимую для роста энергию эти бактерии получают при окислении восстановленных соединений серы и двухвалентного железа в присутствии свободного кислорода. Они окисляют практически все известные в настоящее время сульфиды металлов. Источником углерода для роста бактерий служит при этом углекислый газ. Характерной особенностью их физиологии является потребность в очень кислой среде. Они развиваются при pH от 1 до 4,8 с оптимумом при 2–3. Интервал температур, в котором могут развиваться бактерии этого вида, составляет от 3 до 40 °C с оптимумом при 28 °C. Тионовые бактерии широко распространены в природе. Они обитают в водоемах, почвах, угольных и золоторудных месторождениях. В значительных количествах встречаются они в месторождениях серных и сульфидных руд. Но в условиях естественного залегания таких руд активность тионовых бактерий сдерживается отсутствием кислорода. При разработке сульфидных месторождений руды вступают в контакт с воздухом, и в них развиваются микробиологические процессы, приводящие к выщелачиванию металлов. Применяя определенные биотехнологические мероприятия, этот естественный процесс можно ускорить. Основной технологической операцией этого способа является орошение отвалов добытой руды растворами, содержащими серную кислоту, ионы двух- и трехвалентного железа, а также жизнеспособные клетки тионовых бактерий. Иногда для усиления процессов выщелачивания внутрь отвала подают воздух. В таких условиях выщелачивающий раствор фильтруется через толщу руды и в результате микробиологических и химических процессов обогащается извлекаемыми из руды металлами. Затем этот раствор собирают с помощью системы коллекторов, и из него извлекают металлы одним из физико-химических методов. Ежегодно в мире таким способом добывают сотни тысяч тонн меди, или примерно 5 % от ее общей добычи. В ряде стран этим способом получают также значительные количества урана. Биогеотехнология обессеривания углей — использование тионовых бактерий для удаления серосодержащих соединений из углей. Как бурые, так и каменные угли нередко содержат значительные количества серы. Общее содержание серы в углях может достигать 10–12 %. При сжигании углей содержащаяся в них сера превращается в сернистый газ, который поступает в атмосферу, где из него образуется серная кислота. Из атмосферы серная кислота выпадает на поверхность земли в виде сернокислотных дождей. По имеющимся данным, в некоторых странах Западной Европы в год на 1 га земли с дождями выпадает до 300 кг серной кислоты. Нетрудно себе представить, какой ущерб наносят кислотные дожди здоровью человека, его хозяйственной деятельности и окружающей природе. Кроме этого, высокосернистые угли плохо коксуются и поэтому не могут быть использованы в цветной металлургии. Микробное удаление серы из углей, по мнению специалистов, является экономически выгодным, и с ним связывают надежды на решение проблемы сернокислотных дождей. Первые опыты по направленному удалению серы из угля с использованием микроорганизмов были выполнены в 1959 г. в нашей стране 3. М. Зарубиной, Н. Н. Ляликовой и Е. И. Шмук. В результате этих опытов за 30 суток с участием бактерий Th. ferrooxidans из угля было удалено 23–30 % серы. Позднее несколько работ по микробиологическому обессериванию угля было опубликовано американскими исследователями. Им удалось с помощью тионовых бактерий снизить содержание пиритной серы в каменном угле за четверо суток почти на 50 %. Этот метод будет сопровождаться попутным выщелачиванием различных металлов. Известно, что в заметных количествах содержится в углях германий, никель, бериллий, ванадий, золото, медь, кадмий, свинец, цинк, марганец. Попутное получение ценных металлов при десульфуризации угля должно дать дополнительный экономический эффект. Работы по удалению пиритной серы из угля микробиологическим путем проводятся сейчас во многих странах мира. По последним сообщениям в лабораторных условиях удается снизить содержание серы в угле путем микробиологического выщелачивания за 5 суток почти на 100 %. Микробиологический способ десульфуризации углей рассматривается как весьма перспективный. Биогеотехнология и борьба с метаном в угольных шахтах — использование метанокисляющих бактерий для снижения концентрации метана в угольных пластах и выработанных пространствах. В пластах каменного угля содержится огромное количество метана, достигающее сотни кубометров в 1 т угля. При этом чем глубже залегает уголь в недрах земли, тем больше метана он содержит. При подземной добыче угля метан из разрабатываемых угольных пластов и образующихся при этом выработанных пространств поступает в атмосферу шахт. Скопления этого взрывоопасного газа в горных выработках создают постоянную угрозу для жизни шахтеров. Известны случаи крупных взрывов метана в угольных шахтах мира, унесшие сотни человеческих жизней. Традиционные средства борьбы с метаном в угольных шахтах (вентиляция, вакуумная дегазация, увлажнение пластов водой) в условиях постоянной интенсификации горных работ и перехода на все более глубокие угленосные горизонты часто уже не могут обеспечить одновременно высокий уровень угледобычи и безопасные условия труда. В основе биогеотехнологических способов борьбы с метаном лежит процесс поглощения этого газа метанокисляющими бактериями в угольных пластах и выработанных пространствах. На данном уровне развития наук этот процесс представляет собой единственную возможность разрушения молекулы метана при температурах разрабатываемых угленосных толщ. Идея об использовании метанокисляющих бактерий для борьбы с метаном в угольных шахтах принадлежит советским ученым. В 1939 г. А. 3. Юровский, Г. П. Капилаш и Б. В. Мангуби предложили применять эти бактерии для снижения выделения метана из выработанных пространств. Несмотря на широкое распространение метанокисляющих бактерий в природе, в угольных пластах и прилегающих породах они отсутствуют. Поэтому необходимое количество активных метанокисляющих бактерий выращивают в ферментерах и в виде суспензии в питательной среде подают в поровый объем угольных пластов и выработанные пространства. Рабочая суспензия приготовляется непосредственно в шахте. В рудничную воду добавляют заданное количество биомассы метанокисляющих бактерий и недостающие для их активной жизнедеятельности минеральные соли. Обычно это минеральные соединения азота и фосфора. В угольный пласт рабочая суспензия нагнетается насосами через скважины, пробуренные по углю или из подземных выработок, или с поверхности земли: 1 т угля может принять 20–40 л рабочей суспензии. В угле микроорганизмы распределяются по трещинам и порам. Таким путем осуществляется насыщение угля метаноокисляющими бактериями. Но для развития этих бактерий необходим свободный кислород, которого нет в угольных пластах.Поэтому в насыщенный метанокисляющими бактериями участок угольного пласта через те же скважины компрессором постоянно прокачивается воздух. В таких условиях бактерии потребляют содержащийся в угле метан, и за счет этого происходит уменьшение исходной газоносности угольного пласта. Микробиологические способы борьбы с метаном были неоднократно испытаны в угольных шахтах. Поступление метана как из угольных пластов, так и из выработанных пространств в ходе этих испытаний было снижено в среднем в 2 раза. При прочих равных условиях это позволяет повышать добычу угля примерно в 1,5 раза. Биогеотехнология и повышение нефтеотдачи пластов — использование различных групп микроорганизмов для увеличения вторичной добычи нефти. Нефть, как известно, является в настоящее время основным энергетическим и химическим сырьем. Однако по некоторым прогнозам мировые запасы нефти могут быть исчерпаны уже в течение ближайших 50 лет. Вместе с тем существующая технология позволяет извлекать только половину нефти, содержащейся в месторождениях. Это обусловлено прочной связью нефти с вмещающими ее породами. Повышение нефтеотдачи пластов на 10–15 % было бы равносильно открытию новых месторождений. В связи с этим в настоящее время заметно возрос интерес к поиску путей и средств повышения вторичной добычи нефти. Один из способов предполагает использование комплекса углеводородокисляющих и метанобразующих бактерий для увеличения нефтеотдачи пластов основано на активации геохимической деятельности этих микробов в нефтяной залежи, куда они попадают вместе с закачиваемыми через скважины поверхностными водами. Активация названных микробиологических процессов достигается путем аэрации закачиваемых вод и добавления в них минеральных солей азота и фосфора. Недостаток этих химических элементов чаще всего лимитирует активность микрофлоры в природных условиях. Нагнетание в нефтяную залежь обогащенной кислородом и минеральными солями воды приводит к образованию аэробной зоны в нефтеносном пласте вокруг нагнетательной скважины. Здесь начинают интенсивно идти процессы разрушения нефти аэробными углеводородокисляющими микробами. Это сопровождается накоплением углекислого газа, водорода и низкомолекулярных органических кислот, которые поступают в анаэробную зону нефтяной залежи. Здесь они превращаются метанобразующими бактериями в метан. Разрушение нефти и образование газов приводят к разжижению нефти и повышению газового давления в нефтеносном пласте, что и должно сопровождаться увеличением добычи нефти из добывающих скважин.
Биоэлектроника В области электроники биотехнология может быть использована для создания улучшенных типов биосенсоров и новых приводящих устройств, называемых биочипы. Биотехнология делает возможным создание устройств, в которых белки являются основой молекул, действующих как полупроводники. Для индикации загрязнений различного происхождения в последнее время стали использовать не химические реагенты, а биосенсоры — ферментные электроды, а также иммобилизованные клетки микроорганизмов. Ферменты обладают высочайшей чувствительностью. Биоселективные датчики создают также путем нанесения на поверхность ионоселективных электродов целых клеток микроорганизмов или тканей. Например, Neurospora europea — для определения NH3, Trichosporon brassiacae — для определения уксусной кислоты. В качестве сенсоров используют также моноклональные антитела, обладающие исключительно высокой избирательностью. Лидерами в производстве биодатчиков и биочипов являются японские компании, такие как Hitachi, Sharp. Например, компания Hitachi в начале 90-х годов создает проектную групп численностью в 200 человек исключительно для работ в области биоэлектроники. Компания Sharp проводит исследования по разработке компьютеров с биокомпонентами. Появляется новый тип полупроводников, проводящую функцию в которых осуществляют молекулы белков. Такие ферментные системы работают с большей скоростью, чем кремниевые полупроводники. Биочипы имеют небольшие размеры, надежны и способны к самосборке. Еще одна японская компания, Sony, запатентовала способ производства высококачественных акустических систем из целлюлозы, образуемой бактериями. Гелеобразная целлюлоза высушивается. Полученный материал имеет структуру сот и используется в качестве плоской диафрагмы акустических систем.
Биотехнология в медицине Антибиотики — самый большой класс фармацевтических соединений, синтез которых осуществляется микробными клетками. К этому же классу относятся противогрибковые агенты, противоопухолевые лекарства и алкалоиды. В 1980 г. мировое производство антибиотиков составляло примерно 25000 т, из них 17000 т — пенициллины, 5000 т — тетрациклины, 1200 т — цефалоспорины и 800 т — эритромицины. В 1945 г. Бротзу из Института гигиены в Кальари (Сардиния) выделил из пробы морской воды плесень Cephalosporium acremonium, синтезирующую несколько антибиотиков; один из них, цефалоспорин С, оказался особенно эффективен против устойчивых к пенициллину грамположительных бактерий. Из нескольких тысяч открытых антибиотиков львиная доля принадлежит актиномицетам. Среди актиномицетов наибольший вклад вносит род Streptomyces, включая тетрациклины (один только вид Streptomyces griseus синтезирует более пятидесяти антибиотиков). Наиболее распространенными с коммерческой точки зрения оказались пенициллины, цефалоспорины и тетрациклины. Начиная с середины 1960-х гг. в связи с возросшей сложностью выделения эффективных антибиотиков и распространением устойчивости к наиболее широко применяемым соединениям у большого числа патогенных бактерий исследователи перешли от поиска новых антибиотиков к модификации структуры уже имеющихся. Они стремились повысить эффективность антибиотиков, найти защиту от инактивации ферментами устойчивых бактерий и улучшить фармакологические свойства препаратов. Большинство исследований было сосредоточено на пенициллинах и цефалоспоринах, структура которых включает четырехчленное b-лактамное кольцо. Добавление к b-лактамному кольцу метоксильной (СН3О) — группы привело к появлению цефамицинов, близких к цефалоспоринам и эффективных как против грамотрицательных, так и против пенициллиноустойчивых микробов. Полусинтез состоит в замене химическим путем одной боковой цепи b-лактамного кольца на другую в полученной ферментацией молекуле. Устойчивость к пенициллинам и цефалоспоринам связана с наличием ферментов, так называемых Ь-лактамаз, которые широко распространены среди бактерий, актиномицетов, цианобактерий и дрожжей. Так как гены, кодирующие эти ферменты, находятся в составе плазмид, устойчивость может передаваться при переносе плазмид от одного бактериального штамма к другому. Исследователи фирмы «Мерк, Шарп и Доум» открыли новый класс b-лактамных антибиотиков, тиенамицины, продуцируемых Streptomyces cattleya. Тиенамицины чрезвычайно эффективны против грамположительных и грамотрицательных бактерий, а также способны ингибировать b-лактамазы, что значительно повышает возможности этих антибиотиков. К ингибиторам b-лактамаз относятся также клавулановая и оливановая кислоты, идентифицированные исследователями английской фармацевтической компании «Бичем». Компания выпустила новый антибиотик, аугментин, который представляет собой комбинацию b-лактамного антибиотика амоксициллина и клавулановой кислоты. Антибиотики вырабатываются в результате совместного действия продуктов 10–30 генов, поэтому практически невозможно обнаружить отдельные спонтанные мутации, которые могли бы повысить выход антибиотика с нескольких миллиграммов на литр в штамме дикого типа до 20 г/л и более пенициллина или тетрациклина в промышленных штаммах Penicillium chrysogenum или Streptomyces auerofaclens. Эти высокопродуктивные штаммы были получены в результате последовательных циклов мутагенеза и селекции. В результате мутаций появились новые вторичные метаболиты, в том числе 6-деметилхлортетрациклин и 6-деметилтетрациклин. Определенные мутанты, так называемые идиотрофы, способны синтезировать только половину молекулы антибиотика, а среда должна быть обогащена другой ее половиной. Такая форма мутационного биосинтеза привела к открытию новых производных антибиотиков, среди них принадлежащие к аминоциклитольной группе. Число противоопухолевых веществ микробного происхождения довольно ограниченно. Блеомицин, выделенный Умезавой с сотр. в Токийском институте микробной химии из культур Streptomyces verticilliis, представляет собой гликопептид, который действует, разрывая ДНК опухолевых клеток и нарушая репликацию ДНК и РНК. Другая группа противоопухолевых агентов создана на основе комбинации аминогликозпдной единицы и молекулы антрациклина. Недостатком обоих соединений является их потенциальная опасность для сердца. В медицине применяют также аминокислоты, например, аргинин. В сочетании с аспартатом или глутаматом он помогает при заболевании печени. К-Na-аспартат снимает усталость и облегчает боли в сердце, его рекомендуют при заболевании печени и диабете. Цистеин защищает SH-ферменты в печени и других тканях от окисления и оказывает детоксицирующее действие. Он проявляет также защитное действие от повреждения, вызываемых облучением. Дигидроксифенилаланин и D-фенилаланин эффективны при болезни Паркинсона. Из полиаминокислот получают хороший материал для хирургии. В медицине также используют зеленую водоросль Scenedesmus. Ее культивируют в жидкой питательной среде (установки дают до 80 тонн водорослей в год), извлекают и проводят экстракцию этиловым спиртом. Биомассу отделяют и подвергают ферментативному гидролизу щелочной протеазой. Около 50 % белков при этом распадается до пептидов. Гидролизат содержит почти все незаменимые аминокислоты, представляет собой порошок желтовато-зеленого цвета с приятным запахом и вкусом. Используется этот продукт для быстрого восстановления организма, а также как компонент косметических средств. Если вместо обработки этанолом провести двукратную экстракцию дистиллированной водой, а затем высушить, то получается порошок светло-желтого цвета. Его используют как биостимулятор и готовят из него препараты для лечения плохо заживающих ран. Еще в начале 90-х годов появились статьи, в которых рассматривались перспективы использования сапротрофной микрофлоры как продуцента биологически активных веществ (БАВ). Предполагается вводить в организм сапрофитные микроорганизмы, которые могли бы жить в условиях симбиоза с нормальной микрофлорой организма. Вещества, вырабатываемые бактериальными штаммами включаются в систему биохимических процессов организма. В случае нарушения нормального биохимического статуса организма они корректируют его, а при патологическом процессе — задерживают его или способствуют прекращению. Такое введение получило название «микробиологическая подсадка». К 1992 году было описано уже более 50 таких штаммов и проанализирован диапазон биологических эффектов секретируемых БАВ. Для лечения широкого спектра заболеваний (бактериальные инфекции кишечника, дыхательных путей, гнойных инфекций, аллергий) успешно применяются штаммы Bacillus subtilis (препарат «Бактисубтил», например, используют при лечении диареи). Штаммами Е. coli лечат ряд кишечных заболеваний. БАВ, секретируемые сапротрофами, могут регулировать ферментативные процессы в организме и вступать во взаимодействие с поступающими в организм ксенобиотиками. Штаммы можно получать непосредственно от человека, тогда они будут представлять его естественную микрофлору. Можно целенаправленно выводить лабораторные мутантные штаммы, в том числе методами генной инженерии и вводить их в организм. Способы введения могут быть различны: капсулы, растворимые в кишечном соке, культуры штаммов-продуцентов на пленочной основе, в виде свечей, а при легочных заболеваниях — в виде аэрозолей. Новым направлением в медицине является использование ферментных препаратов типа «контейнер», изготовление которых стало возможным появлению и совершенствованию методов иммобилизации веществ. Эти препараты представляют собой микросферы с более или менее твердой и проницаемой оболочкой. Назначение этих лекарственных препаратов различное. Первым типом «искусственных клеток» следует назвать микрокапсулы. Фермент, находящийся внутри оболочки, не контактирует с жидкостями и тканями организма, не разрушается протеиназами, не ингибируется, не вызывает иммунного ответа организма. Основное достоинство микрокапсул заключается в том, что их можно имплантировать в нужное место, например в непосредственной близости от опухоли. При этом микрокапсула с соответствующим содержанием будет перерабатывать метаболиты, необходимые для роста опухолевой ткани, и эта ткань не будет развиваться. Капсулы могут содержать микроскопические участки тканей. Например, имеются экспериментальные данные по созданию депо инсулина путем имплантации микрокапсул, содержащих островки Лангерганса, синтезирующие в поджелудочной железе инсулин. Известно, что терапии диабетических заболеваний уделяется много внимания. Имплантация лекарственного начала избавила бы пациентов от ежедневных инъекций инсулина. Следует учитывать, что микрокапсулы, вводимые в кровь, могут забивать кровеносные сосуды и, следовательно, являться причиной образования тромбов. Однако эффективность микрокапсул при использовании их в виде колонок для диализа в аппарате «искусственная почка» несомненна. При этом объем аппаратов и, соответственно, количество необходимых и очень дорогих растворов резко сокращается. Например, для микрокапсулированной «искусственной почки» требуется колонка объемом всего 30 мл, которая работает почти в 100 раз быстрее обычного аппарата. Развитие такой техники сдерживается пока высокой стоимостью, а также необходимостью использовать уже существующую тоже очень дорогую технику. Вероятно, ферментные реакторы на микрокапсулах будут применяться для деградации недиализуемых материалов. Внутрь микрокапсул могут быть включены магнитные частицы. В этом случае извне подводят магнитное поле и препарат удерживают вблизи органа-мишени. В ряде случаев используются высокомолекулярные соединения, растворимые в определенных условиях и сохраняющие высокую прочность оболочек в других. Так ведет себя ацетилфталилцеллюлоза, микрокапсулы из которой интактны в желудочном соке и растворяются в кишечнике, освобождая содержимое. Сейчас интенсивно исследуются свойства микрокапсул, стенка которых состоит из оболочек эритроцитов. Содержимое эритроцитов удаляется, а «тень» заполняется ферментом. Серьезные успехи достигнуты при лечении аспарагин-зависимых опухолей препаратами аспарагиназы в оболочках эритроцитов. Используются оболочки и других клеток. Так, описаны лекарственные препараты, включенные в оболочки макрофагов. Последние имеют тенденцию накапливаться в очагах воспалений, а следовательно, могут транспортировать туда как низко-, так и высокомолекулярный лекарственный препарат. Существенной положительной стороной «теней» клеток в качестве носителя является их полная совместимость с организмом пациента, поскольку этот носитель готовят на основе клеток, выделенных из крови пациента, и возвращают их ему же с новым содержимым. Задача введения лекарственного препарата в клетки может быть решена путем создания контейнеров-переносчиков типа липосом или мицелл. Оболочка липосомы представляет собой однослойную или многослойную поверхность, образованную, в свою очередь, бислойной структурой, созданной соединениями, имеющими два гидрофобных, достаточно протяженных участка и гидрофильную группу. Гидрофобные концы слипаются между собой и образуют пленку, обе стороны которой гидрофильны. Липосома, специфически или неспецифически адсорбировавшись на клетке, может быть поглощена ею путем фагоцитоза, и фермент внутри высвобождается. Хорошо известно, что протеиназы, расщепляя денатурированные белки, способствуют очищению ран, и следовательно, их заживлению. В этом направлении в клинической практике с помощью иммобилизованных протеиназ сделано многое. В качестве носителей для иммобилизации протеолитических ферментов наиболее употребимы волокнистые материалы на основе целлюлозы, поливинилового спирта, солей альгиновой кислоты, полиамидное и коллагеновое волокно. Готовят нити, в которые при формовании включают фермент и используют их в качестве шовного материала. Сравнительный анализ действия нативных и иммобилизованных протеиназ (в основном химотрипсина, трипсина, коллагеназы) показал, что уже на 2-4-й день рана очищается от некротических масс и по крайней мере вдвое быстрее наступает грануляция. Убедительные результаты получены при лечении трофических язв, лучевых язв кожи. Особенно эффективны иммобилизованные протеиназы при предоперационной подготовке и после пластических операций. Иммобилизованные протеолитические ферменты с большим успехом применяются в лечении гнойных заболеваний легких и плевры.
Биотехнологии в пищевой промышленности Статистические данные ООН по вопросам продовольствия и сельского хозяйства свидетельствуют о том, что проблема обеспечения населения нашей планеты продуктами питания внушает серьезные опасения. По этим данным, более половины населения Земли не обеспечено достаточным количеством продуктов питания, примерно 500 млн. людей голодают, а около 2 млрд. питаются недостаточно или неправильно. К концу XX в. население нашей планеты с учетом контроля рождаемости составило 7,5 млрд. человек. Следовательно, тяжелое уже сейчас положение с продуктами питания может принять в недалеком будущем для некоторых народов угрожающие масштабы. Пища должна быть разнообразной и содержать белки, жиры, углеводы и витамины. Источники энергии — жиры и углеводы в определенных пределах взаимозаменяемы, причем их можно заменить и белками, но белки нельзя заменить ничем. Проблема питания людей в конечном счете заключается в дефиците белка. Там, где сегодня люди голодают, не хватает прежде всего белка. Установлено, что ежегодный дефицит белка в мире, по самым скромным подсчетам, оценивается в 15 млн. т. Наибольшую популярность как источники белка приобрели семена масличных культур — сои, семян подсолнечника, арахиса и других, которые содержат до 30 процентов высококачественного белка. По содержанию некоторых незаменимых аминокислот он приближается к белку рыбы и куриных яиц и перекрывает белок пшеницы. Белок из сои широко уже используется в США, Англии и других странах как ценный пищевой материал. Эффективным источником белка могут служить водоросли. Увеличить количество пищевого белка можно и за счет микробиологического синтеза, который в последние годы привлекает к себе особое внимание. Микроорганизмы чрезвычайно богаты белком — он составляет 70–80 процентов их веса. Скорость его синтеза огромна. Микроорганизмы примерно в 10-100 тысяч раз быстрее синтезируют белок, чем животные. Здесь уместно привести классический пример: 400-килограммовая корова производит в день 400 граммов белка, а 400 килограммов бактерий — 40 тысяч тонн. Естественно, на получение 1 кг белка микробиологическим синтезом при соответствующей промышленной технологии потребуется средств меньше, чем на получение 1 кг белка животного. Да к тому же технологический процесс куда менее трудоемок, чем сельскохозяйственное производство, не говоря уже об исключении сезонных влияний погоды — заморозков, дождей, суховеев, засух, освещенности, солнечной радиации и т. д. Применяя обычные технологические линии по производству синтетических волокон, можно получать из искусственных белков длинные нити, которые после пропитки их формообразующими веществами, придания им соответствующего вкуса, цвета и запаха могут имитировать любой белковый продукт. Таким способом уже получены искусственное мясо (говядина, свинина, различные виды птиц), молоко, сыры и другие продукты. Они уже прошли широкую биологическую апробацию на животных и людях и вышли из лабораторий на прилавки магазинов США, Англии, Индии, стран Азии и Африки. Только в одной Англии их производство достигает примерно 1500 тонн в год. Интересно, что белковую часть школьных обедов в США уже разрешено на 30 процентов заменять искусственным мясом, созданным на основе соевого белка. Используемое в питании больных Ричмондского госпиталя (США) искусственное мясо получило высокую оценку главного диетолога. Правда, когда больным давали антрекот из искусственного мяса, они жаловались на его тестоватость, хотя и не знали и даже не догадывались о том, что получали не естественный продукт. А когда мясо подавалось в виде мелко нарезанных кусочков, нареканий не было. Обслуживающий персонал также употреблял искусственное мясо, не догадываясь о подделке. Они воспринимали его как натуральную говядину. Врачи госпиталя отмечали также положительное влияние рациона на здоровье пациентов и особенно больных атеросклерозом. В состав такого мяса обязательно включают специально обработанный искусственный белок, небольшое количество яичного альбумина, жиры, витамины, минеральные соли, природные красители, ароматизаторы и прочее, что дает возможность «лепить» изделие с заданными свойствами, учитывая при этом физиологические особенности организма, для которого продукт предназначен. Это особенно важно в диете детей и людей пожилого возраста, больных и выздоравливающих, когда необходимо лимитировать питание по целому ряду пищевых компонентов, что весьма трудно сделать, используя традиционные продукты. Такое мясо можно резать, замораживать, консервировать, сушить или прямо использовать для приготовления различных блюд. Из 20 аминокислот, входящих в состав белков, 8 аминокислот люди не могут синтезировать, и их относят к незаменимым. Это изолейцин, лейцин, лизин, метионин, треонин, триптофан, валин, фенилаланин. Аминокислоты — это не только питательные вещества, но также ароматические и вкусовые агенты, и потому они широко используются в пищевой промышленности. Как питательную добавку в пищу чаще всего вносят лизин и метионин. Глутамат натрия и глицин употребляют как ароматические вещества для усиления и улучшения вкуса пищи. У глицина освежающий, сладкий вкус. Его вводят в сладкие напитки, и кроме того, он проявляет там бактериостатическое действие. Цистеин предотвращает подгорание пищи, улучшает пекарские процессы и качество хлеба. Благодаря некоторым бактериям удается получать около 100 г/л глутаминовой аминокислоты. Ежегодно в мире производят микробиологическим способом 270 000 т этой аминокислоты, основная часть которой идет в пищевую промышленность. По объему продукции второе место после глутаминовой кислоты занимает лизин — 180 000 т в год. Другие аминокислоты производят в гораздо меньших количествах. Аминокислоты в большом количестве применяют как добавку к растительным кормам, которые дефицитны по метионину, треонину, триптофану и особенно по лизину. Если в животных белках содержится 7–9 % лизина, то в белках пшеницы — только около 3 %. Внесение в корма лизина до содержания 0,3 % позволяет сократить их расход больше чем на 20 %. За последние 8 лет количество аминокислот, добавляемых в корма, выросло в 14 раз. Во многих странах метионин добавляют к соевой муке — белковой добавке кормов. Главная область практического применения аминокислот — обогащение кормов. Около 66 % общего количества аминокислот, получаемых в промышленности, используют в кормах, 31 % — в пище и 4 % — в медицине, косметике и как химические реактивы. На основе аминокислот готовят искусственный подсластитель — метиловый эфир L-аспартил-Ь-фенилаланина, который в 150 раз слаще, чем глюкоза.
Биотехнология молочных продуктов[50] Спектр продуктов питания, получаемых при помощи микроорганизмов, обширен. Это продукты, получаемые в результате брожения — хлеб, сыр, вино, пиво, творог и так далее. До недавнего времени биотехнология использовалась в пищевой промышленности с целью усовершенствования освоенных процессов и более умелого использования микроорганизмов, но будущее здесь принадлежит генетическим исследованиям по созданию более продуктивных штаммов для конкретных нужд, внедрению новых методов в технологии брожения. Получение молочных продуктов в пищевой промышленности построено на процессах ферментации. Основой биотехнологии молочных продуктов является молоко. Молоко (секрет молочных желез) — уникальная естественная питательная среда. Она содержит 82–88 % воды и 12–18 % сухого остатка. В состав сухого молочного остатка входят белки (3,0–3,2 %), жиры (3,3–6,0 %), углеводы (молочный сахар лактоза — 4,7 %), соли (0,9–1 %), минорные компоненты (0,01 %): ферменты, иммуноглобулины, лизоцим и т. д. Молочные жиры очень разнообразны по своему составу. Основные белки молока — альбумин, казеин. Благодаря такому составу молоко представляет собой прекрасный субстрат для развития микроорганизмов. В сквашивании молока обычно принимают участие стрептококки и молочнокислые бактерии. Путем использования реакций, которые сопутствуют главному процессу сбраживания лактозы получают и другие продукты переработки молока: сметану, йогурт, сыр и т. д. Свойства конечного продукта зависят от характера и интенсивности реакций ферментации. Те реакции, которые сопутствуют образованию молочной кислоты, определяют обычно особые свойства продуктов. Например, вторичные реакции ферментации, идущие при созревании сыров, определяют вкус отдельных их сортов. В таких реакциях принимают участие пептиды, аминокислоты и жирные кислоты, находящиеся в молоке. Все технологические процессы производства продуктов из молока делятся на две части: 1) первичная переработка — уничтожение побочной микрофлоры; 2) вторичная переработка. Первичная переработка молока включает в себя несколько этапов. Сначала молоко очищается от механических примесей и охлаждается, чтобы замедлить развитие естественной микрофлоры. Затем молоко сепарируется (при производстве сливок) или гомогенизируется. После этого проводят пастеризацию молока, при этом температура поднимается до 80 °C, и оно закачивается в танки или ферментеры. Вторичная переработка молока может идти двумя путями: с использованием микроорганизмов и с использованием ферментов. С использованием микроорганизмов выпускают кефир, сметану, творог, простокваши, казеин, сыры, биофруктолакт, биолакт, с использованием ферментов — пищевой гидролизат казеина, сухую молочную смесь для коктейлей и т. д. При внесении микроорганизмов в молоко лактоза гидролизуется до глюкозы и галактозы, глюкоза превращается в молочную кислоту, кислотность молока повышается, и при pH 4–6 казеин коагулирует. Молочнокислое брожение бывает гомоферментативным и гетероферментативным. При гомоферментативном брожении основным продуктом является молочная кислота. При гетероферментативном брожении образуются диацетил (придающий вкус сливочному маслу), спирты, эфиры, летучие жирные кислоты. Одновременно идут протеолитические и липолитические процессы, что делает белки молока более доступными и обогащает дополнительными вкусовыми веществами. Для процессов ферментации молока используются чистые культуры микроорганизмов, называемые заквасками. Исключение составляют закваски для кефиров, которые представляют естественный симбиоз нескольких видов молочнокислых грибков и молочнокислых бактерий. Этот симбиоз в лабораторных условиях воспроизвести не удалось, поэтому поддерживается культура, выделенная из природных источников. При подборе культур для заквасок придерживаются следующих требований: — состав заквасок зависит от конечного продукта (например, для получения ацидофилина используется ацидофильная палочка, для производства простокваши — молочнокислые стрептококки); — штаммы должны отвечать определенным вкусовым требованиям; — продукты должны иметь соответствующую консистенцию, от ломкой крупитчатой до вязкой, сметанообразной; — определенная активность кислотообразования; — фагорезистентность штаммов (устойчивость к бактериофагам); — способность к синерезису (свойству сгустка отдавать влагу); — образование ароматических веществ; — сочетаемость штаммов (без антагонизма между культурами); — наличие антибиотических свойств, т. е. бактериостатическое действие по отношению к патогенным микроорганизмам; — устойчивость к высушиванию. Культуры для заквасок выделяются из природных источников, после чего проводится направленный мутагенез и отбор штаммов, отвечающих перечисленным выше требованиям. Биотехнологии на основе молока включают, как правило, все основные стадии биотехнологического производства, которые можно рассмотреть на примере сыроварения. Сыроварение — один из древнейших процессов, основанных на ферментации. Сыры бывают самые разнообразные — от мягких до твердых. Мягкие сыры содержат много воды, 50–60 %, а твердые — мало, 13–34 %. На первом этапе идет подготовка молока (первичная обработка). На втором — готовится культура молочнокислых бактерий. Микроорганизмы подбираются в определенной пропорции, обеспечивающей наилучшее качество. Набор бактерий также зависит от температуры термообработки. Третья стадия — стадия ферментации, — в сыроварении в некоторых случаях происходит в 2 этапа, до и после стадии выделения. Сначала молоко инокулируют определенными штаммами микроорганизмов, приводящими к образованию молочной кислоты, а также добавляют сычужный фермент реннин. Реннин ускоряет превращение жидкого молока в сгусток (створаживание) в несколько раз. Эта реакция активируется молочной кислотой, вырабатываемой бактериями. Функции реннина могут выполнять и другие протеиназы, но реннин также участвует в процессах протеолиза, происходящих в сыре при созревании. После образования сгустка сыворотку отделяют, а полученную творожистую массу подвергают термообработке и прессуют в формах. Далее сгусток солят и ставят на созревание. Иногда полученная масса происходит дополнительную обработку, которая заключается в следующем: заражение спорами голубых плесневых грибов при производстве рокфора; нанесение на поверхность спор белых плесневых грибов при производстве камамбера и бри; нанесение бактерий, необходимых для созревания некоторых сыров. Некоторые сыры после выделения должны подвергнуться дальнейшей ферментации (стадия созревания). Микроорганизмы и ферменты в ходе этого процесса гидролизуют жиры, белки и некоторые другие вещества молодого сыра. В результате их распада образуются вещества, придающие сырам характерный вкус. Процессы ферментации при производстве многих молочных продуктов, таких как сметана, творог, многие сыры идут в ферментерах открытого типа. Как правило, они занимают немного времени. К одним из самых простых относят производство кефира, простокваш, сметаны и масла. Например, при производстве сметаны к сливкам добавляют 0,5–1 % закваски, используемой при производстве масла. Далее продукт выдерживают, пока концентрация кислоты не достигнет 0,6 %. В заключение хотелось бы добавить, что процессы получения молочнокислых продуктов весьма просты и доступны для воспроизводства в домашних условиях. Они не требуют строгих условий соблюдения стерильности, протекают, как правило, при комнатной или чуть повышенной температуре. Собственно, изначально они были одними из первых "домашних" биотехнологий, которые были позднее поставлены на промышленную основу.
ОСНОВНЫЕ ТИПЫ БИОПРОЦЕССОВ
В настоящее время существуют следующие основные типы биопроцессов: — производство биомассы (например, белок одноклеточных); — клеточных компонентов (ферменты, нуклеиновые кислоты и т. д.) — метаболитов (химические продукты метаболической активности), включая первичные метаболиты, такие как этанол, молочная кислота; — вторичные метаболиты; — односубстратные конверсии (превращение глюкозы во фруктозу); — многосубстратные конверсии (обработка сточных вод, утилизация лигноцеллюлозных отходов).
Производство биомассы Человек традиционно получает белки, жиры и углеводы (основные компоненты пищи) из животных и растительных источников. Уже сегодня эти источники не покрывают все увеличивающиеся потребности человечества. Выяснилось, что белки и жиры микроорганизмов с успехом могут заменить белки и жиры традиционного происхождения. Преимущества микроорганизмов как продуцентов белка состоит в высоком содержании белка в биомассе и высокой скорости роста микроорганизмов. Термин белок одноклеточных (БОК) был предложен в 1966 г. для обозначения биомассы различных микроорганизмов (бактерий, дрожжей, грибов и водорослей). Кроме высокого содержания белка микробная биомасса содержит также жиры, нуклеиновые кислоты, витамины и минеральные компоненты. Источниками получения пищевого белка могут стать также белковые изоляты из различных видов зеленой биомассы, в т. ч. из табака. Для получения БОК используют самые разнообразные субстраты, включая парафины нефти, метан, водород, метанол, этанол, уксусную кислоту, углекислый газ, молочную сыворотку, мелассу, крахмал и целлюлозосодержащие отходы промышленности и сельского хозяйства. Для промышленного использования перспективными являются термофильные (растущие при высоких температурах до 50 °C) микроорганизмы. Качество биомассы оценивается по высокому содержанию белка, низкому содержанию нуклеиновых кислот и отсутствию вредных веществ.
Получение спиртов и полиолов Некоторые дрожжи и бактерии способны продуцировать этанол и бутанол, а также такие полиспирты как глицерин и 2,3-бутандиол. Эти продукты обычно синтезируют из нефти, однако микробное получение этанола и других спиртов вызывает все больший интерес. В России большая часть этанола получается микробиологическим путем из растительного сырья. Сдвиг в сторону получения микробиологического этанола наблюдается и в др. странах. Найдены бактерии Zymomonas mobilis[51], которые вдвое эффективнее сбраживают углеводы в этанол, чем дрожжи. В Бразилии производство топливного спирта вносит наибольший вклад в энергобаланс страны и составляет миллиарды литров. Ферментация мелассы различными видами Clostridium может быть использована для получения не только этанола, но и ацетальдегида, уксусной кислоты, этилацетата и диэтилового эфира. При изменении условий культивирования, например, в условиях щелочной среды вместо этанола образуется глицерин. Сырьем могут быть гидролизаты древесины, меласса, крахмал, молочная сыворотка. Отходы производства этанола содержат белки, углеводы, рибофлавин и др. витамины, и могут использоваться как кормовая добавка. Штаммы-продуценты первичных метаболитов получают путем индуцированного мутагенеза, так как в природе мутации, ведущие к сверхпродукции одного из метаболитов вредны. Нарушения в обмене веществ приводят к снижению конкурентоспособности и жизнеспособности микроорганизмов.
Производство вторичных метаболитов Из всех продуктов, получаемых с помощью микробных процессов, наибольшее значение имеют вторичные метаболиты. Вторичные метаболиты, называемые также идиолитами, — низкомолекулярные соединения, не требующиеся для роста в чистой культуре. Они производятся ограниченным числом таксономических групп и часто представляют собой смесь близкородственных соединений, относящихся к одной и той же химической группе. К вторичным метаболитам относятся антибиотики, алкалоиды, гормоны роста растений и токсины. Молекулы антибиотиков очень разнообразны по составу и механизму действия на микробную клетку. При этом в связи с возникновением устойчивости патогенных микроорганизмов к старым антибиотикам постоянно существует потребность в новых. В некоторых случаях природные микробные антибиотические продукты химическим или энзиматическим путем могут быть превращены в так называемые полусинтетические антибиотики, обладающие более высокими терапевтическими свойства- Микроорганизмы, производящие вторичные метаболиты, вначале проходят стадию быстрого роста, тропофазу, во время которой синтез вторичных веществ незначителен. По мере замедления роста из-за истощения одного или нескольких необходимых питательных веществ в культуральной среде микроорганизм переходит в идиофазу; именно в этот период синтезируются идиолиты. Эти особенности культурального роста необходимо учитывать при производстве. Например, в случае антибиотиков большинство микроорганизмов в процессе тропофазы чувствительно к собственным антибиотикам, однако во время идиофазы они становятся к ним устойчивыми. Чтобы уберечь микроорганизмы, продуцирующие антибиотики, от самоуничтожения, важно быстро достичь идиофазы и затем культивировать микроорганизмы в этой фазе. Это достигается путем варьирования режимов культивирования и составом питательной среды на стадиях быстрого и медленного роста.
Микробные биотрансформации Микроорганизмы способны осуществлять реакции трансформации, в которых те или другие соединения превращаются в новые продукты. Условия протекания этих реакций мягкие, и во многих случаях микробиологические трансформации предпочтительнее химических. Пример существующих крупномасштабных промышленных биоконверсий — производство уксуса из этанола, глюконовой кислоты из глюкозы. Широко используется микробная модификация стероидов, которые являются сложными полициклическими липидами. Теперь с использованием биоконверсии получают кортизон, гидрокортизон, преднизолон и целый ряд других стероидов. Применение и совершенствование микробной технологии в сотни раз снижает себестоимость производства стероидов.
Производство ферментов Получение ферментов с помощью микроорганизмов более выгодно, чем из растительных и животных источников. Микробные клетки продуцируют более 2 тысяч ферментов, катализирующих биохимические реакции, связанные с ростом, дыханием и образованием продуктов. Многие из этих ферментов могут быть выделены и проявляют свою активность независимо от клетки. В мире производится около 20 ферментов в объеме 65 тыс. тонн (а существует, как предполагают 25000 ферментов). Например, промышленным способом производят такие ферменты как амилаза, глюкоамилаза, протеаза, инвертаза, пектиназа, каталаза, стрептокиназа, целлюлаза и др. Амилазы и протеазы используют в текстильной, хлебопекарной и кожевенной промышленности. Пектолитические ферменты могут быть использованы для мацерации тканей при переработке растительного сырья, например при получении льноволокна. Щелочные протеазы, особенно иммобилизованные, очень эффективно используются в составе моющих средств. Кроме протеолитических ферментов в состав моющих средств вводят липазу, целлюлазу, оксидазу и амилазу для удаления загрязнений крахмального происхождения. Использование иммобилизованной глюкозоизомеразы для непрерывного получения глюкозы является наиболее крупным процессом такого рода в мире. Микробные ферменты активно используют в клинической диагностике при определении уровня холестерина в крови и мочевой кислоты. Ферменты предлагают использовать для очистки канализационных и водопроводных труб и т. д. и т. п. Ферменты для медицинских или аналитических целей должны быть высокоочищенными. Для повышения стабильности выделенных ферментов используют технику иммобилизации, т. е. связывания ферментов на поверхности нерастворимого в воде носителя, например, органических полимеров, стекла, минеральных солей, силикатов и т. п. Иммобилизованные ферменты (ИФ) можно длительное время использовать в биохимических реакторах в условиях непрерывного процесса. Примеры использования ИФ — изомеризация глюкозы во фруктозу, гидролиз белков, трансформация стероидов, гормонов и т. д. Новая область применения ИФ — создание на их основе бессеребряных фотоматериалов. На основе действия ферментов построены биолюминесцентные и иммуноферментные методы анализа, отличительной чертой которых является высокая чувствительность и абсолютная специфичность.
Аминокислоты, органические кислоты, витамины и другие биопродукты Производство аминокислот относится к одной из наиболее передовых областей биотехнологии. Аминокислоты получают путем химического синтеза или экстракцией из белковых гидролизатов. Незаменимые аминокислоты могут получаться микробиологическим путем более эффективно, чем путем химического синтеза. За рубежом 60 % мощностей по производству аминокислот занимают глутаминовая кислота, далее идут метионин, лизин и глицин. Глутаминовая кислота производится при участии в качестве продуцента штамма Corynebacterium. С помощью микроорганизмов можно получить до 60 органических кислот. Многие из них получаются в промышленном масштабе — итаконовая, молочная, уксусная, лимонная. Лимонная кислота во всем мире успешно производится с помощью гриба Aspergillus niger. Уксусную кислоту получают путем микробиологической конверсии водорода и углекислого газа бактериями Acetobacterium woodi и Clostridium асеticum. Витамины синтезируют в основном химическим путем или получают из естественных источников. Однако рибофлавин (В2), витамин В12 и аскорбиновую кислоту получают микробиологическим путем. Существует производство рибофлавина на основе использования дрожжеподобных грибов Eremothecium ashbyii и Ashbia gossypii. Рибофлавин продуцируется также видами Clostridium и Ascomycetes. Микроорганизмы являются также ценным источником получения никотиновой кислоты (витамин РР). Микроорганизмы являются источником получения липидов специального назначения с заранее определенными свойствами. Микробные жиры заменяют растительные (а в ряде случаев и превосходят) и могут использоваться в разных отраслях промышленности, с.-х., медицине. Микроорганизмы являются важным источником получения полимерных материалов на основе полисахаридов. Ценным микробным полисахаридом является декстран, образуемый бактериями рода Leucomonstoc. Декстран служит основой получения медицинских препаратов (кровезаменителей) и препаратов для биохимических исследований — сефадексов и др. молекулярных сит. Микроорганизмы являютсяисточником получения нуклеозидов и нуклеотидов и их производных, поверхностно-активных веществ и т. д.
Биоконверсия лигноцеллюлозных отходов Растительная биомасса — возобновляемый и легкодоступный источник сырья. Основные ее компоненты — целлюлоза (2/3), крахмал, гемицеллюлоза, лигнин. Лигнин — высокомолекулярный нерастворимый трехмерный неупорядоченный ароматический полимер. Целлюлоза — высокомолекулярный нерастворимый полимер глюкозы. Она является главным компонентом как растительной биомассы, так и сельскохозяйственных, бытовых отходов, а также отходов деревообрабатывающей и целлюлозно-бумажной промышленности. В основе биологической деградации лигноцеллюлозы лежит действие целлюлолитических ферментов. Реакционная способность природных целлюлозосодержащих материалов невелика, поэтому сырье для ферментативного осахаривания целлюлозы должно иметь большую поверхность, а микрофибриллярная структура целлюлозы должна быть разрушена. Реакционную способность природных субстратов также снижает наличие лигнина. Наиболее эффективным, а также дорогим и энергоемким способом предварительной подготовки сырья является размол. Поэтому для предобработки используют воздействие 0.5–2 % растворов щелочи, гамма-облучение, механо-термообработку в разбавленной серной кислоте с последующей экстракцией лигнина и др. методы. Гидролиз можно проводить и биологическим способом, с помощью ферментов, выделяемых грибами видов Trichoderma, Aspergillus, Sporotrichum. Далее при использовании дрожжей можно получить спирт, при использовании бактерий Klebsiella или Aeromonas — бутанол. Ряд микроорганизмов рода Clostridium могут продуцировать уксусную и молочную кислоты, лактат, ацетон из опилок, соломы, отходов сахарного тростника. С помощью Trichoderma reesii биомасса разлагается до сахаров. Ферменты и неразложившаяся целлюлоза поступают в повторные циклы, а остаточный лигнин используется в качестве источника энергии для перегонки спирта. Технология, разработанная в Арканзасском университете и используемая в промышленности нефтяной компанией «Галф ойл», заключается в одновременном осахаривании целлюлозы и сбраживании сахаров, полученных путем гидролиза. Для это го к смеси целлюлозной биомассы и дрожжей добавляют раствор целлюлаз. Остающийся лигнин также используется для перегонки в качестве топлива, но пентозы не сбраживаются. Фирма «Био фьюэл индастриз» из Ричмонда намерена построить в шт. Вирджиния фабрику, на которой в 1985 г. будет производиться 500 т этилового спирта в сутки из 2500 т целлюлозных отходов посредством этой технологии и целлюлаз из Trichoderma reesii. Третий вид технологии состоит в прямом сбраживании целлюлозными бактериями гексоз и пентоз, образующихся при гидролизе целлюлозы и гемицеллюлоз. Преимущества этой технологии, разработанной в лабораториях Массачусетского технологического института, заключаются в следующем: помимо одновременной конверсии целлюлоз и пентоз в этанол происходит комбинация целлюлозного и спиртового брожения, а, кроме того, необходимая предварительная обработка субстратов сводится к минимуму. При микробной деградации и конверсии целлюлоз и гемицеллюлоз можно получать этиловый спирт и сырье для химической промышленности (фурфурол, фенолы, крезолы). 200 000 т надлежащим образом переработанной соломы дают 50 000 т этанола и 20 000 т фурфурола. По оценкам некоторых специалистов, при микробной переработке целлюлозы можно получить до 30 % нефтехимикатов. Методы генной инженерии помогут создать штаммы, которые будут лучше адаптированы к этим типам конверсии и дадут больший выход. Это позволит разработать реальную стратегию замещения, которая станет эффективной после 2000 г. (к тому времени химия углерода придет на смену нефтехимии при производстве новых биополимеров, биорастворителей и биодетергентов). Перенос генов целлюлаз и гемицеллюлаз из Clostridium thermocellum в другие виды Clostridium позволит превращать целлюлозы и гемицеллюлозы в этиловый спирт, ацетон, бутанол, уксусную и молочную кислоты. Термофилия определенных штаммов Clostridium (при оптимальной температуре роста 65–75 °C) создает известные преимущества, так как стоимость перегонки этилового спирта и других растворителей уменьшится, а это сделает производственный процесс более экономичным. Исследователи из Университета Нового Южного Уэльса (Австралия) и Рутгерского университета (США) обнаружили, что бактерия Zymomonas mobilis, выделяемая из пальмового вина и мексиканского алкогольного напитка пульке, сбраживает сахара вдвое быстрее, чем дрожжи. Этот вид также подвергается геномной модификации, которая позволит разлагать целлюлозу с одновременным сбраживанием сахаров, получающихся в ходе деградации. В условиях строгого анаэробиоза можно осуществлять биометаногенез ароматических соединений. Этот процесс, надо полагать, широко распространен в природе, особенно в отходах и сточных водах, а также при конверсии некоторых биоцидов. По наблюдениям Ферри и Вольфа, в этом процессе участвуют несколько видов микробов, ответственных за различные стадии деградации ароматических колец до ацетата, который является одним из субстратов для метанобактерий (иными словами, его дегидрирование дает электроны, требующиеся для восстановления двуокиси углерода в метан). Среди бактерий видов превалируют, судя по всему, Methanobacterium formicicum и Methanospirillum hungati. Ферри и Вольфу удалось их вывести в чистые культуры. Бензольное кольцо сначала восстанавливается и затем разрезается на алифатические кислоты под действием грамотрицательных микроорганизмов. Последние превращаются в субстраты, используемые метанобактериями. Образующиеся электроны, вероятно, способствуют образованию водорода, который восстанавливает СО2 в СН4. Разложение бензольного кольца в метан в процессе анаэробиоза не является правилом. Например, в рубце жвачных животных бензоат и ароматические кислоты, получающиеся за счет деградации целлюлозы, не приводят к образованию метана; их можно обнаружить в моче и виде гиппуратов и других сходных соединений. В природных условиях ароматические соединения получаются при медленном разложении таннинов и лигнина главным образом благодаря внеклеточным микробным ферментам. Так как лигнины и таннины составляют значительную часть почвенного органического материала, метаногенез этих полимеров — важный процесс в углеродном цикле биосферы. Одним их отходов сельского хозяйства является солома. Эти отходы трудно использовать, так как скорость разложения соломы невелика. Лучшая утилизация — инокулирование её ассоциацией целлюлолитических грибов[52], азотфиксирующих и полисахаридообразующих бактерий. В таком виде солому можно запахивать в землю как органическое удобрение, а можно через определенное время использовать как высокобелковый витаминизированный корм.
ОБЪЕКТЫ БИОТЕХНОЛОГИИ И ИХ БИОТЕХНОЛОГИЧЕСКИЕ ФУНКЦИИ
Биотехнологические объекты находятся на разных ступенях организации: а) субклеточные структуры (вирусы, плазмиды, ДНК митохондрий и хлоропластов, ядерная ДНК); б) бактерии и цианобактерии; в) грибы; г) водоросли; д) простейшие; е) культуры клеток растений и животных; ж) растения — низшие (анабена-азолла) и высшие — рясковые. Субклеточные структуры будут подробно изучаться в разделе «Основы генетической инженерии», культуры растительных и животных клеток — в соответствующих разделах.
Бактерии и цианобактерии Микроорганизмов, синтезирующих продукты или осуществляющих реакции, полезные для человека, несколько сотен видов. Биотехнологические функции бактерий разнообразны. Бактерии используются при производстве: — пищевых продуктов, например, уксуса (Gluconobacter suboxidans), молочнокислых напитков (Lactobacillus, Leuconostoc) и др.; — микробных инсектицидов (Bacillus thuringiensis); — белка (Methylomonas); — витаминов (Clostridium — рибофла вин); — растворителей и органических кислот; — биогаза и фотоводорода. Полезные бактерии относятся к эубактериям[53]. Уксуснокислые бактерии, представленные родами Gluconobacter и Acetobacter, — это грамотрицательные бактерии, превращающие этанол в уксусную кислоту, а уксусную кислоту в углекислый газ и воду. Род Bacillus относится к грамположительным бактериям, которые способны образовывать эндоспоры и имеют перитрихиальное жгутикование. B.subtilis — строгий аэроб, а В. thuringiensis может жить и в анаэробных условиях. Анаэробные, образующие споры бактерии представлены родом Clostridium. C.acetobutylicum сбраживает сахара в ацетон, этанол, изопропанол и п-бутанол (ацетобутаноловое брожение), другие виды могут также сбраживать крахмал, пектин и различные азотсодержащие соединения. К молочнокислым бактериям относятся представители родов Lactobacillus, Leuconostoc и Streptococcus, которые не образуют спор, грамположительны и нечувствительны к кислороду. Гетероферментативные молочнокислые бактерии рода Leuconostoc превращают углеводы в молочную кислоту, этанол и углекислый газ. Гомоферментативные молочнокислые бактерии рода Streptococcus продуцируют только молочную кислоту, а брожение, осуществляемое представителями рода Lactobacillus, позволяет получить наряду с молочной кислотой ряд разнообразных продуктов. К бактериям рода Corynebacterium, неподвижные грамположительные клетки которых не образуют эндоспор, относятся патогенные (С. diphtheriae, С. tuberculosis) и непатогенные почвенные виды, имеющие промышленное значение. С.glutamicum служит источником лизина и улучшающих вкус нуклеотидов. Корине-бактерии хотя и считаются факультативными анаэробами, лучше растут аэробно. Бактерии используются для микробного выщелачивания руд и утилизации горнорудных отходов. Широко используется такое свойство некоторых бактерий, как диазотрофность, то есть способность к фиксации атмосферного азота. Выделяют 2 большие группы диазотрофов: — симбионты: без корневых клубеньков (азотобактер — лишайники, азоспириллум — лишайники, анабена — лишайники, азолла), с корневым клубеньками (бобовые — ризобии, ольха, лох, облепиха — актиномицеты); — свободноживущие: гетеротрофы (азотобактер, клостридиум, метилобактер), автотрофы (хлоробиум, родоспириллум и амебобактер). Микробные клетки используют для трансформации веществ. К архебактериям попали, например, метаногены. Естественно, что упомянуто только небольшое количество используемых родов бактерий. Бактерии также широко используются в генноинженерных манипуляциях при создании геномных клонотек, введении генов в растительные клетки (агробактерии). Производственные штаммы микроорганизмов должны соответствовать определенным требованиям: способность к росту на дешевых питательных средах, высокая скорость роста и образования целевого продукта, минимальное образование побочных продуктов, стабильность продуцента в отношении производственных свойств, безвредность продуцента и целевого продукта для человека и окружающей среды. В связи с этим все микроорганизмы, используемые в промышленности проходят длительные испытания на безвредность для людей, животных и окружающей среды. Важным свойством продуцента является устойчивость к инфекции, что важно для поддержания стерильности, и фагоустойчивость. Все цианобактерии обладают способностью к азотфиксации, что делает их весьма перспективными продуцентами белка. Анабена (Anabaena) — нитчатая сине-зеленая водоросль. Нити из более или менее округлых клеток, содержат гетероцисты и иногда крупные споры, по всей длине нить одинаковой толщины. В цитоплазме клеток откладывается близкий к гликогену запасной продукт — анабенин. Такие представители цианобактерий, как носток, спирулина, триходесмиум съедобны и непосредственно употребляются в пищу. Носток образует на бесплодных землях корочки, которые разбухают при увлажнении. В Японии местное население использует в пищу пласты ностока, образующиеся на склонах вулкана и называет их ячменным хлебом Тенгу (Тенгу — добрый горный дух). Свое шествие спирулина (Spirulina platensis) начала из Африки — население района озера Чад давно употребляет ее в пищу, называя этот продукт «дихе». Другое место, откуда начала распространяться спирулина, но иного вида (Spirulina maxima) — воды озера Тескоко в Мексике. Еще ацтеки собирали с поверхности озер и употребляли в пищу слизистую массу сине-зеленой водоросли спирулины. Впервые галеты "текуитлатл" упомянуты испанцем Кастильо в 1521 г. Эти галеты продавались на базаре в Мехико и состояли из высушенных слоев S.maxima. В 1964 году бельгийский ботаник Ж.Леонар обратил внимание на галеты сине-зеленого цвета, которые местное население изготовляло из водорослей, растущих в щелочных прудах вокруг озера Чад. Эти галеты представляли собой высушенную массу спирулины. Анализ образцов Spirulina показал, что в ней содержится 65 % белков (больше, чем в соевых бобах), 19 % углеводов, 6 % пигментов, 4 % липидов, 3 % волокон и 3 % золы. Для белков этой водоросли характерно сбалансированное содержание аминокислот. Клеточная стенка этой водоросли хорошо переваривается. Как озеро Тескоко, так и водоемы района озера Чад имеют в воде очень высокое содержание щелочей. Характерно, что в таких озерах спирулина полностью доминирует и растет почти как монокультура — составляет в отдельных озерах до 99 % общего количества водорослей. Растет спирулина в щелочной среде при pH вплоть до 11. Ее собирают также из озер около г. Мехико, получая до 2 т сухого веса биомассы водоросли в сутки, и эта продукция рассылается в США, Японию, Канаду. В других странах спирулину культивируют обычно в искусственных водоемах или специальных емкостях. Спирулину можно культивировать в открытых прудах или, как в Италии, в замкнутой системе из полиэтиленовых труб. Урожайность очень высокая: получают до 20 г сухой массы водоросли с 1 м2 в день, а расчеты на год показали, что она превысит выход пшеницы примерно в 10 раз. Преимущества спирулины по сравнению с другими съедобными водорослями не только в простоте культивирования, но и в несложности сбора биомассы, высушивания ее, например, под солнцем. В ряде стран выращивают спирулину вида Spirulina platensis. Недавно было показано, что в клетках спирулины, помимо ценного белка, углеводов, липидов, витаминов, в значительных количествах запасается, например, такое ценное вещество, как поли-Ь-оксибутират. Отечественная фармацевтическая промышленность выпускает препарат «Сплат» на основе цианобактерии Spirulina platensis. Он содержит комплекс витаминов и микроэлементов и применяется как общеукрепляющее и иммуностимулирующе средство.
Грибы Биотехнологические функции грибов разнообразны. Их используют для получения таких продуктов, как: — антибиотики (пенициллы, стрептомицеты, цефалоспорины); — гиббереллины и цитокинины (физариум и ботритис); — каротиноиды (н-р, астаксантин, придающий мякоти лососевых рыб краснооранжевый оттенок вырабатывают Rhaffia rhodozima, которых добавляют в корм на рыбозаводах); — белок (Candida, Saccharomyces lipolitica); — сыры типа рокфор и камамбер (пенициллы); — соевый соус (Aspergillus oryzae). К грибам относятся[54] актиномицеты, дрожжи и плесени. Истинные актиномицеты — строгие аэробы, они грамположительны и не образуют спор. Наиболее представительный в этой группе — род Streptomyces, отдельные виды которого продуцируют широко применяемые антибиотики. При росте на твердых средах актиномицеты образуют очень тонкий мицелий с воздушными гифами, которые дифференцируются в цепочки конидиоспор. Каждая конидиоспора способна образовать микроколонию. Антибиотики продуцирует и другой вид актиномицетов, Micromonospora, колонии которого лишены воздушных гиф и образуют конидиоспоры непосредственно на мицелии. Из 500 известных видов дрожжей первым люди научились использовать Saccharomyces cerevisiae, этот вид наиболее интенсивно культивируется. К дрожжам, сбраживающим лактозу, относится Kluyveromyces fragilis, который используют для получения спирта из сыворотки. Saccharomycopsis lipolytica деградирует углеводороды и употребляется для получения белковой массы. Все три вида принадлежат к классу аскомицетов. Другие полезные виды относятся к классу дейтеромицетов (несовершенных грибов), так как они размножаются не половым путем, а почкованием. Candida utilis растет в сульфитных сточных водах (отходы бумажной промышленности). Trichosporon cutaneum, окисляющий многочисленные органические соединения, включая некоторые токсичные (например, фенол), играет важную роль в системах аэробной переработки стоков. Phaffia rhodozyma синтезирует астаксантин — каротиноид, который придает мякоти форели и лосося, выращиваемых на фермах, характерный оранжевый или розоватый цвет. Промышленные дрожжи обычно не размножаются половым путем, не образуют спор и полиплоидны. Последним объясняется их сила и способность адаптироваться к изменениям среды культивирования (в норме ядро клетки S.cerevisiae содержит 17 или 34 хромосомы, т. е. клетки либо гаплоидны, либо диплоидны). Плесени вызывают многочисленные превращения в твердых средах, которые происходят пред брожением. Их наличием объясняется гидролиз рисового крахмала при производстве сакэ и гидролиз соевых бобов, риса и солода при получении пищи, употребляемой в азиатских странах. Пищевые продукты на основе сброженных плесневыми грибами Rhizopus oligosporus соевых бобов или пшеницы содержат в 5–7 раз больше таких витаминов, как рибофлавин, никотиновая кислота) и отличаются повышенным в несколько раз содержанием белка. Плесени также продуцируют ферменты, используемые в промышленности (амилазы, пектиназы и т. д.), органические кислоты и антибиотики. Их применяют и в производстве сыров, например, камамбера и рокфора. Искусственное выращивание грибов способно внести и иной, не менее важный вклад в дело обеспечения продовольствием возрастающего населения земного шара. Люди употребляют грибы в пищу с глубокой древности. Поэтому сделать грибы такой же управляемой сельскохозяйственной культурой, как зерновые злаки, овощи, фрукты, давно уже стало актуальной задачей. Наиболее легко поддаются искусственному выращиванию древоразрушающие грибы. Это связано с особенностями их биологии, которые стали нам известны и понятны только сейчас. Их способность легко расти и плодоносить, использовали с древнейших времен. Искусственное разведение древоразрушающих грибов получило довольно широкое распространение. Мицелий съедобных грибов можно выращивают на жидких средах, например, на молочной сыворотке и др., в специальных ферментерах, в так называемой глубинной культуре. Это полностью механизированный и автоматизированный процесс. Так, в Институте микробиологии Академии наук БССР разработаны и апробированы в опытном производстве способы получения белковых грибных препаратов даедалина и пантегрина из мицелия древоразрушающих грибов дедалеопсиса бугристого и пилолистника тигрового, с высоким содержанием белка и биологически активных веществ. По содержанию белка 1 кг этих препаратов эквивалентен 2 кг мяса. По биологической ценности белок этих препаратов не уступает растительным и приближается к животным белкам. Перевариваемость белков данных препаратов составляет свыше 80 %. В основе этого способа получения пищевого белка лежат полученные микологами данные о том, что плодовые тела грибов и их грибница близки по своему химическому составу и пищевой ценности. Грибные белковые препараты даедалин и пантегрин рекомендованы в качестве пищевых добавок после соответствующего медицинского контроля. Исследования в этом направлении продолжаются.
Простейшие Простейшие относятся к числу нетрадиционных объектов биотехнологии. До недавнего времени они использовались лишь как компонент активного ила при биологической очистке сточных вод. В настоящее время они привлекли внимание исследователей как продуценты биологически активных веществ. В этом качестве рациональнее использовать свободноживущих простейших, обладающих разнообразными биосинтетическими возможностями и потому широко распространенными в природе. Особую экологическую нишу занимают простейшие, обитающие в рубце жвачных животных. Они обладают ферментом целлюлазой, способствующей разложению клетчатки в желудке жвачных. Простейшие рубца могут быть источником этого ценного фермента. Возбудитель южноамериканского трипаносомоза — Trypanosoma (Schizotrypanum cruzi) стала первым продуцентом противоопухолевого препарата круцина (СССР) и его аналога — трипанозы (Франция). Изучая механизм действия этих препаратов, советские ученые (Г. И. Роскин, Н. Г. Клюева и их сотрудники), а также их французские коллеги (Ж. Кудер, Ж. Мишель-Брэн и др.) пришли к выводу, что эти препараты оказывают цитотоксический эффект при прямом контакте с опухолью и ингибируют ее опосредованно, путем стимуляции ретикулоэндотелиальной системы. Выяснилось, что ингирующее действие связано с жирнокислотными фракциями. Характерной особенностью этих организмов является высокое содержание ненасыщенных жирных кислот, составляющее у трипаносомид 70–80 %, а у Astasia longa (свободноживущий жгутиконосец) — 60 % от суммы всех жирных кислот. У жгутиконосцев фосфолипиды и полиненасыщенные жирные кислоты имеют такой же состав и строение, как в организме человека и животных. В мире микробов полиненасыщенные жирные кислоты не синтезируются, а многоклеточные животные или растения представляют собой более ограниченную сырьевую базу, чем простейшие, культуры которых можно получать методами биотехнологии независимо от времени года или климатических условий. Поскольку липидный метаболизм простейших обладает относительной лабильностью, были изучены пути его регуляции. Применение к простейшим общепринятого в микробиологии приема повышения биосинтеза липидов за счет снижения содержания в среде источника азота и увеличения содержания источника углерода привело к резкому торможению или остановке роста культур. Для создания условий направленного биосинтеза липидов в среды для культивирования жгутиконосцев добавляли предшественники и стимуляторы биосинтеза липидов: малонат, цитрат, сукцинат, цитидиннуклеотиды в сочетании с определенным режимом аэрации. Российские ученые получили водорастворимый полусинтетический препарат — астазилид, представляющий собой комплекс эфиров сахарозы и жирных кислот, предварительно выделенных из A. longa. Для изучения активности и механизма действия этого препарата были применены различные модели: бислойные липидные мембраны (БЛМ), монослойные культуры почки теленка и карциномы яичника человека, иммунокомпетентные клетки — перитонеальные макрофаги. Было установлено, что астазилид вызывает увеличение проводимости, поверхностного натяжения, а также уменьшение электромеханической стабильности БЛМ. Полученные данные позволяют предполагать, что в основе физиологических эффектов препарата лежит его значительное мембраноактивное действие. Астазилид проявляет мягкие детергентные свойства. Возможно, что увеличение проводимости и некоторая дестабилизация клегочных мембран открывают путь для проникновения внутрь клетки Са2+ и других ионов, играющих ключевую роль в регуляции метаболизма. При изучении действия астазилида на культуру клеток почки теленка было установлено, что препарат увеличивает митотический индекс клеток, снижает их полиморфизм, улучшает адгезивные свойства культуры, обеспечивает более плотное сцепление с субстратом и усиление межклеточных контактов. Препарат не обладал прямым цитотоксическим действием на культуру опухолевых клеток, а его противоопухолевое действие, изученное на 8 штаммах перевиваемых опухолей мышей и крыс, реализовалось через иммунную систему. Астазилид действовал главным образом на клеточное звено иммунитета, вызывая повышение фагоцитарной активности перитонеальных макрофагов, увеличение способности индуцировать развитие гиперчувствительности замедленного типа и некоторых других показателей. Препарат предотвращал гибель 60–80 % животных, зараженных бакте риальными инфекциями (Е. coli, Ps. aerugenosa), а также лейшманиями. Другой группой биологически активных веществ простейших являются полисахариды. Разнообразие полисахаридов, синтезируемых простейшими, достаточно велико. Особый интерес представляет парамилон, характерный для эвгленоидных жгутиконосцев. Представители родов Astasia и Euglena способны к сверхсинтезу парамилона, составляющему свыше 50 % сухого остатка клеток. Этот полисахарид изучается как стимулятор иммунной системы млекопитающих. В наших опытах парамилон A. longa обладал выраженным противоопухолевым эффектом. Действуя опосредованно через иммунную систему, парамилон тормозит рост саркомы 180 на 60 % и снижает прививаемость аденокарциномы Эрлиха. Аденокарцинома Эрлиха вообще не прививалась у 50–60 % мышей, которым профилактически был введен парамилон в дозах 3 и 30 мг/кг веса животного. Парамилон, выделенный из A. longa, практически нетоксичен. Выраженное иммуномодулирующее действие и низкая токсичность этого препарата являются предпосылкой для его углубленного исследования в сочетании с препаратами прямого противоопухолевого действия, радиотерапией и другими адъювантами. В настоящее время в мире придается большое значение производству глюканов не только для медицинских целей, но и для пищевой и текстильной промышленности. До сих пор глюканы получали из культур бактерий или морских водорослей. Эвглениды являются одним из наиболее перспективных источников этого вещества. Структурные полисахариды, входящие в состав клеточных мембран простейших, — это гетерополисахариды, содержащие глюкозу, маннозу, ксилозу, арабинозу, рибозу, галактозу, рамнозу, фруктозу, глюкозамин. Наиболее характерными гетерополисахаридами являются арабиногалактаны, Д-галакто-Д-маннан, фосфаноглюканы и другие. Большой интерес представляет выяснение антигенной взаимосвязи между непатогенными и патогенными для человека видами трипаносомид. Установлено, что при введении мышам полисахаридов из культур непатогенных для человека простейших — Herpetomonas sp. и Crithidia fasciculata — повышалась резистентность животных к Т. cruzi, возбудителю болезни Чагаса у человека. Наличие перекрестных иммунологических реакций между полисахаридами различных типов послужило основанием для вывода о том, что антигенная общность между этими веществами обусловлена не структурой полимера, а отдельными мономерами или олигомерами одинакового химического строения. Биомасса простейших содержит до 50 % белка. Его высокая биологическая ценность заключается в том, что он содержит все незаменимые аминокислоты, причем со держание свободных аминокислот на порядок выше, чем в биомассе микроводорослей, бактерий и в мясе. Это свидетельствует о широких возможностях применения свободноживущих простейших в качестве источника кормового белка.
Водоросли Водоросли используются, в основном, для получения белка. Весьма перспективны в этом отношении и культуры одноклеточных водорослей, в частности высокопродуктивных штаммов рода Chlorella и Scenedesmus. Их биомасса после соответствующей обработки используется в качестве добавки в рационы скота, а также в пищевых целях. Одноклеточные водоросли выращивают в условиях мягкого теплого климата (Средняя Азия, Крым) в открытых бассейнах со специальной питательной средой. К примеру, за теплый период года (6–8 месяцев) можно получить 50–60 т биомассы хлореллы с 1 га, тогда как одна из самых высокопродуктивных трав — люцерна дает с той же площади только 15–20 т урожая. Хлорелла содержит около 50 % белка, а люцерна — лишь 18 %. В целом в пересчете на 1 га хлорелла образует 20–30 т чистого белка, а люцерна — 2–3,5 т. Кроме того, хлорелла содержит 40 % углеводов, 7-10 % жиров, витамины А (в 20 раз больше), В2, К, РР и многие микроэлементы. Варьируя состав питательной среды, можно процессы биосинтеза в клетках хлореллы сдвинуть в сторону накопления либо белков, либо углеводов, а также активировать образование тех или иных витаминов. При завоевании племен майя миссионерами описывался случай, когда испанцы около полутора лет осаждали крепость на вершине горы. Естественно, что все продукты давно должны были кончиться, однако крепость не сдавалась. Когда же она была наконец взята, то испанцы с удивлением увидели в ней небольшие пруды, где культивировались одноклеточные водоросли, из которых индейцы готовили особый сыр. Испанцы попробовали его и нашли весьма приятным на вкус. Однако это было уже после того, как испанцы уничтожили абсолютно всех защитников и секрет племени был утерян. В наше время делались попытки определить этот вид водорослей, из которых готовился сыр, но они не увенчались успехом. В пищу употребляют не менее 100 видов макрофитных водорослей как в странах Европы и Америки, так и особенно на Востоке. Из них готовят много разнообразных блюд, в том числе диетических, салатов, приправ. Их подают в виде засахаренных кусочков, своеобразных конфет, из них варят варенье, делают желе, добавки к тесту и многое другое. В магазине можно купить консервы из морской капусты — ламинарии дальневосточных или северных морей. Ее консервируют с мясом, рыбой, овощами, рисом, употребляют при приготовлении супов и др. Она наряду с микроводорослью хлореллой является самой популярной съедобной и кормовой водорослью. Известны и другие съедобные макрофитные водоросли — ульва, из которой делают разные зеленые салаты, а также алария, порфира, родимения, хондрус, ундария и др. В Японии продукты, получаемые из ламинариевых, называют «комбу», и для того, чтобы их вкусно приготовить, существует более десятка способов. В целом ряде стран водоросли используют как весьма полезную витаминную добавку к кормам для сельскохозяйственных животных. Их прибавляют к сену или дают как самостоятельный корм для коров, лошадей, овец, коз, домашней птицы во Франции, Шотландии, Швеции, Норвегии, Исландии, Японии, Америке, Дании и на нашем Севере. Животным скармливают в виде добавки также биомассу выращиваемых микроводорослей (хлорелла, сценедесмус, дуналиелла и др.). Гидролизаты белка зеленой водоросли Scenedesmus используются в медицине и косметической промышленности. В Израиле на опытных установках проводятся эксперименты с зеленой одноклеточной водорослью Dunaliella bardawil, которая синтезирует глицерол. Эта водоросль относится к классу равножгутиковых и похожа на хламидомонаду. Dunadiella может расти и размножаться в среде с широким диапазоном содержания соли: и в воде океанов, и в почти насыщенных солевых растворах Мертвого моря. Она накапливает свободный глицерол, чтобы противодействовать неблагоприятному влиянию высоких концентраций солей в среде, где она растет. При оптимальных условиях и высоком содержании соли на долю глицерола приходится до 85 % сухой массы клеток. Для роста этим водорослям необходимы: морская вода, углекислый газ и солнечный свет. После переработки эти водоросли можно использовать в качестве корма для животных, так как у них нет неперевариваемой клеточной оболочки, присущей другим водорослям. Они также содержат значительное количество β-каротина. Таким образом, культивируя эту водоросль, можно получать глицерол, пигмент и белок, что весьма перспективно с экономической точки зрения. Наряду с кормами водоросли давно применяют в сельском хозяйстве в качестве удобрений. Биомасса обогащает почву фосфором, калием, йодом и значительным количеством микроэлементов, пополняет также ее бактериальную, в том числе азотфиксирующую, микрофлору. При этом в почве водоросли разлагаются быстрее, чем навозные удобрения, и не засоряют ее семенами сорняков, личинками вредных насекомых, спорами фитопатогенных грибов. Одним из самых ценных продуктов, получаемых из красных водорослей, является агар — полисахарид, присутствующий в их оболочках и состоящий из агарозы и агаропектина. Количество его доходит до 30–40 % от веса водорослей (водоросли лауренция и грацилярия, гелидиум). Водоросли — единственный источник получения агара, агароидов, каррагинина, альгинатов. В мире в 1980 г. было получено 7 тыс. т агара, 222 тыс. т альгинатов, 10 тыс. т каррагинина. В нашей стране основным источником агара служит красная водоросль анфельция. Бурые водоросли являются единственным источником получения одних из самых ценных веществ водорослей — солей альгиновой кислоты, альгинатов. Альгиновая кислота — линейный гетерополисахарид, построенный из связанных остатков β-Д-маннуроновой и α-L-гиулуроновой кислот. Альгинаты исключительно широко применяются в народном хозяйстве. Это изготовление высококачественных смазок для трущихся деталей машин, медицинские и парфюмерные мази и кремы, синтетические волокна и пластики, стойкие к любой погоде лакокрасочные покрытия, не выцветающие со временем ткани, производство шелка, клеящих веществ исключительно сильного действия, строительных материалов, пищевые продукты отличного качества — фруктовые соки, консервы, мороженое, стабилизаторы растворов, брикетирование топлива, литейное производство и многое другое. Альгинат натрия — наиболее используемое соединение — способен поглощать до 300 весовых единиц воды, образуя при этом вязкие растворы. Бурые водоросли богаты также весьма полезным соединением — шестиатомным спиртом маннитом, который с успехом применяют в пищевой промышленности, фармацевтике, при производстве бумаги, красок, взрывчатки и др. Бурые водоросли в ближайшее время планируется использовать для получения биогаза. Каллусные культуры макрофитных водорослей могут быть использованы далее в различных направлениях. В случае, если они получены от агарофитов, можно непосредственно получать из них агар. Каллусные культуры пищевых макрофитных водорослей, например ламинариевых, могут в перспективе использоваться для получения белка, непосредственно идущего в пищу и в пищевые добавки, а также в корма сельскохозяйственным животным. Суспензионные культуры макрофитных водорослей открывают в перспективе возможности использования их в качестве трофического звена в марикультуре. Они могли бы также выступать в качестве партнера в искусственно создаваемых растительных ассоциациях, участники которых обладают полезными свойствами. Выделяемые клетками культуры экзометаболиты, характерные для исходного вида водоросли, будут составлять основу трофического обмена при удачном подборе партнеров в растительной ассоциации или комплексе марикультуры. Необходимо отметить, что при отсутствии токсического и антагонистического действия выделяемых соединений в естественных условиях существуют разнообразные и многочисленные природные ассоциации, например, повсеместно встречающиеся комплексы водорослей и бактерий.
Растения Водный папоротник азолла ценится как органическое азотное удобрение, так как растет в тесном симбиозе с сине-зеленой водорослью анабена. Это позволяет симбиотическому организму анабена-азолла накапливать много азота в вегетативной массе. Анабену-азоллу выращивают на рисовых полях перед посевом риса, что позволяет снижать количество вносимых минеральных удобрений. Представители семейства рясковых (Lemnaceae) — самые мелкие и простые по строению цветковые растения, величина которых редко превышает 1 см. Рясковые — свободноживущие водные плавающие растения. Вегетативное тело напоминает лист или слоевище низших растений, поэтому до начала 18 века ряску относили к слоевищным растениям. В литературе встречается несколько названий тела рясковых. Самое удачное — листец. Тело рясковых — особая структура, не дифференцированная на листья и стебель (листоветвь), представляющая зеленую пластинку, иногда выпуклую с нижней стороны. Рясковые (Lemna minor, L. trisulca, Wolfia, Spirodela polyrhiza) служат кормом для животных, для уток и других водоплавающих птиц, рыб, ондатры. Их используют и в свежем, и в сухом виде как ценный белковый корм для свиней и домашней птицы. Рясковые содержат много протеина (до 45 % от сухой массы). 45 % углеводов, 5 % жиров и остальное — клетчатка и т. д. Они высоко продуктивны, неприхотливы в культуре, хорошо очищают воду и обогащают её кислородом. Это делает рясковые ценным объектом для морфогенетических, физиологических и биохимических исследований.
ПЕРСПЕКТИВЫ РАЗВИТИЯ БИОТЕХНОЛОГИИ
Центральная проблема биотехнологии — интенсификация биопроцессов как за счет повышения потенциала биологических агентов и их систем, так и за счет усовершенствования оборудования, применения биокатализаторов (иммобилизованных ферментов и клеток) в промышленности, аналитической химии, медицине. В основе промышленного использования достижений биологии лежит техника создания рекомбинантных молекул ДНК. Конструирование нужных генов позволяет управлять наследственностью и жизнедеятельностью животных, растений и микроорганизмов и создавать организмы с новыми свойствами. В частности, возможно управление процессом фиксации атмосферного азота и перенос соответствующих генов из клеток микроорганизмов в геном растительной клетки. В качестве источников сырья для биотехнологии все большее значение будут приобретать воспроизводимые ресурсы не пищевых растительных материалов, отходов сельского хозяйства, которые служат дополнительным источником как кормовых веществ, так и вторичного топлива (биогаза), органических удобрений. Одной из бурно развивающихся отраслей биотехнологии считается технология микробного синтеза ценных для человека веществ. По прогнозам, дальнейшее развитие этой отрасли повлечет за собой перераспределение ролей растениеводства и животноводства с одной стороны, и микробного синтеза — с другой, в формировании продовольственной базы человечества. Не менее важным аспектом современной микробиологической технологии является изучения участия микроорганизмов в биосферных процессах и направленная регуляция их жизнедеятельности с целью решения проблемы охраны окружающей среды от техногенных, сельскохозяйственных и бытовых загрязнений. С этой проблемой тесно связаны исследования по выявлению роли микроорганизмов в плодородии почв (гумусообразовании и пополнении запасов биологического азота), борьбе с вредителями и болезнями сельскохозяйственных культур, утилизации пестицидов и др. химических соединений в почве. Имеющиеся в этой области знания свидетельствуют о том, что изменение стратегии хозяйственной деятельности человека от химизации к биологизации земледелия оправдывается как с экономической, так и с экологической точек зрения. В данном направлении перед биотехнологией может быть поставлена цель регенерации ландшафтов. Ведутся работы по созданию биополимеров, которые будут способны заменить современные пластмассы. Эти биополимеры имеют существенное преимущество перед традиционными материалами, так как нетоксичны и подвержены биодеградации, то есть легко разлагаются после их использования, не загрязняя окружающую среду. Биотехнологии, основанные на достижениях микробиологии, наиболее экономически эффективны при комплексном их применении и создании безотходных производств, не нарушающих экологического равновесия. Их развитие позволит заменить многие огромные заводы химической промышленности экологически чистыми компактными производствами. Важным и перспективным направлением биотехнологии является разработка способов получения экологически чистой энергии. Получение биогаза и этанола были рассмотрены выше, но есть и принципиально новые экспериментальные подходы в этом направлении. Одним из них является получение фотоводорода. Если из хлоропластов выделить мембраны, содержащие фотосистему, то на свету происходит фотолиз воды — разложение на кислород и водород. Моделирование процессов фотосинтеза, происходящих в хлоропластах, позволило бы запасать энергию Солнца в ценном топливе — водороде. Преимущества такого способа получения энергии очевидны: • наличие избытка субстрата, воды; • нелимитируемый источник энергии — Солнце; • продукт (водород) можно хранить, не загрязняя атмосферу; • водород имеет высокую теплотворную способность (29 ккал/г) по сравнению с углеводородами (3.5 ккал/г); • процесс идет при нормальной температуре без образования токсических промежуточных продуктов; • процесс циклический, так как при потреблении водорода регенерируется субстрат — вода. Другой механизм превращения энергии у галофитных бактерий Halobacterium halobium, которые используют энергию солнца, поглощаемую пурпурным пигментом бактериородопсином, находящимся в мембране клетки. Поглощение света вызывает химические и физические изменения в мембране, приводящие к направленному транспорту протонов водорода с одной стороны мембраны на другую и созданию электрохимического градиента. Следствием этого является синтез аденозинтрифосфорной кислоты. Н.halobium можно культивировать в мелких водоемов с высоким содержанием NaCl и других минеральных солей. Из 10 литров бактериальной культуры можно получить 0,5 грамма мембран, содержащих до 100000 молекул пигмента. Пигмент можно фиксировать на подложках, обладающих физическими и химическими свойствами для транспорта протонов.
ОСНОВНЫЕ ПРИНЦИПЫ ПРОМЫШЛЕННОГО ОСУЩЕСТВЛЕНИЯ БИОТЕХНОЛОГИЧЕСКИХ ПРОЦЕССОВ
Стадии биотехнологического производства Большое разнообразие биотехнологических процессов, нашедших промышленное применение, приводит к необходимости рассмотреть общие, наиболее важные проблемы, возникающие при создании любого биотехнологического производства. Процессы промышленной биотехнологии разделяют на 2 большие группы: производство биомассы и получение продуктов метаболизма. Однако такая классификация не отражает наиболее существенных с технологической точки зрения аспектов промышленных биотехнологических процессов. В этом плане необходимо рассматривать стадии биотехнологического производства, их сходство и различие в зависимости от конечной цели биотехнологического процесса. В общем виде система биотехнологического производства продуктов микробного синтеза представлена на рис. 1. Существует 5 стадий биотехнологического производства. Две начальные стадии включают подготовку сырья и биологически действующего начала. В процессах инженерной энзимологии они обычно состоят из приготовления раствора субстрата сзаданными свойствами (pH, температура, концентрация) и подготовки партии ферментного препарата данного типа, ферментного или иммобилизованного. При осуществлении микробиологического синтеза необходимы стадии приготовления питательной среды и поддержания чистой культуры, которая могла бы постоянно или по мере необходимости использоваться в процессе. Поддержание чистой культуры штамма-продуцента — главная задача любого микробиологического производства, поскольку высокоактивный, не претерпевший нежелательных изменений штамм может служить гарантией получения целевого продукта с заданными свойствами. Третья стадия — стадия ферментации, на которой происходит образование целевого продукта. На этой стадии идет микробиологическое превращение компонентов питательной среды сначала в биомассу, затем, если это необходимо, в целевой метаболит. На четвертом этапе из культуральной жидкости выделяют и очищают целевые продукты. Для промышленных микробиологических процессов характерно, как правило, образование очень разбавленных растворов и суспензий, содержащих, помимо целевого, большое количество других веществ. При этом приходится разделять смеси веществ очень близкой природы, находящихся в растворе в сравнимых концентрациях, весьма лабильных, легко подвергающихся термической деструкции. Заключительная стадия биотехнологического производства — приготовление товарных форм продуктов. Общим свойством большинства продуктов микробиологического синтеза является их недостаточная стойкость к хранению, поскольку они склонны к разложению и в таком виде представляют прекрасную среду для развития посторонней микрофлоры. Это заставляет технологов принимать специальные меры для повышения сохранности препаратов промышленной биотехнологии. Кроме того, препараты для медицинских целей требуют специальных решений на стадии расфасовки и укупорки, так должны быть стерильными. Далее приводится характеристики каждой из стадий промышленного микробиологического синтеза.

Рис. 2. Система биотехнологического производства
Технология приготовления питательных сред для биосинтеза Основу питательных сред для культивирования микроорганизмов составляют источники углерода. Кроме углерода клетки микроорганизмов в процессе роста испытывают потребность в азоте, фосфоре, макро- и микроэлементах. Все вещества этого рода находятся в питательных средах в виде солей, исключение составляют среды, где азот и фосфор могут усваиваться растущими культурами из органических источников, например автолизатов или гидролизатов микробного или животного происхождения. Отделения приготовления питательной среды представляет собой цех, оборудованный емкостями для хранения жидких и твердых веществ, средствами их транспортировки и аппаратами с перемешивающими устройствами для приготовления растворов, суспензий или эмульсий. При этом питательные соли хранятся обычно в твердом виде, а приготовление их смеси с заданным соотношением компонентов производится в аппарате с мешалкой, куда подаются твердые компоненты в необходимом количестве и далее происходит их растворение. Иногда соединяются и перемешиваются заранее приготовленные растворы. Жидкие и твердые источники углерода обычно вводят в уже готовую питательную среду непосредственно перед ферментацией, так как это устраняет опасность заражения посторонней микрофлорой, вероятность которого возрастает при хранении готовой питательной смеси. При непрерывном культивировании в производстве микробного белка углеводороды и растворы солей вводят в ферментер раздельно по индивидуальным линиям, а смешение и эмульгирование нерастворимых в воде n-алканов происходит уже в самом биореакторе. При культивировании бактерий на метане последний постоянно барботируют в аппарат через специальные устройства. При периодической ферментации в начале процесса инокулят (засевная доза микроорганизмов) вносится в уже готовую питательную среду, содержащую все компоненты. Поэтому источники углерода вводят непосредственно перед засевом или отдельные компоненты среды вводят по мере потребления их культурой, поддерживая в ферментере некоторую оптимальную их концентрацию, которая на разных этапах ферментации может меняться по определенному закону. Важнейшим элементом приготовления питательных сред является соблюдение требований асептики. Это либо создание заданного значения pH, обеспечивающего подавление посторонних микроорганизмов, либо полная стерилизация всех подаваемых потоков и самого биореактора. Для стерилизации газовых потоков (в первую очередь воздуха) используют процесс фильтрации через специальные волокнистые фильтры с последовательно расположенными фильтрующими элементами. Фильтрующий материал периодически стерилизуется подачей острого пара в отключенный фильтр через заданные промежутки времени. Жидкостные потоки стерилизуют различными методами, из которых практический интерес представляют термический, радиационный, фильтрационный и отчасти химический. Термический — самый распространенный, при температурах порядка 120–150 °C. Радиационный — гамма-излучение, применяется редко из-за трудностей создания и эксплуатации мощных источников этого излучения. В отдельных случаях применяют химические стерилизующие агенты (вещества с ярко выраженным асептическим действием). Основная проблема в этом случае — необходимость устранения стерилизующего агента из питательной среды после гибели микрофлоры до внесения инокулята. Химические антисептики должны быть не только высокоэффективны, но и легко разлагаемы при изменении условий после завершения стерилизации. К числу лучших относится пропиолактон, обладающий сильным бактерицидным действием и легко гидролизуемый в молочную кислоту. Мало распространен и метод фильтрации, что объясняется аппаратными трудностями. Метод основан на способности полупроницаемых мембран с крупными порами пропускать жидкую фазу и концентрировать клетки микроорганизмов. В принципе этот метод является идеальным для стерилизации термически неустойчивых жидких и газовых средств, поскольку может осуществляться при низкой температуре и требует лишь градиента давления по разные стороны мембраны. Основная трудность — наличие термостойких мембран, способных выдерживать многократную стерилизацию их самих. В настоящее время эта проблема решается путем применения термостойких полимеров в производстве мембран. В заключение заметим, что ряд субстратов не требует стерилизации, так как они сами обладают асептическим действием; сюда относят метанол, этанол, концентрированная уксусная кислота и др. В этом случае ограничиваются стерилизацией прочих элементов питательной среды.
Поддержание чистой культуры и получение засевной дозы В технологическом процессе используются полезные свойства штамма, следовательно, необходимо сохранять и, если возможно, улучшать его производственные качества. Поэтому в биотехнологическом производстве имеется отделение чистой культуры, задачей которого является постоянное и надежное воспроизведение полезных свойств продуцента, найденных или достигнутых в свое время в ходе лабораторных исследований. Такое отделение проводит лабораторные операции по контролю и сохранению чистой культуры, а также маломасштабное культивирование для постоянной передачи штамма на стадию ферментации. Фактически это микробиологическая лаборатория, с музеем штаммов-продуцентов. В ходе контрольных высевов и маломасштабных ферментаций (в пробирках, колбах и т. д.) контролируется устойчивость всех имевшихся или приобретенных признаков, послуживших основанием для рекомендации к промышленному применению этих культур. По мере необходимости из отделения чистой культуры поступает заданная масса инокулята, идущая в производство. При периодическом процессе культивирования (при производстве метаболитов) в отделении чистой культуры готовят засевную дозу клеток для каждой из операций основного производства. При непрерывном производстве кормового белка этого не требуется, однако для повышения качества продукта предпочитают время от времени вводить клетки штамма-продуцента из отделения чистой культуры. Для этого в отделении имеется ферментационная часть, где производится выращивание достаточно крупных партий микроорганизма продуцента. Посевные дозы выращиваются последовательно в колбах и бутылях на 10–20 литров, находящихся на качалках или просто в термостатируемом помещении, и далее в последовательности ферментеров объемом (по необходимости) 10, 100, 500 и 1000 литров, в которых осуществляется перемешивание, аэрация и термостатирование культуральной жидкости с клетками. Отделение чистой культуры должно иметь достаточно большую коллекцию штаммов продуцентов, так как возможны временные переходы с одного штамма на другой, вызванные различными причинами. Например, сезонные изменения температуры частично компенсируются подбором достаточно продуктивных термотолерантных штаммов. Кроме того, микробиологическая промышленность зачастую вынуждена использовать в качестве компонентов питательных сред отходы сельского хозяйства и пищевой промышленности (меласса, кукурузный экстракт), что ведет к сезонным изменениям сырья и предполагает адаптацию продуцента к особенностям среды. Все это делает роль микробиологической службы производства достаточно высокой.
Ферментация Стадия ферментации — центральная среди этапов промышленного производства. Под ферментацией понимают всю совокупность последовательных операций от внесения в заранее приготовленную и термостатированную среду инокулята до завершения процессов роста, биосинтеза или биотрансформации. Ферментация проходит в специальных емкостях, называемых ферментерами или биореакторами. Конструкция биореактора приведена на рис. 3. Основными элементами ферментера являются двойные стенки, промежуток между которыми заполняется охлаждающей или нагревающей жидкостью, входные отверстия для газовых и жидких потоков, система контроля за составом питательной среды и условиями внутри реактора. Поскольку в промышленной биотехнологии выделяют 2 типа процессов — накопление биомассы и накопление ценных веществ, возникающих в ходе роста и последующего развития культуры, то меняется и характер построения производства во времени. Биомасса одноклеточных выращивается непрерывным способом в аппаратах хемостатного типа, а все процессы второй группы осуществляются периодически, когда в одном и том же аппарате в производственном цикле протекают все необходимые фазы развития клеток и биосинтеза. Процессы двух рассматриваемых типов отличаются по требованиям к степени асептики, что связано с их объёмами — белок одноклеточных выпускается миллионами тонн сухого вещества, а выпуск продуктов второго типа составляет, как максимум, тысячи или десятки тысяч тонн. Поэтому в производстве белковых веществ ограничиваются достаточно высокой, но не 100 % степенью асептики, обеспечивая последнюю подбором режима культивирования, подходящего для продуцента, но неблагоприятного для возможных примесных штаммов. Технологическое оформление процессов промышленной биотехнологии в значительной мере определяется отношением микроорганизма-продуцента к кислороду. При использовании аэробных культур ферментационное оборудование и нормы технологического режима подбираются таким образом, чтобы массообмен (перенос кислорода из газовой в жидкую фазу) обеспечивал поступление кислорода к клеткам в количествах, необходимых и оптимальных для данной культуры в данной фазе роста. Промышленное использование факультативных анаэробов не ставит задачи абсолютного исключения кислорода из среды, поэтому процессы этого типа (брожение) технологически проще аэробных. В начальной фазе этих процессов требуется лишь удалить кислород из газовой фазы над культуральной жидкостью, что может быть достигнуто введением инертного газа или просто вытеснением воздуха углекислотой, выделяемой клетками при метаболизме. Технологическое оформление строго анаэробных процессов сложнее, чем для процессов брожения, так как в этом случае необходимо полностью исключить возможность попадания кислорода в газовую, а оттуда и в жидкую среду.
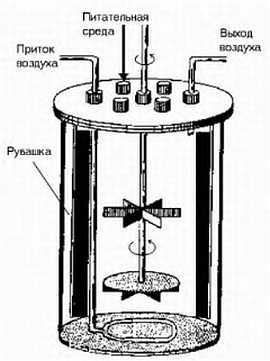
Рис. 3. Устройство ферментера
Вопросы термостатирования ферментационного процесса (подвода или отвода тепла в ходе ферментации) являются очень острыми в целом ряде производств биотехнологии. В аэробных условиях микробиологический синтез протекает со значительным тепловыделением, что вызывает необходимость отвода тепла из аппаратов большого объема (сотни и тысячи кубометров). Технологические требования к скорости теплоотвода очень жесткие из-за узкого температурного оптимума роста культуры (2–3 °C). Наиболее приемлемый на практике способ теплоотвода — охлаждение водой через змеевики, рубашки и др. устройства — осложняется небольшой разностью температур между содержимым биореактора (32–34 °C для дрожжей Candida) и охлаждающей водой (20 °C), температура которой в жаркое время года еще выше. Поэтому в реакторе создается развитая поверхность газообмена, увеличивается скорость движения жидкостей и т. д. Важно также поддерживать определенный состав питательной среды. В непрерывных процессах биосинтеза задача технолога сводится к поддержанию концентрации всех питательных веществ (и кислорода) и дозированному введению кислоты или щелочи для рН-статирования системы на заданном уровне. Простейшим вариантом управления стадией ферментации в периодическом режиме является изменение концентраций компонентов среды и её pH, а также введение необходимых добавок по заранее разработанной программе, реализуемой технологом в каждом цикле ферментации. Этот способ относительно прост и легко поддается автоматизации. Во многих случаях необходимо возможно полно исчерпывать компоненты питательной среды, чтобы они не попадали на последующие стадии переработки. Эта необходимость может быть вызвана рядом причин: — дороговизна или дефицитность субстрата; — вредное воздействие субстрата на качество готового продукта (например, при производстве дрожжей на парафинах, когда выделение остаточных количеств углеводородов из клеточной массы затруднено, поэтому добавляют дополнительные секции для дозревания или утилизации запасенных в цитоплазме углеводородов); — затруднения, возникающие на стадии выделения и очистки метаболитов при одновременном присутствии в культуральной жидкости неутипизированных веществ.
Общие принципы разделения веществ Продукты микробного синтеза поступают из биореактора в виде водных суспензий или растворов, при этом характерно невысокое содержание основного компонента и наличие многих примесных веществ. В большинстве промышленных производств на первом этапе переработки культуральной жидкости производят отделение массы продуцента от жидкой фазы — сепарацию. Жидкость далее также подвергается переработке, если содержит метаболиты, представляющие практическую ценность. В производствах, где целевым продуктом являются клетки как источник белка, культуральная жидкость подвергается лишь очистке, позволяющей использовать водную фазу многократно и снизить образование сточных вод. Технологические приемы, используемые для отделения клеток от среды зависят от природы продуцента. Например, сахаромицеты (хлебопекарные дрожжи) имеют относительно большие клетки и способны флотироваться, поэтому после сгущения биомассы флотацией их отделяют на обычных барабанных вакуум-фильтрах. В дальнейшем биомассу, снятую с фильтра, подвергают прессованию и получают продукт с высоким содержанием живых клеток, имеющих высокую хлебопекарную активность. Дрожжи же рода Candida, служащие источником кормового белка плохо флотируются и фильтруются. Поэтому дрожжи, растущие на углеводородах, а также бактерии-продуценты белка на основе метана и метанола, на первом этапе сепарируются, причем в несколько ступеней. Оставшаяся вода удаляется путем выпаривания, а все компоненты жидкой фазы остаются в конечном продукте. К аналогичному приему прибегают и при производстве бактериальных энтомопатогенных препаратов и удобрений. Конечный продукт удается получить в активной форме лишь в принципе отказавшись от выделения его из культуральной жидкости: содержимое реактора выпаривают и сушат в условиях, обеспечивающих жизнеспособность конечного продукта. Неутилизированные компоненты культуральной жидкости могут отразиться на способности продукта к хранению. При выделении и очистке метаболитов биомасса, если она не содержит заметных количеств целевого продукта, осаждается добавлением извести или других твердых компонентов, увлекающих клетки или мицелий на дно — физическое осаждение. Отделение твердой фазы (мелкодисперсный клеточный материал, внутриклеточные биополимеры возможно и методом фильтрации. Так как фильтруемая суспензия склонная к гелеобразованию, то производительность фильтров быстро падает. Предотвратить это можно добавлением в смесь или на фильтрующую ткань размолотых вулканических пород, содержащих оксиды кремния и алюминия, тогда осадки приобретают пористую структуру. Некоторые виды биомассы отделяют центрифугированием. Осаждение взвешенных частиц происходит под действием центробежной силы. После разделения образуется 2 фракции: биомасса (твердая) и культуральная жидкость. Культуральная жидкость перерабатывается путем экстракции, ионообмена, кристаллизации или с помощью микро- и ультрафильтрации через полимерные мембраны со специально подобранным размером пор. Для выделения и очистки продуктов, находящихся внутри клеток продуцента (например интерферонов, гормонов) вводится стадия разрушения клеточных оболочек (дезинтеграция биомассы); обычно для этого применяются механические, химические или комбинированные методы. К физическим методам дезинтеграции относятся обработка ультразвуком, вращение лопасти или вибратора, встряхивание со стеклянными бусами, продавливание через узкое отверстие под давлением, раздавливание замороженной клеточной массы, растирание в ступке, осмотический шок, замораживание-оттаивание, декомпрессия (сжатие с последующим резким снижением давления). Химические и химико-ферментативные методы более избирательны. Клетки могут быть разрушены толуолом или бутанолом, антибиотиками, ферментами. Культуральную жидкость освобождают от сопутствующих растворимых веществ и фракционируют. Освобождение от растворимых веществ производят несколькими способами: 1. Осаждение — физическое (нагревание, охлаждение, разбавление, концентрирование) или химическое (с помощью органических и неорганических веществ). Осаждение органическими растворителями основано на снижении диэлектрической постоянной среды[55]. Устойчивость белковых растворов обусловлена наличием гидратного слоя у молекулы. Если его разрушить, белки осаждаются. Для этого молекулы добавляемых веществ должны быть более гидрофильны, чем молекулы белков. В качестве осадителей используют этанол, метанол, ацетон, изопропанол. При разных количествах растворителя и разных значения pH осаждаются разные фракции. Пример: 50 % этанол осаждает 80 % протеазы и 3–5 % амилазы, 70 % спирт осаждает 98 % амилазы. Высаливание — механизм тот же, что и при действии органических веществ, гидратируются диссоциирующие ионы неорганических солей. Как наиболее дешёвый реагент используют сульфат аммония. Также применяют сульфаты натрия, магния и фосфат калия. 2. Экстракция. При твердожидкофазной экстракции вещество из твердой фазы переходит в жидкую, при жидкожидкофазной — из одной жидкости в другую (например, хлорофилл из спиртовой вытяжки переходит в бензин). Для извлечения антибиотиков, витаминов, каротиноидов, липидов применяют жидкожидкофазную экстракцию, когда культуральную жидкость смешивают с органическими растворителями. 3. Адсорбция — частный случай экстракции, когда экстрагирующий агент — твердое тело. Адсорбция применяется для веществ, имеющих функциональные группы, заряженные положительно или отрицательно. В качестве адсорбента используют иониты на основе целлюлозы: — катионит — карбоксиметилцеллюлоза (КМЦ); — анионит — диэтиламиноэтилцеллюлоза (ДЭАЭ), а также сефадексы на основе декстрана и т. д. Адсорбция идет по ионообменному механизму.
Методы тонкой очистки и разделения препаратов Более тонкую очистку веществ осуществляют несколькими способами. Наибольшее распространение получила хроматография. Каплю образца[56] наносят на специальную бумагу (хроматография на бумаге) или пластинку стекла или пластмассы, покрытую тонким слоем инертного сорбента, например, целлюлозы или силикагеля (хроматография в тонком слое или тонкослойная хроматография). Затем такую пластинку одним концом помещают в смесь растворителей (например, воды и спирта). По мере движения растворителей по пластинке, они подхватывают те молекулы образца, которые растворяются в них. Растворители выбирают таким образом, чтобы они связывались сорбентом по-разному. В результате молекулы образца, более растворимые в связанном растворителе, движутся медленнее, а другие, более растворимые в слабо сорбированном растворителе, движутся быстрее. Через несколько часов пластинку сушат, окрашивают и определяют положение различных молекул. Можно разделять молекулы методом хроматографии на колонках (колоночная хроматография). В этом случае смесь молекул в растворе пропускают через колонку, содержащую твердый пористый матрикс. В результате взаимодействия с матриксом различные молекулы проходят через колонку с различной скоростью. После того как они достигнут в определенной последовательности дна колонки, их собирают отдельными фракциями. В настоящее время разработано и применяется множество матриксов различных типов, используя которые можно делить белки согласно их: — заряду (ионообменная хроматография), — гидрофобности (гидрофобная хроматография), — размеру (хроматография гель-фильтрацией) или способности связываться различными химическими группами (аффинная хроматография). При ионообменной хроматографии нерастворимый матрикс содержит ионы, задерживающие молекулы с противоположным зарядом. Для разделения молекул используются следующие матриксы: диэтиламиноэтилцеллюлоза (ДЭАЭ-целлюлоза) — заряжена положительно; карбоксиметилцеллюлоза (КМ-целлюлоза) и фосфоцеллюлоза — заряжены отрицательно. Силы взаимодействия между молекулами в растворе и ионообменником определяются ионной силой и pH элюирующего раствора. Гидрофобные колонки наполнены шариками, из которых выступают гидрофобные цепи; в таких колонках задерживаются белки с обнаженными гидрофобными участками. Колонки, предназначенные для гель-фильтрации, заполнены крошечными пористыми инертными шариками; при использовании таких колонок происходит разделение белков по размерам. Молекулы небольшого размера по мере прохождения через колонку проникают внутрь шариков, а более крупные молекулы остаются в промежутках между шариками. В результате они быстрее проходят через колонку и выходят из нее первыми. В качестве матрикса можно использовать зерна поперечно-сшитого полисахарида (декстран или агароза). Гель-фильтрация обычно используется и для разделения молекул, и для определения их размеров. Гораздо более эффективен метод аффинной хроматографии (хроматография по сродству). В основе этого метода лежат биологически важные взаимодействия, происходящие на поверхности белковых молекул. При аффинной хроматографии используется нерастворимый матрикс, ковалентно связанный со специфичными лигандами (антителами или субстратом ферментов), которые присоединяют определенный белок. Связываемые иммобилизованным субстратом молекулы фермента можно элюировать концентрированными растворами субстрата в свободной форме, а молекулы, связанные с иммобилизованными антителами, можно элюировать за счет диссоциации комплекса антитело-антиген концентрированными растворами соли или растворами низкого или высокого pH. Однократная хроматография на такой колонке позволяет зачастую достигнуть очень высокой степени очистки препарата. Разрешение обычной колоночной хроматографии ограничено негомогенностью матриксов (например, целлюлозы), что вызывает неравномерное протекание растворителя через колонку. Разработанные недавно хроматографические смолы (в основу которых обычно положен кремний) имеют форму мельчайших сфер от 3 до 10 мкм в диаметре, которые упакованы в специальный чехол и образуют гомогенную колонку. Такие колонки для высокоэффективной жидкостной хроматографии (ВЖХ) обеспечивают высокий уровень разрешения. Поскольку частицы носителя в колонках для ВЖХ упакованы очень плотно, в отсутствие высокого давления скорость потока через них незначительна. По этой причине такие колонки обычно помещают в стальные цилиндры, соединенные со сложной системой насосов и шлангов, которые обеспечивают необходимое для высокой скорости протока давление. В традиционной колоночной хроматографии скорость протекания через колонку может быть довольно низкой (примерно один объем колонки в час), таким образом, у разделяемых растворов достаточно времени для уравновешивания с внутренним содержимым крупных частиц матрикса. В условиях ВЖХ происходит быстрое уравновешивание растворов с внутренним содержимым крошечных сфер, так что растворы, обладающие различным сродством к матриксу, эффективно разделяются даже при высокой скорости потока. Таким образом, ранее для достижения плохого разделения с помощью колоночной хроматографии требовались часы, а в настоящее время благодаря ВЖХ качественное фракционирование занимает минуты. Вот почему именно этот метод чрезвычайно популярен сейчас для разделения и белков, и малых молекул. Экстракты разрушенных клеток можно фракционировать, подвергая их высокоскоростному центрифугированию. Такая обработка делит клеточные компоненты по их размеру: более крупные частицы при центрифугировании движутся быстрее. Крупные компоненты экстракта, в том числе ядра или неразрушенные клетки, быстро оседают (седиментируют) при относительно низких скоростях и образуют осадок на дне центрифужной пробирки. Центрифугирование является, как правило, первым этапом фракционирования, с его помощью разделяются только значительно отличающиеся по размеру компоненты. Чтобы достигнуть более высокой степени разделения фракций, необходимо гомогенат наслоить тонким слоем поверх солевого раствора. При ультрацентрифугировании различные фракции седиментируют с различной скоростью и образуют отдельные полосы, которые можно выделить. Во избежание перемешивания осажденных компонентов солевой раствор должен содержать инертный и хорошо растворимый материал (например, сахарозу), плотность которого постепенно увеличивается сверху вниз, формируя градиент плотности. При седиментации сквозь такие градиенты сахарозы различные компоненты клетки собираются в отдельные полосы, которые можно выделить. Скорость седиментации каждого из компонентов определяется его размерами и формой и обычно выражается с помощью коэффициента седиментации, обозначаемого S[57]. Скорость вращения до 80000 об/мин, так что на разделяемые частицы действуют силы, превосходящие силу тяготения более чем в 500000 раз. Под действием столь больших сил даже сравнительно небольшие макромолекулы, такие, как тРНК или простейшие ферменты, разделяются и распределяются в строгом соответствии со своими размерами. Измерение коэффициента седиментации макромолекулярных комплексов обычно используют для определения их общей массы и количества, входящих в их состав субъединиц. Ультрацентрифуга разделяет клеточные компоненты не только по массе, но и по плавучей плотности. В этом случае образец седиментирует в крутом градиенте, образованном высококонцентрированным раствором сахарозы или хлористого цезия. Компоненты клеток опускаются по градиенту до тех пор, пока не достигнут участка, плотность раствора, в котором равна собственной плотности компонентов. Дальнейшей седиментации компонентов не происходит и они «застревают» на этом уровне. Таким образом, в центрифужной пробирке возникает набор различных полос. Метод центрифугирования в градиенте хлористого цезия был разработан в 1957 году для доказательства полуконсервативности репликации ДНК. Электрофорез — метод разделения белков и нуклеиновых кислот в свободном водном растворе и пористом матриксе, в качестве которого можно использовать полисахариды, например, крахмал или агарозу. Биомолекулы обычно несут суммарный положительный или отрицательный заряд, обусловленный наличием на их поверхности положительно или отрицательно заряженных групп аминокислот. Если белковые молекулы поместить в электрическое поле, они начинают перемещаться со скоростью, которая определяется их суммарным зарядом, а также формой и размерами. В середине 60-х годов был разработан модифицированный метод электрофореза — электрофорез в полиакриламидном геле в присутствии додецил-сульфата натрия (ДСН-ПААГ). При использовании данного метода белки мигрируют в инертном матриксе-полиакриламидном геле с высоким содержанием поперечных сшивок. Обычно гель готовят полимеризацией мономеров непосредственно перед использованием. Размеры пор геля могут быть подобраны произвольно с тем, чтобы гель мог замедлить миграцию определенных молекул. При этом белки находятся в растворе, содержащем мощный, отрицательно заряженный детергент — додецил-сульфат натрия или ДСН (SDS). Связываясь с гидрофобными участками белковой молекулы, этот детергент вызывает развертывание белковых молекул в длинные вытянутые цепи. Каждая молекула белка связывает значительное количество молекул детергента, приобретая суммарный отрицательный заряд. По этой причине белок после того, как будет приложено напряжение, начнет двигаться в направлении положительного электрода. Белки одного размера ведут себя сходным образом, поскольку, во-первых, их природная структура полностью нарушена ДСН так, что их форма идентична, во-вторых, они связывают одинаковое количество ДСН и приобретают одинаковый негативный заряд. Крупные белки, обладающие большим зарядом, подвергаются действию значительных электрических сил, а также более существенному торможению. В обычных растворах эти эффекты, как правило, взаимно погашаются, но в порах полиакриламидного геля, действующего как молекулярное сито, большие молекулы тормозятся значительно сильнее, чем малые, поэтому оказываются ближе к стартовой линии. Смесь молекул делится на ряд полос, расположенных в соответствии с их молекулярной массой. Выявить эти полосы можно путем окрашивания соответствующим красителем[58]. Например, белки индентифицируются красителем кумасси синим. Известно, что близко расположенные полосы в геле могут перекрываться. Этот эффект препятствует выявлению большого количества белков (не больше 50) с помощью одномерных методов их разделения. Метод двумерного гель-электрофореза, в котором объединены две различные процедуры разделения, позволяет идентифицировать более 1000 белков. Результаты при этом получают в виде «двумерной» белковой карты. При работе данным методом на первом этапе белки разделяют по их заряду. Для этого образец помещают в небольшой объем раствора, содержащего неионный (незаряженный) детергент-меркаптоэтанол, и в качестве денатурирующего агента-мочевину. В этом растворе происходит солюбилизация, денатурация и диссоциация всех без исключения полипептидных цепей; при этом изменения заряда цепей не происходит. Диссоциированные полипептидные цепи разделяют затем методом изоэлектрического фокусирования, основанном на изменении заряда белковой молекулы при изменении pH окружающей среды. Каждый из белков может быть охарактеризован изоэлектрической точкой — значением pH, при котором суммарный заряд белковой молекулы равен нулю, и, следовательно, белок не способен перемещаться под действием электрического поля. При изоэлектрическом фокусировании белки подвергаются электрофорезу в геле, в котором с помощью специальных буферов создается градиент pH. Под действием электрического поля каждый белок перемещается в ту зону градиента, которая соответствует его изоэлектрической точке и остается в ней. Так происходит разделение белков в одном направлении двумерного гельэлектрофореза. На втором этапе гель, содержащий разделенные белки, снова подвергается электрофорезу, на этот раз в направлении перпендикулярном тому, что на первом этапе. В этом случае электрофорез ведут в присутствии ДСН и белки разделяют по их молекулярной массе, как в одномерном ДСН-ПААГ. Исходный гель пропитывают додецил-сульфатом натрия и, поместив его на блок ДСН-ПААГ-геля, проводят электрофорез, в ходе которого каждая из полипептидных цепей мигрирует сквозь блок геля и образует в нем отдельную полосу. Неразделенными в результате остаются только те бели, которые неразличимы как по изоэлектрической точке, так и по молекулярной массе; такое сочетание встречается очень редко.
Получение товарных форм препаратов Все товарные формы биопрепаратов с точки зрения технологии их получения можно разделить на три основные группы. 1. Биопрепараты, имеющие в товарном продукте в качестве основного компонента жизнеспособные микроорганизмы. К этой группе относятся средства защиты растений, бактериальные удобрения, закваски для силосования кормов, биодеграданты, другие активные средства биотрансформации. 2. Биопрепараты, в состав которых входит инактивированная биомасса клеток и продукты ее переработки. Это кормовые дрожжи, грибной мицелий и т. д. 3. Биопрепараты на основе очищенных продуктов метаболизма микроорганизмов. К ним относятся витамины, аминокислоты, ферменты, антибиотики, биолипиды, полисахариды, продукты комплексной переработки микробных масс и метаболитов. В зависимости от принятых на предыдущей стадии решения товарные формы представляют собой либо сложную смесь, содержащую некоторое количество основного вещества, либо высокоочищенный препарат, отвечающий ряду специальных требований. Продукт может выпускаться в жидком (например жидкий концентрат лизина) или сухом виде (белково-витаминный концентрат, энтомопатогенные препараты, кормовой концентрат лизина). Стадия фасовки рассмотренных комплексных препаратов заключается в помещении их в тару (мешки, барабаны и т. п.), размеры и тип которой определяются потребностями заказчика и свойствами продукта (его слеживаемостью, гигроскопичностью, стойкостью к загниванию и т. д.). Другие требования предъявляются к медицинским препаратам и биохимическим реактивам.
ПРОИЗВОДСТВО БЕЛКА МИКРООРГАНИЗМОВ
Продуценты белка Производство микробной биомассы — самое крупное микробиологическое производство. Микробная биомасса может быть хорошей белковой добавкой для домашних животных, птиц и рыб. Производство микробной биомассы особенно важно для стран, не культивирующих в больших масштабах сою (соевую муку используют как традиционную белковую добавку к кормам). При выборе микроорганизма учитывают удельную скорость роста и выход биомассы на данном субстрате, стабильность при поточном культивировании, величину клеток. Клетки дрожжей крупнее, чем бактерий, и легче отделяются от жидкости при центрифугировании. Можно выращивать полиплоидные мутанты дрожжей с крупными клетками. В настоящее время известны только две группы микроорганизмов, которым присущи свойства, необходимые для крупномасштабного промышленного производства: это дрожжи рода Candida на n-алканах (нормальных углеводородах) и бактерии Methylophillus methylоtrophus на метаноле. Микроорганизмы можно выращивать и на других питательных средах: на газах, нефти, отходах угольной, химической, пищевой, винно-водочной, деревообрабатывающей промышленности. Экономические преимущества их использования очевидны. Так, килограмм переработанной микроорганизмами нефти дает килограмм белка, а, скажем, килограмм сахара[59] — всего 500 граммов белка. Аминокислотный состав белка дрожжей практически не отличается от такового, полученного из микроорганизмов, выращенных на обычных углеводных средах. Биологические испытания препаратов из дрожжей, выращенных на углеводородах, которые проведены и у нас в стране и за рубежом, выявили полное отсутствие у них какого-либо вредного влияния на организм испытуемых животных. Опыты были проведены на многих поколениях десятков тысяч лабораторных и сельскохозяйственных животных. В непереработанном виде дрожжи содержат неспецифические липиды и аминокислоты, биогенные амины, полисахариды и нуклеиновые кислоты, а их влияние на организм пока еще плохо изучено. Поэтому и предлагается выделять из дрожжей белок в химически чистом виде. Освобождение его от нуклеиновых кислот также уже стало несложным. В современных биотехнологических процессах, основанных на использовании микроорганизмов, продуцентами белка служат дрожжи, другие грибы, бактерии и микроскопические водоросли. С технологической точки зрения наилучшими из них являются дрожжи. Их преимущество заключается прежде всего в "технологичности": дрожжи легко выращивать в условиях производства. Они характеризуются высокой скоростью роста, устойчивостью к посторонней микрофлоре, способны усваивать любые источники питания, легко отделяются, не загрязняют воздух спорами. Клетки дрожжей содержат до 25 % сухих веществ. Наиболее ценный компонент дрожжевой биомассы — белок, который по составу аминокислот превосходит белок зерна злаковых культур и лишь немного уступает белкам молока и рыбной муки. Биологическая ценность дрожжевого белка определяется наличием значительного количества незаменимых аминокислот. По содержанию витаминов дрожжи превосходят все белковые корма, в том числе и рыбную муку. Кроме того, дрожжевые клетки содержат микроэлементы и значительное количество жира, в котором преобладают ненасыщенные жирные кислоты. При скармливании кормовых дрожжей коровам повышаются удои и содержание жира в молоке, а у пушных зверей улучшается качество меха. В последнее время в качестве продуцентов белка стали использовать бактерии, которые отличаются высокой скоростью роста и содержат в биомассе до 80 % белка. Бактерии хорошо поддаются селекции, что позволяет получать высокопродуктивные штаммы. Их недостатками являются трудная осаждаемость, обусловленная малыми размерами клеток, значительная чувствительность к фаговым инфекциям и высокое содержание в биомассе нуклеиновых кислот. Последнее обстоятельство неблагоприятно только в том случае, если предусматривается пищевое использование продукта. Снижать содержание нуклеиновых кислот в биомассе, употребляемой на корм животным, нет необходимости, так как мочевая кислота и ее соли, образующиеся при разрушении азотистых оснований, превращаются в организме животных в алантоин, который легко выделяется с мочой. У человека избыток солей мочевой кислоты может способствовать развитию ряда заболеваний. Следующую группу продуцентов белка составляют грибы. Они привлекают внимание исследователей благодаря способности утилизировать самое разнообразное по составу органическое сырье: мелассу, молочную сыворотку, сок растений и корнеплодов, лигнин- и целлюлозосодержащие твердые отходы пищевой, деревообрабатывающей, гидролизной промышленности. Грибной мицелий богат белковыми веществами, которые по содержанию незаменимых аминокислот ближе всего к белкам сои. Вместе с тем белок грибов богат лизином, основной аминокислотой, недостающей в белке зерновых культур. Это позволяет на основе зерна и грибной биомассы составлять сбалансированные пищевые и кормовые смеси. Грибные белки имеют достаточно высокую биологическую ценность и хорошо усваиваются организмом. Положительным фактором является и волокнистое строение выращенной культуры. Это позволяет имитировать текстуру мяса, а с помощью различных добавок — его цвет и запах. Хранят грибной мицелий обычно в замороженном виде. В качестве субстрата грибами используются глюкоза и другие питательные вещества, а общим источником азота служат аммиак и аммонийные соли. После завершения стадии ферментации культуру подвергают термообработке для уменьшения содержания рибонуклеиновой кислоты, а затем отделяют мицелий методом вакуумного фильтрования. Источниками белковых веществ могут служить и водоросли. При фототрофном способе питания и образования биомассы они используют углекислый газ атмосферы. Выращивают водоросли, как правило, в поверхностном слое прудов, где с площади 0,1 га можно получить столько же белка, сколько с 14 га посевов фасоли. Белок водорослей пригоден не только для кормовых, но и пищевых целей. Наконец, хорошими продуцентами белка являются рясковые, которые накапливают протеина до 45 % от сухой массы, а также до 45 % углеводов. Однако, несмотря на свои малые размеры, они не принадлежат к вышеперечисленным производителям белка (микроорганизмам), так как не только являются многоклеточными организмами, но и относятся к высшим растениям.
Субстраты для получения белка В качестве источников вещества и энергии микроорганизмы используют самые разнообразные субстраты — нормальные парафины и дистилляты нефти, природный газ, спирты, растительные гидролизаты и отходы промышленных предприятий. Для выращивания микроорганизмов с целью получения белка хорошо бы иметь богатый углеродом, но дешевый субстрат. Этому требованию вполне отвечают нормальные (неразветвленные) парафины нефти. Выход биомассы может достигать при их использовании до 100 % от массы субстрата. Качество продукта зависит от степени чистотыпарафинов. Дрожжи, выращенные на недостаточно очищенных парафинах, содержат неметаболизированные компоненты. При использовании парафинов достаточной степени очистки, полученная дрожжевая масса может успешно применяться в качестве дополнительного источника белка в рационах животных. В нашей стране мало районов, пригодных для выращивания сои, являющейся основным источником белковых добавок. Поэтому наложено крупнотоннажное производство кормовых дрожжей на n-парафинах. Действует несколько заводов мощностью от 70 до 240 тыс. тонн в год[60]. Сырьем служат жидкие очищенные парафины. Одним из перспективных источников углерода для культивирования продуцентов белка высокого качества считается метиловый спирт. Его можно получать методом микробного синтеза на таких субстратах, как древесина, солома, городские отходы. Использование метанола в качестве субстрата затруднено из-за его химической структуры: молекула метанола содержит один атом углерода, тогда как синтез большинства органических соединений осуществляется через двухуглеродные молекулы. Наилучшими продуцентами на этом субстрате считаются бактерии, потому что они могут расти на метаноле с добавлением минеральных солей. Процессы получения белка на метаноле достаточно экономичны. По данным концерна Ай-Си-Ай (Великобритания), себестоимость продукта, производимого на метаноле, на 10–15 % ниже, чем при аналогичном производстве, базирующемся на основе высокоочищенных n-парафинов. Высокобелковые продукты из метанола получают фирмы ряда развитых стран мира: Великобритании, Швеции, Германии, США, Италии. Продуцентами белка служат бактерии рода Methylomonas. Например, западногерманская фирма Хёхст производит из метанола бактериальную биомассу на установке производительностью 1000 тонн в год. В продукте содержится 60 % белка. Цель фирмы — получение пищевого белка. Использование этанола как субстрата снимает проблему очистки биомассы от аномальных продуктов обмена с нечетным числом углеродных атомов. Стоимость такого производства несколько выше. Биомассу на основе этанола производят в Чехословакии, Испании, Германии, Японии, США. В США, Японии, Канаде, ФРГ, Великобритании разработаны технологические процессы получения белка на природном газе. Выход биомассы в этом случае может составлять 66 % от массы субстрата. В разработанном в Великобритании процессе используется смешанная культура: бактерии Methylomonas, усваивающие метан, Hypomicrobium и Pseudomonas, усваивающие метанол, и два вида неметилотрофных бактерий. Культура характеризуется высокой скоростью роста и продуктивностью. Главные достоинства метана (кстати сказать, основного компонента природного газа) — доступность, относительно низкая стоимость, высокая эффективность преобразования в биомассу метаноокисляющими микроорганизмами, значительное содержание в биомассе белка, сбалансированного по аминокислотному составу. Бактерии, растущие на метане хорошо переносят кислую среду и высокие температуры, в связи с чем устойчивы к инфекциям. Субстратом для микробного синтеза может быть и минеральный углерод — углекислый газ. Окисленный углерод в данном случае с успехом восстанавливается микроводорослями при помощи солнечной энергии и водородоокисляющими бактериями при помощи водорода. На корм скоту используют суспензию водорослей. Для работы установок по выращиванию водорослей необходимы стабильные климатические условия — постоянные температуры воздуха и интенсивность солнечного света. Наиболее перспективно получение белка с помощью водородоокисляющих бактерий, которые развиваются за счет окисления водорода кислородом воздуха. Энергия, высвобождающаяся в этом процессе, идет на усвоение углекислого газа. Для получения биомассы используются, как правило, бактерии рода Hydrogenomonas. Первоначально интерес к ним возник при разработке замкнутых систем жизнеобеспечения, а затем их стали изучать с точки зрения использования в качестве продуцентов высококачественного белка. В институте микробиологии Геттингенского университета (Германия) разработан способ культивирования водородоокисляющих бактерий, при котором можно получать 20 г сухого вещества на 1 литр суспензии клеток. Возможно, в будущем эти бактерии станут основным источником пищевых микробных белков. Исключительно доступным и достаточно дешевым источником углеводов для производства микробного белка является растительная биомасса. Любое растение содержит разнообразные сахара. Целлюлоза — полисахарид, состоящий из молекул глюкозы. Гемицеллюлоза состоит из остатков арабинозы, галактозы, маннозы, фруктозы. Проблема в том, что полисахариды древесины связаны жесткими оксифенилпропановыми звеньями лигнина — полимера, почти не поддающегося разрушению. Поэтому гидролиз древесины происходит только в присутствии катализатора — минеральной кислоты и при высоких температурах. При этом образуются моносахара — гексозы и пентозы. На жидкой, содержащей сахара, фракции гидролизата выращивают дрожжи. При кислотном гидролизе древесины образуется ряд побочных продуктов (фурфурол, меланины), а из-за высоких температур может произойти карамелизация сахаров. Эти вещества препятствуют нормальному росту дрожжей, их отделяют от гидролизата и по возможности используют. В качестве продуцентов используют штаммы Candida scotti и С. tropicalis. Наиболее крупным производителем сырья для гидролизной промышленности являются деревообрабатывающие предприятия, отходы которых достигают ежегодно десятки миллионов тонн. К сожалению, нерационально или не используются вообще отходы производства лубяных волокон (из льна и конопли), картофелекрахмального производства, пивоваренной, плодоовощной, консервной промышленности, свекловичный жом. Особого внимания заслуживают способы прямой биоконверсии продуктов фотосинтеза и их производных в белок с помощью грибов. Эти организмы благодаря наличию мощных ферментных систем способны утилизировать сложные растительные субстраты без предварительной обработки. Исследования условий биоконверсии растительных субстратов в микробный белок активно ведутся в США, Канаде, Индии, Финляндии, Швеции, Великобритании, в нашей стране и других странах мира. Однако в литературе сведения о широкомасштабном производстве белков микробного происхождения немногочисленны. Наиболее известным и доведенным до стадии промышленной реализации является процесс "Ватерлоо", разработанный в университете Ватерлоо в Канаде. Это процесс, основанный на выращивании целлюлозоразрушающих грибов Chaetomium cellulolyticum, можно осуществлять как в глубинной культуре, так и поверхностным методом. Содержание белка в конечном продукте (высушенном грибном мицелии) составляет 45 %. Финская фирма "Тампелла" разработала технологию и организовала производство белкового кормового продукта "Пекило" на отходах целлюлозно-бумажного производства. Продукт содержит до 60 % протеина с хорошим аминокислотным профилем и значительное количество витаминов группы В. В большинстве стран — производителей молока традиционным способом утилизации сыворотки является скармливание её животным. Степень конверсии белка сыворотки в белок животного весьма невысока (для выработки 1 кг животного белка необходимо 1700 кг сыворотки). В последние 10–15 лет из сыворотки методом ультрафильтрации выделяют белки высокого качества, на основе которых делают заменители сухого обезжиренного молока и другие продукты. Концентраты можно использовать как пищевые добавки и компоненты детского питания. Из сыворотки производится и молочный сахар — лактоза, применяемая в пищевой и медицинской промышленности. При всем при этом объем промышленной переработки сыворотки составляет 50–60 % от её общего производства. Следовательно, налицо большие потери ценнейшего молочного белка и лактозы. Более того, возникает проблема утилизации отходов, так как процесс естественного разложения сыворотки происходит крайне медленно. Лактоза молочной сыворотки может служить источником энергии для многих видов микроорганизмов, сырьем для производства продуктов микробного синтеза (органических кислот, ферментов, спиртов, витаминов) и белковой биомассы. Из всех известных микроорганизмов самым высоким коэффициентом конверсии белка сыворотки в микробный белок обладают дрожжи. Впервые дрожжи на молочной сыворотке стали выращивать в Германии. В качестве продуцентов применяли различные штаммы сахаромицетов. Разработаны способы получения микробных продуктов, основанные на использовании лактозы как монокультурой, так и смесью дрожжей и бактерий. В настоящее время в качестве продуцентов используют дрожжи родов Candida, Trichosporon, Torulopsis. Молочная сыворотка с выросшими в ней дрожжами по биологической ценности значительно превосходит исходное сырье и её можно использовать в качестве заменителя молока. Приведенный перечень микроорганизмов и процессов получения белка одноклеточных не является исчерпывающим. Однако потенциал этой новой отрасли производства используется далеко не полностью. Кроме того, мы еще не знаем всех возможностей деятельности микроорганизмов в качестве продуцентов белка, но по мере углубления наших знаний, они будут расширены.
ТЕХНОЛОГИЯ ПОЛУЧЕНИЯ МИКРОБНЫХ ЛИПИДОВ
Под липидами подразумеваются все растворимые в неполярных растворителях клеточные компоненты микроорганизмов. В настоящее время ведутся поиски новых источников получения жиров, в том числе и на технические нужды. Этим источником могут стать микроорганизмы, липиды которых после соответствующей обработки пригодны для использования в различных отраслях промышленности: медицинской, химико-фармакоцевтической, лакокрасочной, шинной и других, что позволит высвободить значительные количества масел животного и растительного происхождения. Технологический процесс получения микробных липидов, в отличие от получения белковых веществ, обязательно включает стадию выделения липидов из клеточной массы методом экстракции в неполярном растворителе (бензине или эфире). При этом получают одновременно два готовых продукта: микробный жир (биожир) и обезжиренный белковый препарат (биошрот). Сырьем для этого процесса являются те же среды, что и для производства кормовой биомассы. В процессе культивирования микроорганизмов на различных средах получаются три класса липидов: простые, сложные липиды и их производные. Простые липиды — нейтральные жиры и воски. Нейтральные жиры (основные запасные компоненты клетки) — эфиры глицерина и жирных кислот, основная масса которых триацилглицериды (есть, впрочем ещё и моно- и диглицериды). Воски — эфиры жирных кислот или моноооксикислот и алифатических спиртов с длинной углеродной цепью. По структуре и свойствам близки к нейтральным липидам. Наибольшее количество нейтральных липидов синтезируют дрожжи и мицелиальные грибы. Простые липиды находят применение как технологические смазки в процессах холодной и тепловой обработки металлов. Продуцентами сложных липидов являются в основном бактерии. Сложные липиды делятся на две группы: фосфолипиды и гликолипиды. Фосфолипиды (фосфоглицериды и сфинголипиды) входят в состав различных клеточных мембран и принимают участие в переносе электронов. Их молекулы полярны и при pH 7,0 фосфатная группа несет отрицательный заряд. Концентрат фосфолипидов находит применение в качестве антикоррозийной присадки к маслам и как добавка при флотации различных минералов. Гликолипиды в отличие от фосфолипидов не содержат молекулы фосфорной кислоты, но также являются сильнополярными соединениями за счет наличия в молекуле гидрофильных углеводных групп (остатков глюкозы, маннозы, галактозы и др.). К производным липидов относят жирные кислоты, спирты, углеводороды, витамины Д, Е и К. Жирные кислоты представлены насыщенными и ненасыщенными с одной двойной связью кислотами нормального строения и четным числом углеродных атомов (пальмитиновая, стеариновая, олеиновая). Среди диеновых жирных кислот можно выделить линолевую. Двойные связи в ненасыщенных жирных кислотах микробных липидов часто располагаются так, что делят их на части, число углеродных атомов в которых кратно трем. Очищенные монокарбоновые кислоты с числом углеродных атомов 14–18 находят широкое применение в мыловаренной, шинной, химической, лакокрасочной и других отраслях промышленности. Спирты, присутствующие в липидах, делятся на три группы: спирты с прямой цепью, спирты с β-ионовым кольцом, включающие витамин А и каротиноиды, а также стерины — компоненты неомыляемой части липидов (например, эргостерин, облучение которого ультрафиолетовым светом позволяет получать витамин Д2). Для промышленного использования важное значение имеет способность усиленно накапливать липиды. Этой способностью обладают немногие микроорганизмы, в первую очередь дрожжи. Процесс образования липидов у большинства дрожжей состоит из двух четко разграниченных стадий: — первая характеризуется быстрым образованием белка в условиях усиленного снабжения культуры азотом и сопровождается медленным накоплением липидов (в основном глицерофосфатов и нейтральных жиров); — вторая — прекращением роста дрожжей и усиленным накоплением липидов (в основном нейтральных). Типичными липидообразователями являются дрожжи Cryptococcus terricolus. Они могут синтезировать большое количество липидов (до 60 % от сухой массы) в любых условиях, даже наиболее благоприятных для синтеза белка. Из других липидообразующих дрожжей промышленный интерес представляют дрожжи С. guilliermondii, утилизирующие алканы. Они синтезируют в основном фосфолипиды. Накапливают большие количества липидов и активно развиваются на углеводных субстратах (на мелассе, гидролизатах торфа и древесины) также дрожжи видов Lipomyces lipoferus и Rhodotorula gracilis. У этих видов дрожжей липогенез сильно зависит от условий культивирования. Эти продуценты накапливают значительные количества (до 70 %) триацилглицеридов. Микроскопические грибы пока не получили большого распространения в получении липидов, хотя жир грибов по своему составу близок к растительному. Выход жиров у Asp.terreus, например, на углеводных средах достигает 51 % от абсолют но сухого веса (АСВ). Липидный состав грибов представлен в основном нейтральными жирами и фосфолипидами. Липиды, синтезируемые бактериями, своеобразны по своему составу, так как включают в основном сложные липиды, тогда как нейтральные жиры составляют незначительную часть биомассы. При этом бактерии производят разнообразные жирные кислоты (содержащие от 10 до 20 атомов углерода), что важно для промышленного получения специфических жирных кислот. Водоросли перспективны для культивирования в качестве липидообразователей, так как не нуждаются в органическом источнике углерода. Химический состав (соотношение белков и жиров) водорослей также сильно варьирует в зависимости от содержания в среде азота. Недостатки — малая скорость роста и накопление токсических соединений в клетках, — ограничивают промышленное применение. Итак, основную роль в процессе биосинтеза липидов играют различные штаммы дрожжей. Они используют те же источники сырья, что и для получения кормового белка, причем от ценности углеродного питания зависят выход биомассы, количество и состав синтезируемых липидов. Для обеспечения направленного биосинтеза липидов в питательной среде употребляются легкоассимилируемые источники азота. На сдвиг биосинтеза в сторону образования липидов или белка влияет соотношение углерода и азота в среде. Так, повышение концентрации азота вызывает снижение липидообразования, а недостаток азота при обеспеченности углеродом ведет к понижению выхода белковых веществ и высокому процентному содержанию жира. Установлено, что оптимальное соотношение N: C тем меньше, чем труднодоступнее для дрожжей источник углерода. Обычно для углеводородного сырья соотношение N: C = 1:30, а для углеводного — 1:40. Накопление липидов возможно только при наличии в среде фосфора. При его недостатке источники углерода используются не полностью, при избытке — накапливаются нелипидные продукты. На фракционный состав липидов изменение содержания фосфора влияния не оказывает. Воздействие остальных элементов среды (микро- и макроэлементов) сказывается на интенсивности роста дрожжей и скорости утилизации источника углерода, что влияет и на количество накопленных липидов, но не на их качество. На фракционный состав синтезируемых липидов оказывают другие условия культивирования: аэрация, pH и температура. От интенсивности аэрации зависит синтез фосфоглицеридов, жирных кислот и триацилглицеридов. При недостаточной аэрации липиды содержат в 4 раза меньше триацилглицеридов, в 2 раза больше фосфоглицеридов и в 8 раз больше жирных кислот, чем при нормальной. При интенсификации аэрации возрастает степень ненасыщенности липидов и увеличивается относительное количество всех групп ненасыщенных кислот. Повышение pH среды ведет к увеличению содержания фосфоглицеридов и жирных кислот при одновременном снижении количества триацилглицеридов. Оптимальные температуры роста и липидообразования для клеток совпадают, причем содержание липидов не зависит от температуры культивирования. Однако, регулируя температуру, можно создавать разные соотношения насыщенных и ненасыщенных жирных кислот в составе фосфолипидных мембран. Для углеводных субстратов наиболее отработана технология получения липидов на гидролизатах торфа и древесины. Как показали исследования, соотношение гидролизатов торфа и древесины 1:4 обеспечивает наибольший выход биомассы в стадии культивирования (до 10 г/л) при максимальном содержании липидов (до 51 % от АСВ) и высоком коэффициенте усвоения субстрата (до 0,54). Из 1 тонны абсолютно сухого торфа после его гидролиза и ферментации можно получить 50–70 кг микробного жира с преимущественным содержанием триацилглицеридов.
ТЕХНОЛОГИЯ ФЕРМЕНТНЫХ ПРЕПАРАТОВ
Ферменты, получаемые промышленным способом, их применение Классификация ферментов основана на механизме их действия и включает 6 классов. Ферменты как биокатализаторы обладают рядом уникальных свойств, например, таких как высокая каталитическая активность и избирательность действия. В ряде случаев ферменты обладают абсолютной специфичностью, катализируя превращение только одного вещества. Для каждого фермента существует свой оптимум pH, при котором его каталитическое действие максимально. При резком изменении pH ферменты инактивируются из-за необратимой денатурации. Ускорение реакции при повышении температуры также лимитировано определенными пределами, поскольку уже при температуре 40–50 °C многие ферменты денатурируют. Эти свойства ферментов приходится учитывать при разработке технологии нового препарата. Поскольку ферменты — вещества белковой природы, в смеси с другими белками их количество определить практически невозможно[61]. Наличие фермента в препарате может быть установлено лишь по протеканию той реакции, которую катализирует фермент. При этом количественную оценку содержания фермента можно дать, определив либо количество образовавшихся продуктов реакции, либо количество израсходовавшегося субстрата. За единицу активности фермента принимают то его количество, которое катализирует превращение одного микромоля субстрата в 1 минуту при заданных стандартных условиях — стандартная единица активности. По решению Международного биохимического союза активность решено определять при t = 30 °C по начальной скорости реакции, когда концентрация насыщения фермента и временная зависимость близка к кинетике реакции нулевого порядка. Остальные параметры реакции индивидуальны для каждого фермента. Активность ферментного препарата выражается в микромолях субстрата, прореагировавшего под действием 1 мл ферментного раствора или 1 грамма препарата в оптимальных условиях за 1 минуту. Если ферментный препарат не содержит балласта, то его активность выражается в тех же стандартных единицах на 1 мг фермента. Если же есть балласт, то активность считается на 1 мг белка в ферментном препарате. Активность выпускаемого препарата — важнейший нормируемый показатель качества. Основную часть ферментов, получаемых промышленным способом, составляют гидролазы. К ним относятся, в первую очередь амилолитические ферменты: α-амилаза, β-амилаза, глюкоамилаза. Их основная функция — гидролиз крахмала и гликогена. Крахмал при гидролизе расщепляется на декстрины, а затем до глюкозы. Эти ферменты применяются в спиртовой промышленности, хлебопечении. Протеолитические ферменты образуют класс пептидгидролаз. Их действие заключается в ускорении гидролиза пептидных связей в белках и пептидах. Важная их особенность — селективный характер действия на пептидные связи в белковой молекуле. Например, пепсин действует только на связь с ароматическими аминокислотами, трипсин — на связь между аргинином и лизином. В промышленности протеолитические ферменты классифицируют по способности проявлять активность в определенной области pH: — pH 1.5–3.7 — кислые протеазы; — pH 6.5–7.5 — протеазы; — pH > 8.0 — щелочные протеазы. Протеазы находят широчайшее применение в разных отраслях промышленности: — мясная — для смягчения мяса; — кожевенная — смягчение шкур; — кинопроизводство — растворение желатинового слоя при регенерации пленок; — парфюмерная — добавки в зубную пасту, кремы, лосьоны; — производство моющих средств — добавки для удаления загрязнений белковой природы; — медицина — при лечении воспалительных процессов, тромбозов и т. д. Пектолитические ферменты уменьшают молекулярную массу и снижают вязкость пектиновых веществ. Пектиназы делятся на две группы — гидролазы и трансэлиминазы. Гидралазы отщепляют метильные остатки или разрывают гликозидные связи. Трансэлиминазы ускоряют негидролитическое расщепление пектиновых веществ с образованием двойных связей. Применяются в текстильной промышленности (вымачивание льна перед переработкой), в виноделии — осветление вин, а также при консервировании фруктовых соков. Целлюлолитические ферменты очень специфичны, их действие проявляется в деполимеризации молекул целлюлозы. Обычно используются в виде комплекса, доводящего гидролиз целлюлозы до глюкозы (в гидролизной промышленности). В медицинской промышленности их используют для выделения стероидов из растений, в пищевой — для улучшения качества растительных масел, в сельском хозяйстве — как добавки в комбикорма для жвачных животных. Существует ряд факторов, влияющих на биосинтез ферментов. В первую очередь, к ним относится генетический. Состав и количество синтезируемых ферментов наследственно детерминированы. Применяя мутагены можно изменить генетические свойства микроорганизмов и получить штаммы с ценными для промышленности свойствами. К мутагенным факторам относятся ионизирующее и неионизирующее излучения, изотопы, антибиотики, другие химические соединения, преобразующие наследственные элементы клетки. Несмотря на определяющую роль генетического фактора в биосинтезе ферментов, производительность биотехнологических процессов зависит и от состава питательной среды. При этом важно не только наличие источников основных питательных веществ, но и веществ, играющих роль индукторов или репрессоров биосинтеза данного конкретного фермента или их групп. Механизм этого явления еще не вполне изучен, но сам факт должен учитываться при выборе технологии. Рассмотрим несколько примеров. Фермент липаза почти не синтезируется грибом Asp.awamori на среде без индуктора, добавление жира кашалота усиливает биосинтез фермента в сотни раз. При добавлении же в среду крахмала и при полном исключении минерального фосфора интенсивно синтезируется фосфатаза. Не только наличие индуктора способно увеличивать выход фермента. Важную роль играет состав питательной среды и условия культивирования. При разработке процесса биосинтеза а-амилазы культурой Asp.oryzae замена сахарозы (как источника углерода) на крахмал увеличила активность фермента в 3 раза, добавление солодового экстракта (из проросших семян злаковых) ещё в 10 раз, а повышение концентрации основных элементов питательной среды на 50 % — ещё в 2 раза. Для интенсификации процесса роста и синтеза ферментов добавляют различные факторы роста, например, аминокислоты, пуриновые основания и их производные, РНК и продукты её гидролиза. В качестве источника углерода используют крахмал, кукурузный экстракт, соевую муку, гидролизаты биомассы дрожжей. Микроорганизмы могут утилизировать и минеральные источники азота. В состав питательных сред входят и ионы Mg, Mn, Zn, Fe, Си и др. металлов. Механизм действия большинства из них неизвестен. Некоторые входят в состав фермента. Ионы Са повышают устойчивость а-амилазы, ионы Fe и Mg активизируют и стабилизируют протеолитические ферменты. Оптимальный состав питательной среды для каждого продуцента может быть определен двумя способами: эмпирический и построение математической модели с использованием компьютера. Последний, естественно, предпочтительнее. По характеру культивирования все технологические процессы производства ферментных препаратов делятся на две большие группы: глубинный и поверхностный методы.
Глубинный метод культивирования продуцентов ферментов В этом случае микроорганизмы выращиваются в жидкой питательной среде. Технически более совершенен, чем поверхностный, так как легко поддается автоматизации и механизации. Концентрация фермента в среде при глубинном культивировании обычно значительно ниже, чем в водных экстрактах поверхностной культуры. Это вызывает необходимость предварительного концентрирования фильтрата перед его выделением. При глубинном культивировании продуцентов ферментов выделяют, как и в любом биотехнологическом процессе, 5 этапов. 1. Приготовление питательных сред зависит от состава компонентов. Некоторые предварительно измельчают, отваривают или гидролитически расщепляют. Готовые к растворению компоненты подают при постоянном помешивании в емкость для приготовления среды в определенной последовательности. Стерилизацию среды проводят либо путем микрофильтрации с помощью полупроницаемых мембран, либо при помощи высоких температур. Время обработки в этом случае зависит как от интенсивности фактора, так и от уровня обсемененности объекта. Стерилизуются также все коммуникации и аппараты. Воздух очищается до и после аэрирования. До — потому что содержит частицы пыли органической и неорганической природы, после — так как несет клетки продуцента. 2. Получение засевного материала. Для засева питательной среды материал готовят также глубинным методом. Вид его зависит от продуцента: для грибов это мицелиальная вегетативная масса, для бактерий — молодая растущая культура на начальной стадии спорообразования. Получение посевного материала состоит в увеличении массы продуцента в 3–4 стадии. Объем посевного материала зависит от физиологических особенностей продуцента. Если продуцент размножается толь ко вегетативно, он резко возрастает (до 5-20 %). Если же происходит обильное спороношение — сокращается до 1 %. 3. Производственное культивирование. Биосинтез ферментов в глубинной культуре протекает в течение 2–4 суток при непрерывной подаче воздуха и перемешивании. Высокая концентрация питательных веществ на первых этапах могут тормозить рост биомассы продуцента, поэтому часто свежая среда или некоторые её компоненты вводятся в ферментер на стадии активного роста. Температурный оптимум находится в интервале 22–32 °C. В современных технологических процессах ведется непрерывное автоматическое определение содержания в среде углеводов, количества образовавшихся метаболитов и концентрации клеток. Данные поступают в ЭВМ, которая определяет стратегию коррекции процесса и автоматически регулирует его. Этим достигается максимальная производительность и наилучшее качество продуктов. 4. Выделение. В мицелии трёхсуточной культуры обычно остается не более 15 % ферментов. Остальные выделяются в окружающую клетки жидкую среду. В этом случае препараты ферментов выделяют из фильтратов после отделения биомассы. 5. Получение товарной формы.
Поверхностный метод культивирования продуцентов ферментов При поверхностном методе культура растет на поверхности твердой увлажненной питательной среды[62]. Мицелий полностью обволакивает и довольно прочно скрепляет твердые частицы субстрата, из которого получают питательные вещества. Поскольку для дыхания клетки используют кислород, то среда должна быть рыхлой, а слой культуры-продуцента небольшим. Выращивание производственной культуры происходит обычно в асептических условиях, но среду и кюветы необходимо простерилизовать. Перед каждой новой загрузкой также необходима стерилизация оборудования. Преимущества поверхностной культуры: значительно более высокая конечная концентрация фермента на единицу массу среды (при осахаривании крахмала 5 кг поверхностной культуры заменяют 100 кг культуральной жидкости), поверхностная культура относительно легко высушивается, легко переводится в товарную форму. Посевной материал может быть трёх видов: — культура, выросшая на твердой питательной среде; — споровый материал; — мицелиальная культура, выращенная глубинным способом. В три этапа получают и посевную культуру. Сначала музейную культуру продуцента пересевают на 1–1.5 г увлажненных стерильных пшеничных отрубей в пробирку и выращивают в термостате до обильного спорообразования. Второй этап — аналогично, но в колбах, третий — в сосудах с 500 г среды. Основу питательной среды составляют пшеничные отруби, как источник необходимых питательных и ростовых веществ. Кроме того, они создают необходимую структуру среды. Для повышения активности ферментов к отрубям можно добавлять свекловичный жом, соевый шрот, крахмал, растительные отходы. Стерилизуют среду острым паром при помешивании (температура — 105–140 °C, время 60–90 минут). После этого среду засевают и раскладывают ровным слоем в стерильных кюветах. Кюветы помещают в растильные камеры. Культивируют в течение 36–48 часов. Рост делится на три периода, примерно равных по времени. Сначала происходит набухание конидий и их прорастание (температура не ниже 28 °C), затем рост мицелия в виде пушка серовато-белого цвета (необходимо выводить выделяемое тепло) и образование конидий. Для создания благоприятных условий роста и развития продуцента необходима аэрация и поддержание оптимальной влажности (55–70 %). Выросшая в неподвижном слое при поверхностном культивировании культура представляет корж из набухших частиц среды, плотно связанных сросшимся мицелием. Массу размельчают до гранул 5–5 мм. Культуру высушивают до 10–12 % влажности при температурах не выше 40 °C, не долее 30 минут. Иногда препарат применяют прямо в неочищенном виде — в кожевенной и спиртовой промышленности. В пищевой и особенно медицинской промышленности используются ферменты только высокой степени очистки. Схема очистки сводится к следующему: — освобождение от нерастворимых веществ; — освобождение от сопутствующих растворимых веществ; — фракционирование (как правило, хроматографическими методами). Для выделения фермента из поверхностной культуры необходима экстракция. Как правило, экстраген — вода. При этом в раствор переходят сахара, продукты гидролиза пектиновых веществ и целлюлозы. Стадию выделения и очистки завершает сушка. После сушки препарат должен содержать не более 6–8 % влаги, тогда он может в герметичной упаковке храниться до года без потери активности. Стандартизация ферментного препарата — доводка активности фермента до стандартной, соответствующей требованиям ГОСТ. Для этого используются различные нейтральные наполнители — крахмал, лактоза и др. Учитывая огромные перспективы применения ферментных препаратов в различных отраслях промышленности и сельского хозяйства, медицине, можно сделать заключение о необходимости расширения исследований в этой области для оптимизации технологии и гарантийного получения высокоактивных и стабильных препаратов микробных ферментов.
БИОТЕХНОЛОГИЯ ПРЕПАРАТОВ ДЛЯ СЕЛЬСКОГО ХОЗЯЙСТВА
Препараты, производимые для сельского хозяйства, делятся на 3 группы: — энтомопатогенные препараты; — бактериальные удобрения; — антибиотики. Отечественное биотехнологическое производство выпускает 3 группы энтомопатогенных препаратов: 1. Бактериальные препараты на основе Bacillus thuringiensis — энтобактерин-3, дендробациллин, инсектин, токсобактерин. 2. Грибной препарат боверин на основе гриба Beauveria bassiana. 3. Препараты на основе вирусов ядерного полиэдра (вирин-ЭНШ, вирин-ЭКС и ДР.). Все микробные патогены выпускаются в виде смачивающих порошков, паст, реже — гранул, эмульсии спор и кристаллов. При непосредственном применении предполагается использование различных добавок в виде растворителей, прилипателей, способствующих повышению их эффективности. Технология получения бактериальных энтомопатогенных препаратов. Наибольшее распространение среди промышленно выпускаемых микробных патогенов получили бактериальные препараты. Их отличительными особенностями являются высокая вирулентность по отношению к насекомым-вредителям, безопасность для окружающей флоры и фауны, достаточно высокая скорость воздействия на вредителей и др. В настоящее время производятся препараты против более 160 видов насекомых.
Бактериальные энтомопатогенные препараты Из всех энтомопатогенных бактерий наиболее исследованы грамположительные бактерии Вас. thuringiensis. Она не только разрушает насекомое, попадая внутрь, но и продуцирует ряд токсичных продуктов. Среди этих токсичных продуктов выделяют 4 компонента: — α-экзотоксин, или фосфолипаза С, — продукт растущих клеток бактерий. Токсическое действие фермента связывают с индуцируемым им распадом незаменимых фосфолипидов в ткани насекомого, что приводит к гибели последнего. — β-экзотоксин — накапливается в культуральной жидкости при росте клеток. Считают, что молекула β-токсина состоит из нуклеотида, связанного через рибозу и глюкозу с аллослизевой кислотой. Его действие, видимо, обусловлено ингибированием нуклеотидазы и ДНК-зависимой РНК-полимеразы, связанных с АТФ, что приводит к прекращению синтеза РНК. По сравнению с другими токсинами действует медленнее, в основном при переходе от одного цикла развития к другому. По наблюдениям, р-экзотоксин — мутаген, поражающий генетический аппарат особей. — γ-экзотоксин — малоизученный компонент, неидентифицированный фермент (или группа ферментов). — δ-эндотоксин — параспоральный кристаллический эндотоксин. Образуется в процессе споруляции бактерии в противоположной от формирующейся споры части бактерии. На завершающей стадии спорообразования токсин приобретает форму 8-гранного кристалла. Кристаллы состоят из белка, аминокислотный состав которого близок для различных штаммов. Доказано, что кристаллический белок в кишечнике восприимчивых насекомых распадается на молекулы протоксина. Протоксин под действием протеиназ распадается на токсические фрагменты. Различие в восприимчивости некоторых видов насекомых к действию кристалла, по-видимому, связано с присутствием специальных кишечных протеаз, осуществляющих гидролиз кристаллов in vivo. Такими протеазами обладают не все насекомые, отсюда и избирательность действия 5-токсина. Чтобы насекомое погибло, кристаллы должны попасть в его организм. После поглощения кристаллов гусеницы перестают питаться. Первичным местом действия 5-токсина является средний отдел кишечника. В зависимости от реакции на кристаллы насекомые делятся на три группы: — характерен общий паралич; — паралич среднего отдела кишечника; — реакция на препарат в целом: гибель в результате прорастания спор и последующего размножения бактерий. Бактерии Вас. thuringiensis антагонистичны к 130 видам насекомых. Наибольший эффект достигается при применении препаратов этой группы против листогрызущих вредителей. Наиболее распространенные препараты на основе различных вариаций Вас. thuringiensis: энтобактерин, инсектин, алестин, экзотоксин, токсобактерин, дендробациллин, битоксибациллин. Промышленное производство энтомопатогенных бактерий (ЭБ) заключается в глубоком культивировании. При этом ставится задача получения максимального титра клеток в культуральной жидкости и накопления токсина. Требования к промышленным штаммам ЭБ: принадлежность штамма к определенному серотипу, высокая вирулентность и продуктивность на промышленных средах, устойчивость к комплексу фагов и т. д. Технология производства включает все стадии, типичные для любого биотехнологического производства. Температуру культивирования на всех стадиях поддерживают постоянной (28–30 °C), продолжительность ферментации составляет 35–40 часов. Используют дрожже-полисахаридную среду, содержащую в процентах: кормовые дрожжи — 2–3; кукурузную муку — 1–1.5; кашалотовый жир — 1. Перед началом культивирования pH составляет около 6.3, к концу ферментации — повышается до 8.0–8.5, что может привести к разрушению кристаллов на более мелкие фрагменты и затруднить их выделение. Чтобы предотвратить это, культуральную жидкость перед переработкой подкисляют до 6.0–6.2. Культивирование заканчивают при степени споруляции 90–95 % и титре спор не менее 109 в 1 мл. После сепарации культуральной жидкости получают пасту влажностью 85 % с выходом около 100 кг в 1 кубометре культуральной жидкости и титром порядка 20*109 спор в 1 грамме. Фугат можно употребить для приготовления питательной среды, но не более 1–2 раз, так как в культуральной жидкости накапливаются вещества, ингибирующие развитие культуры. Фугат находит свое применение в качестве сырья при производстве кормовых дрожжей, что обеспечивает сокращение промышленных стоков и снижает расход воды. Пасту перемешивают в течение получаса для однородного распределения спор и кристаллов и отбирают пробы на проверку титра, влажности, вирулентности, наличия фага. Конечный продукт — смачивающий порошок или стабилизированная паста. Первый получают путем высушивания увлажненной пасты на распылительной сушке. Готовый препарат фасуют по 20 кг в четырехслойные крафт-мешки с полиэтиленовым вкладышем. Вторую — внесением в пату КМЦ. При смешении молекулы КМЦ сорбируют белковые кристаллы и споры, заряжая их отрицательно, что способствует равномерному распределению активного начала по всему объёму и увеличению срока хранения. Готовый препарат — вязкая жидкость кремового или светло-серого цвета, без запаха, не замерзающую при хранении. Препарат предназначен для борьбы с садово-огородными вредителями, эффективен против 60 видов насекомых. Применяют путем опрыскивания растений водной эмульсией в период активного роста вредителя. Основная масса вредителей погибает в течение 2-10 дней. На 1 га расходуют: для овощных культур 1–3 кг, садовых — 3–5 кг.
Грибные энтомопатогенные препараты Энтомопатогенные препараты на основе микроскопических грибов вызывают у насекомых микозы. Грибы обладают рядом особенностей: — поражение происходит через кутикулу; — насекомые поражаются в фазе развития куколки и имаго; — большая скорость роста и огромная репродуктивная способность, в виде спор могут длительное время находится в природе без снижения энтомопатогенной активности; — высокая специфичность, вирулентность сильно зависит от штамма гриба. Действие грибного препарата на насекомое начинается с проникновения споры в полость тела через кожные покровы. Попав в тело, спора прорастает в гифу, затем разрастается мицелий, от которого отчленяются конидии. Оказавшись в теле, конидии циркулируют в гемолимфе. Уже на этой стадии возможно поражение насекомого вследствие выделения некоторыми штаммами значительного количества токсинов. В отсутствие токсина мицелий постепенно заполняет все тело насекомого, прежде всего поражается мышечная ткань. Рост гриба продолжается до тех пор, пока все ткани не будут разрушены. Могут образовываться конидиеносцы, прорывающие кутикулу и обволакивающие мертвую личинку. В промышленном производстве используются отдельные штаммы в основном трех родов: Beaveria, Metarrhizium, Entomophtora. В нашей стране освоено промышленное производство препарата боверина на основе B.bassiana. Готовый препарат — порошок кремового или белого цвета, содержащий в 1 г от 1.5 до 6 млрд. конидиоспор. Препарат безвреден для теплокровных животных и человека, не вызывает ожогов у растений. Получать боверин, можно используя, как поверхностное, так и глубинное культивирование. Первый способ более трудоемок и длителен, поэтому имеет ограниченное значение. Производство конидиоспор при выращивании его в жидкой среде также непростая задача. Основная трудность в том, что при этом способе культивирования гриб размножается вегетативно, образуя гонидии. Гонидии по вирулентности не уступают конидиям, однако неустойчивы к высоким температура на стадии высушивания. При традиционной распылительной сушке погибает 90 % гонидиоспор и 20–50 % конидиоспор. Проблема решается подбором питательной среды и условий ферментации, обеспечивающих переход 90 % выращенных клеток в конидиоспоры. Технология получения боверина методом глубинного культивирования включает обычные стадии. Питательная среда содержит в процентах: дрожжи кормовые — 2, крахмал — 1, хлорид натрия — 0.2, хлорид марганца — 0.01, хлорид кальция — 0.05. Последний компонент обеспечивает устойчивость конидий к неблагоприятным факторам, поэтому его содержание может сильно варьировать (до 5 %). Культивируют при pH 4.5–5.6, температуре 25–28 °C 3–4 суток в условиях постоянного перемешивания и аэрации. В среде необходимо также наличие аминного азота, так как его недостаток снижает скорость роста культуры и процент образования конидиоспор, избыток ведет к образованию гонидий. Культуральную жидкость подвергают сепарации и фильтрованию, после чего пастусушат на распылительной сушке.
Вирусные энтомопатогенные препараты Из всех энтомопатогенных препаратов вирусные обладают наибольшей специфичностью по отношению к насекомому-хозяину. Они поражают не более одного вида. Их ярко выраженная специфичность обуславливает практическую безвредность вирусных препаратов для человека, флоры и фауны. Вирусы отличает высокая устойчивость к неблагоприятным факторам окружающей среды, они способны сохранять активность в течение 10–15 лет, находясь вне насекомого. Заражение вирусом происходит при питании вредителя. Попавшие в кишечник тельца-включения при щелочных значениях pH разрушаются. Освобожденные вирионы проникают через стенку кишечника в клетки, где в ядрах происходит репликация вирусов. Высвободившиеся вирусы заражают другие клетки, что в итоге приводит к гибели насекомого. Отличительной особенностью вирусов является то, что они могут размножаться только в живой ткани. Это создает определенные трудности в организации промышленного производства, так как технология размножения вирусов должна быть связана с использованием живых насекомых-хозяев. В нашей стране осуществляется выпуск трех вирусных энтомопатогенных препаратов: вирин-ЭКС (против капустной совки), ЭНШ (против непарного шелкопряда) и АББ (против американской белой бабочки). Производство любого из вирусных препаратов начинают с разведения насекомого-хозяина на искусственных питательных средах, обеспечивающих их физиологически здоровое состояние. На определенной стадии развития (обычно на стадии гусеницы) насекомых заражают, добавляя вирусную суспензию к корму. При этом инокулят предварительно получают от нескольких больных личинок. После заражения насекомых выдерживают в строго определенных условиях, обеспечивающих максимальное накопление вируса в тканях. Через 7–9 суток собирают мертвые и отмирающие личинки, подсушивают при температуре 33–35 °C, измельчают механическим способом для вывода телец-включений из тканей. К полученной массе добавляют физиологический раствор или дистиллированную воду из расчета 1 мл на гусеницу, взвесь полученных тканей фильтруют. При производстве вирин-ЭКС полиэдры осаждают из фильтрата центрифугированием. Осадок суспендируют в минимальном количестве дистиллированной воды и добавляют простерилизованный глицерин до титра 1 млрд. полиэдров в 1 мл. Готовый препарат разливают во флаконы. При производстве вирин-ЭНШ в фильтрат добавляют лактозу, а после перемешивания ацетон в соотношении 4:1 к объему суспензии. После отстаивания надосадочную жидкость сливают, осадок подсушивают до полного удаления ацетона. Если препарат планируется выпускать в виде порошка, то сухой осадок смешивают с мелкодисперсным наполнителем (каолином, например) до получения титра полиэдров 1 млрд. полиэдров в 1 грамме. Масляную форму препарата получают путем диспергирования осадка в стерильном 50 % растворе глицерина до титра 2 млрд. полиэдров в 1 мл, а затем добавляют равный объем солярового масла, перемешивают и разливают по флаконам. Вирусные энтомопатогенные препараты применяют путем внесения полиэдров в плотные популяции насекомых-вредителей с целью возникновения в них эпизоотий. Данный способ обработки предполагает внесение небольших количеств препарата. В другом случае опрыскивание или опыление производят на зараженных участках в период рождения личинок или на ранних стадиях их развития.
Бактериальные удобрения на основе клубеньковых бактерий Микрофлора почвы оказывает непосредственное влияние на её плодородие и, как следствие, на урожайность растений. Почвенные микроорганизмы в процессе роста и развития улучшают структуру почвы, накапливают в ней питательные вещества, минерализуют различные органические соединения, превращая их в легко усвояемые растением компоненты питания. Для стимуляции этих процессов применяют различные бактериальные удобрения, обогащающие ризосферу растений полезными микроорганизмами. Микроорганизмы, используемые для производства бактериальных препаратов, способствуют снабжению растений не только элементами минерального питания, но и физиологически активными веществами (фитогормонами, витаминами и др.). В настоящее время выпускают[63] такие бактериальные удобрения, как нитрагин, ризоторфин, азотобактерин, фосфобактерин, экстрасол. Отечественная промышленность выпускает два вида препаратов клубеньковых бактерий: нитрагин и ризоторфин. Оба препарата производятся на основе активных жизнеспособных клубеньковых бактерий из рода Rhizobium. Эти бактерии в симбиозе с бобовыми культурами способны фиксировать свободный азот атмосферы, превращая его в соединения, легкоусвояемые растением. Бактерии рода Rhizobium — строгие аэробы. Среди них различают активные, малоактивные и неактивные культуры. Критерием активности клубеньковых бактерий служит их способность в симбиозе с бобовым растением фиксировать атмосферный азот и использовать его в виде соединений для корневого питания растений. Фиксация атмосферного азота возможна только в клубеньках, образующихся на корнях растений. Возникают они при инфицировании корневой системы бактериями из рода Rhizobium. Заражение корневой системы происходит через молодые корневые волоски. После внедрения бактерии прорастают внутри них до самого основания в виде инфекционной нити. Выросшие нити проникают сквозь стенки эпидермиса в кору корня, разветвляются и распределяются по клетками коры. При этом индуцируется деление клеток хозяина и разрастание тканей. В месте локализации бактерий на корне растения-хозяина образуются клубеньки, в которых бактерии быстро размножаются и располагаются по отдельности или группами в цитоплазме растительных клеток. Сами бактериальные клетки увеличиваются в несколько раз и меняют окраску. Если клубеньки имеют красноватую или розовую окраску, обусловленную наличием пигмента легоглобина (леггемоглобина) — аналог гемоглобина крови животных, то они способны фиксировать молекулярный азот. Неокрашенные ("пустые") или имеющие зеленоватую окраску клубеньки не фиксируют азот. Бактерии, находящиеся в клубеньках, синтезируют ферментную систему с нитрогеназной активностью, восстанавливающую молекулярный азот до аммиака. Ассимиляция аммиака происходит, в основном, путем вовлечения его в ряд ферментативных превращений, приводящих к образованию глутамина и глутаминовой кислоты, идущих в дальнейшем на биосинтез белка. Помимо критерия активности в характеристике клубеньковых бактерий используют критерий вирулентности. Он характеризует способность микроорганизма вступать в симбиоз с бобовым растением, то есть проникать через корневые волоски внутрь корня и вызывать образование клубеньков. Большое значение имеет скорость такого проникновения. В симбиотическом комплексе растение — Rhizobium бактерии обеспечиваются питательными веществами, а сами снабжают растение азотистым питанием. С вирулентностью связана и видовая избирательность, которая характеризует способность данного вида бактерий к симбиозу с определенным видом бобового растения. Классификация различных видов Rhizobium учитывает растение-хозяина, например: Rhizobium phaseoli — для фасоли, Rhizobium lupini — для люпина, сараделлы и т. д. Вирулентность и видоспецифичность взаимосвязаны и не являются постоянными свойствами штамма. Задачей производства бактериальных удобрения является максимальное накопление жизнеспособных клеток, сохранение их жизнеспособности на всех стадиях технологического процесса, приготовление на их основе готовых форм препарата с сохранением активности в течение гарантийного срока хранения. Отечественная промышленность выпускает два вида нитрагина: почвенный и сухой. Впервые культура клубеньковых бактерий на почвенном субстрате была приготовлена в 1911 году на бактериально-агрономической станции в Москве. В настоящее время его производство имеет ограниченное значение, так как технология довольно сложна и трудоёмка при выполнении отдельных операций. Более перспективна технология производства сухого нитрагина. Сухой нитрагин — порошок светло-серого цвета, содержащий в 1 г не менее 9 млрд. жизнеспособных бактерий в смеси с наполнителем. Влажность не превышает 5–7 %. Промышленное производство имеет типичную схему. Необходимо отметить, что важно подбирать штаммы, устойчивые к высушиванию. Для производства посевного материала исходную культуру клубеньковых бактерий выращивают на агаризованной среде, содержащей отвар бобовых семян, 2 % агара и 1 % сахарозы, затем культуру размножают в колбах на жидкой питательной среде в течение 1–2 суток при 28–30 °C и pH 6.5–7.5. На всех этапах промышленного культивирования применяют питательную среду, включающую такие компоненты, как меласса, кукурузный экстракт, минеральные соли в виде сульфатов аммония и магния, мел, хлорид натрия и двузамещенный фосфат калия. Основная ферментация идет при тех же условиях в течение 2–3 суток. Готовую культуральную жидкость сепарируют, получается биомасса в виде пасты с влажностью 70–80 %. Пасту смешивают с защитной средой, содержащей тиомочевину и мелассу (1:20) и направляют на высушивание. Сушат путем сублимации (в вакуум-сушильных шкафах). Высушенную биомассу размалывают. Производительнее высушивание в распылительных сушках, но при этом 75 % клеток теряют жизнеспособность. Препараты сухого нитрагина фасуют и герметизируют в полиэтиленовые пакеты по 0.2–1 кг, хранят при температуре 15 °C не более 6 месяцев. Семена опудривают перед посевом. Внесение нитрагина повышает урожайность в среднем на 15–25 %. Препарат клубеньковых бактерий может выпускаться и в виде ризоторфина. Впервые торфяной препарат клубеньковых бактерий был приготовлен в 30-х годах, но технология была создана в 1973-77 гг. Для приготовления ризоторфина торф сушат при температуре не выше 100 °C и размалывают в порошок. Наиболее эффективным способом стерилизации является облучение его гамма-лучами. Перед стерилизацией размолотый, нейтрализованный мелом и увлажненный до 30–40 % торф расфасовывают в полиэтиленовые пакеты. Затем его облучают и заражают клубеньковыми бактериями, используя шприц, с помощью которого впрыскивается питательная среда, содержащая клубеньковые бактерии. Прокол после внесения бактерий заклеивается липкой лентой. Каждый грамм ризоторфина должен содержать не менее 2.5 млрд. жизнеспособных клеток с высокой конкурентоспособностью и интенсивной азотфиксацией. Препарат хранят при температуре 5–6 °C и влажности воздуха 40–55 %. Пакеты могут быть весом от 0.2 до 1.0 кг. Доза препарата составляет 200 г на га. Заражение семян производят следующем образом: ризоторфин разбавляют водой и процеживают через двойной слой марли. Полученной суспензией обрабатывают семена. Семена высевают в день обработки или на следую- Обработка семян бобовых культур прочно вошла в мировую сельскохозяйственную практику. Крупнейшими производителями таких препаратов являются США и Австралия.
Технология получения азотобактерина Азотобактерин — бактериальное удобрение, содержащее свободноживущий почвенный микроорганизм Azotobacter chroococcum, способный фиксировать до 20 мг атмосферного азота на 1 г использованного сахара. Внесенные в качестве удобрения в почву бактерии также выделяют биологически активные вещества (никотиновую и пантотеновую кислоты, пиридоксин, биотин, гетероауксин, гиббереллин и др.). Эти вещества стимулируют рост растений. Кроме того, продуцируемые Azotobacter фунгицидные вещества из группы анисомицина угнетают развитие некоторых нежелательных микроскопических грибов в ризосфере растения. Все виды Azotobacter строгие аэробы. Чувствительны к содержанию в среде фосфора и развиваются лишь при высоком его содержании в питательной среде. Азотфиксирующая способность культуры подавляется аммиаком (вообще содержание в среде связанного азота угнетает азотфиксацию). Стимулируют процесс фиксации азота соединения молибдена. Установлено, что при фиксации азота процесс его восстановления протекает на одном и том же синтезируемом азотобактером ферментном комплексе и лишь конечный продукт (аммиак) отделяется от фермента. Нитрогеназная азотфиксирующая система представляет собой мультиферментный комплекс, содержащий не связанное с геном железо, молибден и SH-группы. Микробиологическая промышленность выпускает несколько видов азотобактерина: сухой, почвенный и торфяной. Технология получения сухого азотобактерина имеет много общего с технологией производства сухого нитрагина. Сухой азотобактерин — активная культура высушенных клеток азотобактера с наполнителем. В 1 г препарата содержится не менее 0.5 млрд. жизнеспособных клеток. Культуру микроорганизма выращивают методом глубинного культивирования на среде, содержащей те же компоненты, что и при культивировании клеток Rhizobium. Дополнительно вводят только сульфаты железа и марганца, а также сложную соль молибденовой кислоты, pH 5.7–6.5. Процесс ферментации проводят до стационарной фазы развития культуры, так как в этой фазе биологически активные вещества выделяются из клетки и остаются в культуральной жидкости. Биологически активные вещества могут также полностью или частично теряться при высушивании, однако жизнеспособные клетки быстро восстанавливают способность их продуцировать. Высушенную культуру стандартизируют, фасуют в полиэтиленовые пакеты по 0.4–2 кг и хранят при температуре 15 °C не более 3 месяцев. Почвенный и торфяной азотобактерин представляют собой активную культуру азотобактера, размноженную на твердой питательной среде, и содержат в 1 г не менее 50 млн. жизнеспособных клеток. Для их приготовления берут плодородную почву или разлагающийся торф с нейтральной реакцией среды. К просеянному субстрату добавляют 2 % извести и 0.1 % суперфосфата. По 500 г полученной смеси переносят в бутыли емкостью по 0.5 л, увлажняют на 40–60 % по объему водой, закрывают ватными пробками и стерилизуют. Посевной материал готовят на агаровых средах, содержащих 2 % сахарозы и минеральные соли. Когда агар полностью покрывается слизистой массой коричневого цвета, полученный материал стерильно смывается дистиллированной водой и переносится на приготовленный субстрат. Содержимое бутылок тщательно перемешивают и термостатируют при 25–27 °C. Культивирование продолжают до тех пор, пока бактерии не размножатся до необходимого количества. Полученный препарат сохраняет свою активность в течение 2–3 месяцев. Использовать азотобактерин рекомендуется только на почвах, содержащих фосфор и микроэлементы. Азотобактерин применяют для бактеризации семян, рассады, компостов. При этом урожайность увеличивается на 10–15 %. Семена зерновых опудривают сухим азотобактерином из расчета 100 млрд. клеток на 1 гектарную порцию семян. Картофель и корневую систему рассады равномерно смачивают водной суспензией бактерий. Для получения суспензии 1 гектарную норму (300 млрд. клеток) разводят в 15 литрах воды. При обработке почвенным или торфяным азотобактерином семена перемешивают с увлажненным препаратом и для равномерного высева подсушивают. Корневую систему рассады смачивают приготовленной суспензией.
Технология получения фосфобактерина Фосфобактерин — бактериальное удобрение, содержащее споры микроорганизма Bacillus megaterium var. phosphaticum. Представляет собой порошок светло-серого или желтоватого цвета. Бактерии обладают способностью превращать сложные фосфорорганические соединения (нуклеиновые кислоты, нуклеопротеиды и т. д.) и трудноусвояемые минеральные фосфаты в доступную для растений форму. Кроме этого бактерии вырабатывают биологически активные вещества (тиамин, пиридоксин, биотин, пантотеновую и никотиновую кислоты и др.), стимулирующие рост растения. Фосфобактерин относится к числу препаратов со стимулирующим эффектом. Bacillus megaterium var. phosphaticum представляют собой мелкие, грамположительные аэробные спорообразующие палочки размером 2*6 мкм. Клетки содержат значительное количество соединений фосфора. В ранней стадии развития это подвижные одиночные палочки, при старении образуют эндоспоры, локализующиеся в одном из концов клетки. В силу вышеизложенного технология выращивания сводится к получению спор. В целом производство фосфобактерина похоже на производство азотобактерина и препаратов клубеньковых бактерий. Состав питательной среды в процентах: кукурузный экстракт — 1.8, меласса — 1.5, сульфат аммония — 0.1, мел — 1, остальное — вода. Культивирование ведется глубинным методом в строго асептических условиях при постоянном перемешивании и принудительной аэрации до стадии образования спор. Основные параметры проведения процесса: температура 28–30 °C, pH 6.5–7.5, длительность культивирования 1.5–2 суток. Полученную в ходе культивирования биомассу клеток отделяют центрифугированием и высушивают в распылительной сушилке при температуре 65–75 °C до остаточной влажности 2–3 %. Высушенные споры смешивают с наполнителем. Готовый препарат должен содержать не менее 8 млрд. клеток в 1 г. Расфасовывают препарат в полиэтиленовые пакеты по 50-500 г. В отличие от нитрагина и азотобактерина фосфобактерин обладает большей устойчивостью при хранении. Фосфобактерин рекомендуют применять на черноземных почвах, которые содержат наиболее значительное количество фосфороорганических соединений. Необходим для повышения урожайности зерновых, картофеля, сахарной свеклы и др. сельскохозяйственных растений. Семена обрабатывают смесью сухого фосфобактерина с наполнителем (золой, почвой и др.) в соотношении 1:40. На 1 гектарную порцию требуется 5 г препарата и 200 г наполнителя. Клубни картофеля равномерно увлажняют суспензией спор, приготовленной из расчета 15 г препарата на 15 л воды. Урожай при этом повышается на 10 %. Антибиотики для сельского хозяйства Антибиотики применяют в нескольких целях: — для борьбы с болезнями животных; — для борьбы с болезнями растений; — как стимуляторы роста животных; — при консервировании продуктов; — в научных исследованиях (в области биохимии, молекулярной биологии, генетике, онкологии). Современное определение термина[64] "антибиотик" принадлежит М.М.Шемякину и А.С.Хохлову (1961), которые предложили считать антибиотическими веществами все продукты обмена любых организмов, способные избирательно убивать или подавлять рост и развитие микроорганизмов. Полная химическая структура установлена только для трети антибиотиков, а может быть получена химическим путем лишь половина из них. Синтез микроорганизмами антибиотиков — одна из форм проявления антагонизма, связан с определенным характером обмена веществ, возникшим и закрепленным в ходе эволюции. Воздействуя на постороннюю микробную клетку, антибиотик вызывает нарушения в её развитии. Некоторые антибиотики способны подавлять синтез оболочки бактериальной клетки в период размножения, другие изменяют проницаемость цитоплазматической мембраны, некоторые ингибируют реакции обмена веществ. Механизм действия антибиотиков выявлен не полностью. В течение многих лет антибиотики используют как стимуляторы роста сельскохозяйственных животных и птицы, как средства борьбы с заболеваниями растений и посторонней микрофлорой в ряде бродильных производств, как консерванты пищевых продуктов. Механизм стимулирующего действия антибиотиков также не до конца выяснен. Предполагают, что стимулирующий эффект низких концентраций антибиотиков на организм животного связан с двумя факторами: — воздействие на микрофлору кишечника, — непосредственное влияние на организм животного. В первом случае антибиотики снижают число вредных и увеличивают количество полезных для организма микроорганизмов. Во втором случае — снижают pH содержимого кишечника, уменьшают поверхностное натяжение клеток организма, что способствует ускорению их деления. Кроме того, антибиотики увеличивают количество ростовых гормонов, приспособляемость организма к неблагоприятным условиям и т. д. Кормовые антибиотики применяют в виде неочищенных препаратов, представляющих собой высушенную массу продуцента, содержащую помимо антибиотика аминокислоты, ферменты, витамина группы В и другие биологически активные вещества. Все производимые кормовые антибиотики: а) не используются в терапевтических целях и не вызывают перекрестной резистентности бактерий к антибиотикам, применяемым в медицина; б) практически не всасываются в кровь из пищеварительного тракта; в) не меняют своей структуры в организме; г) не обладают антигенной природой, способствующей возникновению аллергии. В настоящее время выпускаются несколько видов кормовых антибиотиков: препараты на основе хлортетрациклина (биовит, кормовой биомицин), бацитрацин, гризин, гигромицин Б и др. Из этих препаратов только бацитрацин представляет собой высушенную культуральную жидкость, полученную в результате глубинного выращивания Bacilus licheniformis. Остальные антибиотики являются продуктами жизнедеятельности разных видов Actinomyces. Антибиотики используют и как средство борьбы с различными фитопатогенами. Воздействие антибиотика сводится к замедлению роста и гибели фитопатогенных микроорганизмов, содержащихся в семенах и вегетативных органах растений. К таким антибиотикам относятся фитобактериомицин, трихотецин, полимицин. Применение антибиотиков в пищевой промышленности позволяет снизить длительность термообработки продуктов питания при их консервировании. Используемые антибиотики воздействуют на клостридиальные и термофильные бактерии, устойчивые к нагреванию. Наиболее эффективным признан низин, который практически не токсичен для человека и позволяет вдвое снизить время термообработки. В заключение хотелось бы отметить, что в настоящем пособии не рассмотрены промышленные производства всех веществ, получаемых с помощью биотехнологии. Однако получить представление об основных принципах организации и осуществления биотехнологических процессов при производстве первичных и вторичных метаболитов, а также жизнеспособных микроорганизмов можно, думается, и из приведенного выше материала. Кроме того, рассмотренные примеры могут быть использованы в той или иной форме при изложении материала на уроках биологии.
ИММОБИЛИЗОВАННЫЕ ФЕРМЕНТЫ
Общая характеристика В современной биотехнологии одно из видных мест принадлежит ферментам. Ферменты и ферментные системы широко используются в различных отраслях промышленности, медицине, сельском хозяйстве, химическом анализе и т. д. Ферменты — вещества белковой природы и поэтому неустойчивы при хранении, а также чувствительны к тепловым воздействиям. Кроме того, ферменты не могут быть использованы многократно из-за трудностей в отделении их от реагентов и продуктов реакции. Решить эти проблемы помогает создание иммобилизованных ферментов. Начало этому методу было положено в 1916 году, когда Дж. Нельсон и Е.Гриффин адсорбировали на угле инвертазу и показали, что она сохраняет в таком виде каталитическую активность. Сам термин "иммобилизованные ферменты узаконен в 1971 году, и означает любое ограничение свободы передвижения белковых молекул в пространстве. Преимущества иммобилизованных ферментов перед нативными предшественниками: 1. Гетерогенный катализатор легко отделим от реакционной среды, что дает возможность остановить реакцию в любой момент, использовать фермент повторно, а также получать чистый от фермента продукт. 2. Ферментативный процесс с использованием иммобилизованных ферментов можно проводить непрерывно, регулируя скорость катализируемой реакции и выход продукта. 3. Модификация фермента целенаправленно изменяет его свойства, такие как специфичность (особенно в отношении макромолекулярного субстрата), зависимость каталитической активности от pH, ионного состава и других параметров среды, стабильность к денатурирующим воздействиям. 4. Можно регулировать каталитическую активность иммобилизованных ферментов путем изменения свойств носителя действием физических факторов, таких как свет и звук. Иммобилизовать ферменты можно как путем связывания на нерастворимых носителях, так и путем внутримолекулярной или межмолекулярной сшивки белковых молекул низкомолекулярными бифункциональными соединениями, а также путем присоединения к растворимому полимеру.
Классификация носителей Для получения иммобилизованных ферментов используется ограниченное число как органических, так и неорганических носителей. К носителям предъявляются следующие требования (Дж. Порат, 1974): — высокая химическая и биологическая стойкость; — высокая химическая прочность; — достаточная проницаемость для фермента и субстратов, пористость, большая удельная поверхность; — возможность получения в виде удобных в технологическом отношении форм (гранул, мембран); — легкая активация; — высокая гидрофильность; — невысокая стоимость. Классификация носителей схематично представлена на рисунке 4.

Рис. 4. Классификация носителей для иммобилизованных ферментов
Следует отметить, что органические носители (как низко-, так и высокомолекулярные) могут быть природного или синтетического происхождения. Природные полимерные органические носители делят в соответствии с их биохимической классификацией на 3 группы: полисахаридные, белковые и липидные. Синтетические полимеры также можно разделить на группы в связи с химическим строением основной цепи макромолекул: полиметиленовые, полиамидные, полиэфирные. Для иммобилизации ферментов наиболее широко используются природные полисахариды и синтетические носители полиметильного типа, остальные применяются значительно реже. Большое значение природных полимеров в качестве носителей для иммобилизации объясняется их доступностью и наличием реакционно-способных функциональных групп, легко вступающих в химические реакции. Характерной особенностью этой группы носителей также является их высокая гидрофильность. Недостаток природных полимеров — неустойчивость к воздействию микроорганизмов и довольно высокая стоимость. Наиболее часто для иммобилизации используются такие полисахариды, как целлюлоза, декстран, агароза и их производные. Целлюлоза гидрофильна, имеет много гидроксильных групп, что позволяет модифицировать её, замещая эти группы. Для увеличения механической прочности целлюлозу гранулируют путем частичного гидролиза, в результате которого разрушаются аморфные участки. На их место для сохранения пористости между кристаллическими участками вводят химические сшивки. Гранулированную целлюлозу довольно легко превратить в различные ионообменные производные, такие как ДЭАЭ-целлюлоза, КМЦ и т. д. Широко распространены носители на основе декстрана, выпускаемые под названием "сефадексы". При высушивании они легко сжимаются, в водном растворе сильно набухают. В этих носителях размер пор в геле регулируется степенью сшитости. К группе декстранов относят и крахмал. Химически модифицированный крахмал сшивается агентами, такими как формальдегид. Таким способом был получен губчатый крахмал, обладающий повышенной устойчивостью по отношению к ферментам, гидролизу. Водорастворимые препараты на основе декстрана часто применяются как носители лекарственных средств в медицине. Хорошим носителем считается агар. Его свойства улучшаются после химической сшивки, например, диэпоксидными соединениями. Такой агар становится устойчивым к нагреванию, прочен, легко модифицируется. Белки в качестве носителей обладают рядом достоинств: вместительны, способны к биодеградации, могут применяться в качестве тонкой (толщиной 80 мкм) мембраны. Иммобилизацию ферментов на белковых носителях можно проводить как в отсутствие, так и в присутствии сшивающих агентов. Белки используются и в фундаментальных биологических исследованиях, и в медицине. К недостаткам белков в качестве носителей относят их высокую иммуногенность (за исключением коллагена и фибрина). Наиболее для иммобилизации используются структурные (кератин, фибрин, коллаген), двигательные (миозин) и транспортные (альбумин) белки. Синтетические полимерные носители применяются для ковалентной и сорбционной иммобилизации ферментов, для получения гелей, микрокапсул. Полимеры на основе стирола применяются сорбционной иммобилизации. Они могут иметь макропористую, изопористую структуру, а также гетеропористую структуру. Для получения полимерных гидрофильных носителей широко используется акриламид — производное акриловой кислоты. Широкое распространение получил метод включения ферментов и клеток в полиакриламидный гель, имеющий жесткую пространственную сетчатую структуру. Полиакриламидный гель устойчив к химическим воздействиям. Очень интересную группу представляют полиамидные носители. Это группы различных гетероцепных полимеров с повторяющейся амидной группой — С(О)-NH-. Например, полимеры на основе N-винилпирролидона используются для получения иммобилизованных ферментов, способных медленно распадаться в организме. Кроме того, они биологически инертны, что особенно важно при использовании в медицинских целях. Существенным недостатком большинства полимерных носителей является их способность накапливаться в организме. В этом отношении предпочтение отдается природным полимерам, которые гидролизуются ферментами. Поэтому в состав лекарственных препаратов часто входит декстран, а из синтетических носителей — полимеры на основе N-винилпирролидона. В настоящее время ведутся эксперименты по созданию синтетических полимеров, расщепляющихся с образованием нетоксичных продуктов обмена.
Методы иммобилизации ферментов Существует два основных метода иммобилизации ферментов: физический и химический. Физическая иммобилизация ферментов представляет собой включение фермента в такую среду, в которой для него доступной является лишь ограниченная часть общего объема. При физической иммобилизации фермент не связан с носителем ковалентными связями. Существует четыре типа связывания ферментов: — адсорбция на нерастворимых носителях; — включение в поры геля; — пространственное отделение фермента от остального объема реакционной системы с помощью полупроницаемой перегородки (мембраны); — включение в двухфазную среду, где фермент растворим и может находиться только в одной из фаз. Перечисленные подходы проиллюстрированы рисунке 5.
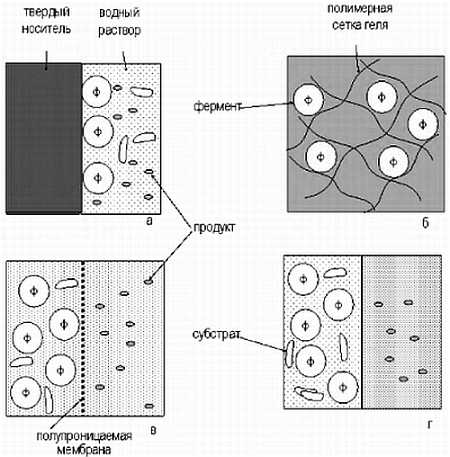
Рис. 5. Способы иммобилизации ферментов: а — адсорбция на нерастворимых носителях, б — включение в поры геля, в — отделение фермента с помощью полупроницаемой мембраны, г — использование двухфазной реакционной среды
Адсорбционная иммобилизация является наиболее старым из существующих способов иммобилизации ферментов, начало ей было положено еще в 1916 г. Этот способ достаточно прост и достигается при контакте водного раствора фермента с носителем. После отмывки неадсорбировавшегося белка иммобилизованный фермент готов к использованию. Удерживание адсорбированной молекулы фермента на поверхности носителя может обеспечиваться за счет неспецифических ван-дер-ваальсовых взаимодействий, водородных связей, электростатических и гидрофобных взаимодействий между носителем и поверхностными группами белка. Вклад каждого из типов связывания зависит от химической природы носителя и функциональных групп на поверхности молекулы фермента. Взаимодействия с носителем могут оказаться настолько сильными, что сорбция биокатализатора может сопровождаться разрушением его структуры. Например, при адсорбции некоторых растительных клеток на гранулах цитодекса клеточная стенка деформируется, повторяя рельеф поверхности частиц носителя. Преимуществом метода адсорбционной иммобилизации является доступность и дешевизна сорбентов, выступающих в роли носителей. Им также можно придать любую конфигурацию и обеспечить требуемую пористость. Важным фактор — простота применяемых методик. При адсорбционном связывании можно решить и проблему очистки фермента, так как связывание белка с носителем во многих случаях достаточно специфическое. К сожалению, прочность связывания фермента с носителем не всегда достаточно высока, что ограничивает применение метода. К недостаткам адсорбционной иммобилизации следует отнести отсутствие общих рекомендаций, позволяющих сделать правильный выбор носителя и оптимальных условий иммобилизации конкретного фермента. Некоторых из перечисленных затруднений можно избежать при иммобилизации ферментов путем включения в гели. Суть этого метода иммобилизации состоит в том, что молекулы фермента включаются в трехмерную сетку из тесно переплетенных полимерных цепей, образующих гель. Среднее расстояние между соседними цепями в геле меньше размера молекулы включенного фермента, поэтому он не может покинуть полимерную матрицу и выйти в окружающий раствор, т. е. находится в иммобилизованном состоянии. Дополнительный вклад в удерживание фермента в сетке геля могут вносить также ионные и водородные связи между молекулой фермента и окружающими ее полимерными цепями. Пространство между полимерными цепями в геле заполнено водой, на долю которой обычно приходится значительная часть всего объема геля. Например, широко применяемые гели полимеров акриловой кислоты в зависимости от концентрации полимера и его природы содержат от 50 до 90 % воды. Для иммобилизации ферментов в геле существует два основных способа. При одном из них фермент помещают в водный раствор мономера, а затем проводят полимеризацию, в результате чего образуется полимерный гель с включенными в него молекулами фермента. В реакционную смесь часто добавляют также бифункциональные (содержащие в молекуле две двойные связи) сшивающие агенты, которые придают образующемуся полимеру структуру трехмерной сетки. В другом случае фермент вносят в раствор готового полимера, который затем каким-либо образом переводят в гелеобразное состояние. Способ иммобилизации ферментов путем включения в полимерный гель позволяет создавать препараты любой геометрической конфигурации, обеспечивая при этом равномерное распределение биокатализатора в объеме носителя. Метод универсален, применим для иммобилизации практически любых ферментов, полиферментных систем, клеточных фрагментов и клеток. Фермент, включенный в гель, стабилен, надежно защищен от инактивации вследствие бактериального заражения, так как крупные клетки бактерий не могут проникнуть в мелкопористую полимерную матрицу. В то же время, эта матрица может создавать значительные препятствия для диффузии субстрата к ферменту, снижая каталитическую эффективность иммобилизованного препарата, поэтому для высокомолекулярных субстратов данный метод иммобилизации не применим вообще. Общий принцип иммобилизации ферментов с использованием мембран заключается в том, что водный раствор фермента отделяется от водного раствора субстрата полупроницаемой перегородкой. Полупроницаемая мембрана легко пропускает небольшие молекулы субстрата, но непреодолима для крупных молекул фермента. Существующие модификации этого метода различаются лишь способами получения полупроницаемой мембраны и ее природой. Водный раствор фермента можно включать внутрь микрокапсул, представляющих собой замкнутые сферические пузырьки с тонкой полимерной стенкой (микрокапсулирование). При двойном эмульгировании получается водная эмульсия из капель органического раствора полимера, содержащих, в свою очередь, еще более мелкие капли водного раствора фермента. Через некоторое время растворитель затвердевает, образуя сферические полимерные частицы с иммобилизованным в них ферментом. Если вместо водонерастворимого отвердевающего полимера используются жидкие углеводороды с высокой молекулярной массой, метод называется иммобилизацией путем включения в жидкие мембраны. К модификациям метода иммобилизации ферментов с использованием полупроницаемых оболочек относятся также включение в волокна (при этом вместо капель, содержащих ферменты, получаются нити) и включение в липосомы. Применение систем мембранного типа позволяет получать иммобилизованные препараты с высоким содержанием фермента. Метод, как и предыдущий, достаточно универсален, т. е. применим как ферментам, так и к клеткам, а также их фрагментам. Благодаря высокому отношению поверхности к объему и малой толщине мембраны удается избежать значительных диффузионных ограничений скорости ферментативных реакций. Основной недостаток мембранных систем — невозможность ферментативного превращения высокомолекулярных субстратов. При иммобилизации ферментов с использование систем двухфазного типа ограничение свободы перемещения фермента в объеме системы достигается благодаря его способности растворяться только в одной из фаз. Субстрат и продукт ферментативного превращения распределяются между обеими фазами в соответствии с их растворимостями в этих фазах. Природа фаз подбирается таким образом, что продукт накапливается в той из них, где фермент отсутствует. После завершения реакции эту фазу отделяют и извлекают из нее продукт, а фазу, содержащую фермент, вновь используют для проведения очередного процесса. Одним из важнейших преимуществ систем двухфазного типа является то, что они позволяют осуществлять ферментативные превращения макромолекулярных субстратов, которые невозможны при применении жестких носителей с ограниченным размером пор. Главным отличительным признаком химических методов иммобилизации является то, что путем химического взаимодействия на структуру фермента в его молекуле создаются новые ковалентные связи, в частности между белком и носителем. Препараты иммобилизованных ферментов, полученные с применением химических методов, обладают по крайней мере двумя важными достоинствами. Во-первых, ковалентная связь фермента с носителем обеспечивает высокую прочность образующегося конъюгата. При широком варьировании таких условий, как pH и температура, фермент не десорбируется с носителя и не загрязняет целевых продуктов катализируемой им реакции. Это особенно важно при реализации процессов медицинского и пищевого назначения, а также для обеспечения устойчивых, воспроизводимых результатов в аналитических системах. Во-вторых, химическая модификация фер ментов способна приводить к существенным изменениям их свойств, таких как субстратная специфичность, каталитическая активность и стабильность. Химическая иммобилизация ферментов является искусством, уровень которого определяется, в первую очередь, умением экспериментатора. Основная задача экспериментатора заключается в формировании новых ковалентных связей в молекуле фермента при использовании его функциональных групп, несущественных для проявления его каталитической активности. При химической модификации фермента его активный центр желательно защищать. При сопоставлении различных приемов иммобилизации химические методы для крупномасштабных биотехнологических процессов кажутся малопривлекательными из-за сложности и дороговизны. В промышленных процессах обычно используются те или иные методы физической иммобилизации.
Применение иммобилизованных ферментов Особенно ощутимый вклад иммобилизованные ферменты внесли в тонкий органический синтез, в анализ, в медицину, в процессы конверсии энергии, в пищевую и фармацевтическую промышленности. Для синтетической органической химии важно то, что в двухфазных реакционных средах фермент сохраняет каталитическую активность даже при исключительно малом содержании воды, поэтому равновесие катализируемой реакции (выход продукта) экспериментатор может регулировать в широких пределах, подбирая нужный органический растворитель. Иммобилизованные ферменты дали толчок к созданию принципиально новых методов "безреагентного" непрерывного анализа многокомпонентных систем органических (в ряде случаев и неорганических) соединений. В будущем важную роль в контроле окружающей среды и в клинической диагностике должны сыграть такие методы, как биолюминесцентный анализ и иммуноферментный анализ. В медицине иммобилизованные ферменты открыли путь к созданию лекарственных препаратов пролонгированного действия со сниженной токсичностью и аллергенностью. Иммобилизационные подходы способствуют решению проблемы направленного транспорта лекарств в организме. Проблемы биоконверсии массы и энергии в настоящее время пытаются решить микробиологическим путем. Тем не менее иммобилизованные ферменты вносят ощутимый вклад в осуществление фотолиза воды и в биоэлектрокатализ. Заслуживает внимание и использование иммобилизованных ферментов в процессах переработки лигноцеллюлозного сырья. Иммобилизованные ферменты могут использоваться и как усилители слабых сигналов. На активный центр иммобилизованного фермента можно подействовать через носитель, подвергая последний ультразвуковой обработке, механическим нагрузкам или фотохимическим превращениям. Это позволяет регулировать каталитическую активность системы фермент — носитель под действием механических, ультразвуковых и световых сигналов. На этой основе были созданы механо- и звукочувствительные датчики и открыт путь к бессеребряной фотографии. Промышленные процессы с применением иммобилизованных ферментов внедрены прежде всего в пищевую и фармацевтическую промышленность. В пищевой промышленности с участием иммобилизованных ферментов идут процессы получения глюкозо-фруктовых сиропов, глюкозы, яблочной и аспарагиновой кислоты, оптически активных L-аминокислот, диетического безлактозного молока, сахаров из молочной сыворотки и др. В медицине иммобилизованные ферменты используются также как лекарственные препараты, особенно в тех случаях, когда необходимо локальное воздействие. Кроме того, биокатализаторы широко используются в различных аппаратах для перфузионной очистки различных биологическихжидкостей. Возможности и перспективы использования в медицине ферментов в иммобилизованном состоянии гораздо шире, чем достигнутые на сегодняшний день, именно на этом пути медицину ждет создание новых высокоэффективных методов лечения.
ИММОБИЛИЗОВАННЫЕ КЛЕТКИ МИКРООРГАНИЗМОВ
В 70-х годах XX века появились первые публикации об иммобилизации клеток микроорганизмов, а первое промышленное применение иммобилизованных клеток было осуществлено в Японии в 1974 г. С их помощью получали аспарагиновую кисло- Иммобилизованные клетки имеют ряд преимуществ как перед иммобилизованными ферментами, так и перед свободными клетками: — отсутствие затрат на выделение и очистку ферментов; — снижение затрат на выделение и очистку продуктов реакции; — более высокая активность и стабильность; — возможность создания непрерывных и полунепрерывных автоматизированных процессов; — способность к длительному функционированию полиферментных систем без экзогенных кофакторов. Для иммобилизации могут быть использованы клетки в различном состоянии: живые и поврежденные в различной степени. Одностадийные реакции могут осуществлять и живые, и поврежденные клетки. Полиферментные реакции проводят с применением живых клеток, которые могут длительное время регенерировать АТФ и коферменты (НАДФ, НАД). Проблема использования ферментативной активности иммобилизованных микроорганизмов имеет глубокие корни. Более 150 лет назад быстрый способ получения уксуса был основан на применении микроорганизмов, адсорбированных на древесной стружке. Методы иммобилизации клеток схожи с методами иммобилизации ферментов. Химический метод основан на образовании ковалентных связей с активированным носителем, на поперечной сшивке клеток за счет активных групп в клеточной оболочке с бифункциональными реагентами (например, глутаровым альдегидом) К физическим методам относятся адсорбция и агрегация. Иммобилизация клеток путем включения в различные гели, мембраны, волокна основана на химических и физических взаимодействиях. Химические методы используются реже по сравнению с другими методами и малопригодны для иммобилизации живых клеток. Гораздо большее распространение получило включение клеток в состав гелей, мембран и волокон. При таком способе иммобилизации клетки могут сохранять жизнеспособность и в присутствии питательной среды размножаться в приповерхностных слоях гелей. Биокаталитическая активность целых иммобилизованных клеток в настоящее время может быть использована в различных отраслях науки и техники: — при биосинтезе и трансформации таких соединений, как аминокислоты, органические кислоты, антибиотики, стероиды углеводы, углеводороды, нуклеотиды и нуклеозиды; — в пивоварении и виноделии; — при очистке сточных и природных вод; — при извлечении металлов из сточных вод; — при ассимиляции солнечной энергии; — при изготовлении водородных солнечных элементов; — в азотфиксации; — в аналитических целях при изготовлении электродов. Наибольшее количество исследований по иммобилизации клеток микроорганизмов проведено японскими исследователями. Особые успехи были достигнуты ими в области синтеза аминокислот, органических кислот и антибиотиков. В Московском государственном университете был разработан метод получения аспарагиновой кислоты, который по эфективности не уступает японским. Клетки E.coli, включенные в армированный полиакриламидный гель, были с успехом использованы для получения аспарагиновой кислоты, период полужизни катализатора — 110 суток. Иммобилизовать можно не только клетки микроорганизмов, но и клетки растительных и животных тканей, используя их для синтеза физиологически активных соединений. Интересные возможность открываются и при иммобилизации клеточных органелл как активных полиферментных систем. Все это свидетельствует о перспективности развития одного из направлений биотехнологии, связанного с изучением и применением иммобилизованных клеток.
БИОТЕХНОЛОГИЯ И ЭКОЛОГИЧЕСКИЕ ПРОБЛЕМЫ
Биодеградация ксенобиотиков В удалении ксенобиотиков из окружающей среды важны несколько факторов: — устойчивость ксенобиотиков к различным воздействиям; — растворимость их в воде; — летучесть ксенобиотиков; — pH среды; — способность ксенобиотиков поступать в клетки микроорганизмов; — сходство ксенобиотиков и природных соединений, подвергающихся естественной биодеградации. Для биодеградации ксенобиотиков лучше использовать ассоциации микроорганизмов, так как они более эффективны, чем отдельно взятые виды. При этом типы связей в подобной ассоциации могут быть различны. Один вид микроорганизмов может непосредственно участвовать в разложении ксенобиотиков, а другой — поставлять недостающие питательные вещества. Это может быть метаболическая «атака» на субстрат, когда синтезируются разные компоненты ферментативного комплекса, или же цепочка ферментативных реакций (многосубстратные конверсии) и т. д. Особенно трудно разлагаются такие биоциды, как детергенты, пластики и углеводороды. Самыми способными к борьбе с загрязнителями различного типа являются представители рода Pseudomonas — они практически «всеядны». Клетки этих микроорганизмов содержат оксидоредуктазы и гидроксилазы, способные разлагать большое число молекул углеводородов и ароматических соединений, таких как бензол, ксилол, толуол. Гены, кодирующие эти ферменты, находятся в составе плазмид. Например, плазмида ОСТ отвечает за разложение октана и гексана, XYL — ксилола и толуола, NAH — нафталина, САМ — камфары. Плазмиды САМ и NAH обеспечивают собственный перенос, индуцируя скрещивание бактериальных клеток; остальные плазмиды могут быть перенесены только в том случае, если в бактерии введены другие плазмиды, обеспечивающие скрещивание. В 1979 г. Чакрабарти (в то время совместно с компанией «Дженерал электрик») после успешных скрещиваний получил штамм, содержащий плазмиды XYL и NAH, а также гибридную плазмиду, полученную путем рекомбинации частей плазмид САМ и ОСТ (сами по себе они несовместимы, т. е. не могут сосуществовать как отдельные плазмиды в одной бактериальной клетке). Этот штамм способен быстро расти на неочищенной нефти, так как он метаболизирует углеводороды гораздо активнее, чем любой из штаммов, содержащих только одну плазмиду. Штамм может быть особенно полезен в очистных водоемах для сточных вод, где можно контролировать температуру и другие внешние факторы. Эти микроорганизмы удобно использовать для очистки нефтяных пятен на суше или море при различных авариях. Для большей эффективности создают микроэмульсию, содержащую бактериальные штаммы и капсулы со смесью основных питательных элементов — азота, фосфора и калия внутри. Добавление этих веществ стимулирует размножение бактериальных штаммов. Применение такого метода позволяет очистить от 70 до 90 % загрязненной поверхности, за это же время очищается всего порядка 10–20 % необработанной поверхности. Преимущество бактериальной очистки по сравнению с химической в том, что она не вызывает появления нового загрязняющего агента в окружающей среде. Плотность фитопланктона после бактериальной очистки повышается. Некоторые микроорганизмы способны изменять молекулу ксенобиотика и делать ее доступной и привлекательной для других микроорганизмов («кометаболизм»). Примером может служить разложение инсектицида паратиона под действием двух штаммов Pseudomonas — P. aeruginosa и P. stuzeri. В некоторых случаях происходит неполное превращение молекулы ксенобиотика — фосфорилирование, метилирование, ацетилирование и т. д., результатом которого является утрата этим веществом токсичности. Одним из сильных загрязнителей является ЭДТА (этилендиаминтетрауксусная кислота). Причина в том, что ЭДТА связывает тяжелые металлы, способствуя их накоплению в почве. Бактрии родов Pseudomonas и Bacillus способны за две недели разрушить все связи комплекса Fe-ЭДТА. Эти бактерии успешно применяются для очистки бытовых сточных вод, куда попадают детергенты моющих средств. Кроме Pseudomonas, биодеградацию ксенобиотиков могут осуществлять и представители родов Acinetobacter, Metviosinus. Однако в некоторых случаях внесение этих микроорганизмов в почву может изменить экосистему местности. Избежать этого можно ограничивая время жизнедеятельности бактерий. Например, облучая штаммы ультрафиолетом, получили мутант, ауксотрофный по лейцину. Бактерии размножают в питательной среде, содержащей лейцин. Суспензией микроорганизмов в питательной среде пропитывают древесную стружку, которую разбрасывают по загрязненной территории. Количество лейцина рассчитывается на время, достаточное для уничтожения вредных примесей, поэтому после очистки мутантные штаммы гибнут. Еще эффективнее, чем бактерии, справляются с почвенными загрязнителями грибы. Они могут разрушать такие вещества, как пентахлорбензол, пентахлофенол. В одном из экспериментов грибами обработали около 10000 тонн почвы с территории деревоперерабатывающего комплекса. В этой почве содержание пентахлорфенола достигало 700 мг/кг, но за год деятельности оно снизилось до 10 мг/кг, что является допустимой нормой. Бактерии смогли бы переработать эту почву лишь за 4–5 лет. Грибы активны и зимой, разрушают высокомолекулярные полиароматические углеводороды, действуют внеклеточно, выделяя неспецифические ферменты. Стоимость грибной и бактериальной очистки одинаковы, но применение грибов позволяет сокращать сроки деградации и существенно удешевляет ее.
Аэробные системы очистки Биологическая переработка отходов опирается на ряд дисциплин: биохимию, генетику, химию, микробиологию, вычислительную технику. Усилия этих дисциплин концентрируются на трех основных направлениях: — деградация органических и неорганических токсичных отходов; — возобновление ресурсов для возврата в круговорот веществ углерода, азота, фосфора, азота и серы; — получение ценных видов органического топлива. При очистке сточных вод выполняют четыре основные операции: 1. При первичной переработке происходит усреднение и осветление сточных вод от механических примесей (усреднители, песколовки, решетки, отстойники). 2. На втором этапе происходит разрушение растворенных органических веществ при участии аэробных микроорганизмов. Образующийся ил, состоящий главным образом из микробных клеток, либо удаляется, либо перекачивается в реактор. При технологии, использующей активный ил, часть его возвращается в аэрационный тенк. 3. На третьем (необязательном) этапе производится химическое осаждение и разделение азота и фосфора. 4. Для переработки ила, образующегося на первом и втором этапах, обычно используется процесс анаэробного разложения. При этом уменьшается объем осадка и количество патогенов, устраняется запах и образуется ценное органическое топливо — метан. На практике применяются одноступенчатые и многоступенчатые системы очистки. Одноступенчатая схема очистки сточной воды представлена на рис. 6.
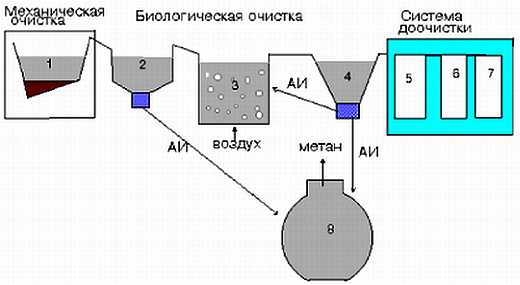
Рис. 6. Принципиальная схема очистных сооружений: 1 — пескоуловители; 2 — первичные отстойники; 3 — аэротенк; 4 — вторичные отстойники; 5 — биологические пруды; 6 — осветление; 7 — реагентная обработка; 8 — метатенк; АИ — активный ил
Сточные воды поступают в усреднитель, где происходит интенсивное перемешивание стоков с различным качественным и количественным составом. Перемешивание осуществляется за счет подачи воздуха. В случае необходимости в усреднитель подаются также биогенные элементы в необходимых количествах и аммиачная вода для создания определенного значения pH. Время пребывания в усреднителе составляет обычно несколько часов. При очистке фекальных стоков и отходов нефтепереработки необходимым элементом очистных сооружений является система механической очистки — песколовки и первичные отстойники. В них происходит отделение очищаемой воды от грубых взвесей и нефтепродуктов, образующих пленку на поверхности воды. Биологическая очистка воды происходит в аэротенках. Аэротенк представляет собой открытое железобетонное сооружение, через которое проходит сточная вода, содержащая органические загрязнения и активный ил. Суспензия ила в сточ ной воде на протяжении всего времени нахождения в аэротенке подвергается аэрации воздухом. Интенсивная аэрация суспензии активного ила кислородом приводит к восстановлению его способности сорбировать органические примеси. В основе биологической очистки воды лежит деятельность активного ила (АИ) или биопленки, естественно возникшего биоценоза, формирующегося на каждом конкретном производстве в зависимости от состава сточных вод и выбранного режима очистки. Активный ил представляет собой темно-коричневые хлопья, размером до нескольких сотен микрометров. На 70 % он состоит из живых организмов и на 30 % — из твердых частиц неорганической природы. Живые организмы вместе с твердым носителем образуют зооглей — симбиоз популяций микроорганизмов, покрытый общей слизистой оболочкой. Микрооганизмы, выделенные из активного ила относятся к различным родам: Actynomyces, Azotobacter, Bacillus, Bacterium, Corynebacterium, Desulfomonas, Pseudomonas, Sarcina и др. Наиболее многочисленны бактерии рода Pseudomonas, о всеядности которых упоминалось ранее. В зависимости от внешней среды, которой в данном случае является сточная вода, та или иная группа бактерий может оказаться преобладающей, а остальные становятся спутниками основной группы. Существенная роль в создании и функционировании активного ила принадлежит простейшим. Функции простейших достаточно многообразны; они сами не принимают непосредственного участия в потреблении органических веществ, но регулируют возрастной и видовой состав микроорганизмов в активном иле, поддерживая его на определенном уровне. Поглощая большое количество бактерий, простейшие способствуют выходу бактериальных экзоферментов, концентрирующихся в слизистой оболочке и тем самым принимать участие в деструкции загрязнений. В активных илах встречаются представители четырех классов простейших: саркодовые (Sarcodina), жгутиковые инфузории (Mastigophora), реснитчатые инфузории (Ciliata), сосущие инфузории (Suctoria). Показателем качества активного ила является коэффициент протозойности, который отражает соотношение количества клеток простейших микроорганизмов к количеству бактериальных клеток. В высококачественном иле на 1 миллион бактериальных клеток должно приходиться 10–15 клеток простейших. При изменении состава сточной воды может увеличится численность одного из видов микроорганизмов, но другие культуры все равно остаются в составе биоценоза. На формирование ценозов активного ила могут оказывать влияние и сезонные колебания температуры, обеспеченность кислородом, присутствие минеральных компонентов. Все это делает состав или сложным и практически невоспроизводимым. Эффективность работы очистных сооружений зависит также от концентрации микроорганизмов в сточных водах и возраста активного ила. В обычных аэротенках текущая концентрация активного ила не превышает 2–4 г/л. Увеличение концентрации ила в сточной воде приводит к росту скорости очистки, но требует усиления аэрации, для поддержания концентрации кислорода на необходимом уровне. Таким образом, аэробная переработка стоков включает в себя следующие стадии: 1) адсорбция субстрата на клеточной поверхности; 2) расщепление адсорбированного субстрата внеклеточными ферментами; 3) поглощение растворенных веществ клетками; 4) рост и эндогенное дыхание; 5) высвобождение экскретируемых продуктов; 6) "выедание" первичной популяции организмов вторичными потребителями. В идеале это должно приводить к полной минерализации отходов до простых солей, газов и воды. На практике очищенная вода и активный ил из аэротенка подаются во вторичный отстойник, где происходит отделение активного ила от воды. Часть активного ила возвращается в систему очистки, а избыток активного ила, образовавшийся в результате роста микроорганизмов, поступает на иловые площадки, где обезвоживается и вывозится на поля. Избыток активного ила можно также перерабатывать анаэробным путем. Переработанный активный ил может служить и как удобрения, и как корм для рыб, скота. Система полной доочистки может состоять из множества элементов, которые определяются дальнейшим назначением сточной воды. Возможно применение биологических прудов, где биологически очищенная вода проходит осветление и насыщается кислородом. Пруды также относятся к системе биологической очистки, в которой под воздействием биоценоза активного ила происходит окисление органических примесей. Состав биоценозов биологических прудов определяется глубиной нахождения данной группы микроорганизмов. В верхних слоях развиваются аэробные культуры, в придонных — факультативные аэробы и анаэробы, способные осуществлять процессы метанового брожения или восстановление сульфатов. Насыщение воды кислородом происходит за счет процессов фотосинтеза, осуществляемого водорослями, из которых особенно широко представлены Clorella, Scenedesmus, встречаются эвгленовые, вольвоксовые и т. д. В прудах также в той или иной мере представлена микро- и макрофауна: простейшие, черви, коловратки, насекомые и др. В биопрудах из воды хорошо удаляются нефтепродукты, фенолы и другие органические соединения. В некоторых случаях воду после биологической очистки подвергают реагентной обработке — хлорированию или озонированию. Интенсифицировать процессы биологической очистки можно путем аэрации суспензии активного ила чистым кислородом. Этот процесс можно осуществить в модифицированных аэротенках закрытого типа — окситенках, с принудительной аэрацией сточной воды. В отличие от аэротенков в биофильтрах (или перколяционных фильтрах) клетки микроорганизмов находятся в неподвижном состоянии, так как прикреплены к поверхности пористого носителя. Образовавшуюся таким образом биопленку можно отнести к иммобилизованным клеткам. В этом случае иммобилизована не монокультура, а целый консорциум, неповторимый по качественному и количественному составу и различающийся в зависимости от его местонахождения на поверхности носителя. Очищаемая вода контактирует с неподвижным носителем, на котором иммобилизованы клетки и за счет их жизнедеятельности происходит снижение концентрации загрязнителя. Преимущество применения биофильтров состоит в том, что формирование конкретного ценоза приводит к практически полному удалению всех органических примесей. Недостатками этого метода можно считать: — нереальность использования стоков с высоким содержанием органических примесей; — необходимость равномерного орошения поверхности биофильтра сточными водами, подаваемыми с постоянной скоростью; — сточные воды перед подачей должны быть освобождены от взвешенных частиц во избежание заиливания. В качестве носителей можно использовать керамику, щебень, гравий, керамзит, металлический или полимерный материал с высокой пористостью. Для биофильтров характерно наличие противотока воды, которая поступает сверху и воздуха, подающегося снизу. Оторвавшиеся частицы микробной пленки после отделения их во вторичном отстойнике не возвращаются обратно в биофильтр, а идут на иловые площадки или в анаэробную преработку. Существуют также системы, сочетающие в себе как систему биофильтров, так и активного ила в аэротенках. Это так называемые аэротенки-вытеснители. В аэрируемую сточную воду помещают либо стеклоерши, либо создают систему сеток внутри тенка, в которые вкладываются прокладки из пористого полиэфира. В пустотах этих прокладок и на поверхности стеклоершей происходит накопление биоценоза активного ила. Носитель периодически удаляется из тенка, биомасса снимается, после чего носитель возвращается в реактор. Система с иммобилизованными на мобильном носителе клетками отличается от биофильтров своей экономичностью, так как используются высокие концентрации микроорганизмов и нет необходимости осаждать конечные продукты. Такая система может найти применение в очистке локальных стоков, с узким спектром загрязнений. Их целесообразно очищать в самостоятельных биологических системах, не смешивая со стоками других производств. Это позволяет получить биоценозы микроорганизмов, адаптированные к данному узкому спектру загрязнений, при этом скорость и эффективность очистки резко возрастают.
Анаэробные системы очистки Как уже упоминалось, избыток активного ила может перерабатываться двумя способами: после высушивания как удобрение или же попадает в систему анаэробной очистки. Такие же способы очистки применяют и при сбраживании высококонцентрированных стоков, содержащих большое количество органических веществ. Процессы брожения осуществляются в специальных аппаратах — метатенках. Распад органических веществ состоит из трех этапов: — растворение и гидролиз органических соединений; — ацидогенез; — метаногенез. На первом этапе сложные органические вещества превращаются в масляную, пропионовую и молочную кислоты. На втором этапе эти органические кислоты превращаются в уксусную кислоту, водород, углекислый газ. На третьем этапе метанообразующие бактерии восстанавливают диокись углерода в метан с поглощением водорода. По видовому составу биоценоз метатенков значительно беднее аэробных биоценозов. Насчитывают около 50 видов микроорганизмов, способных осуществлять первую стадию — стадию кислотообразования. Самые многочисленные среди них — представители бацилл и псевдомонад. Метанообразующие бактерии имеют разнообразную форму: кокки, сарцины и палочки. Этапы анаэробного брожения идут одновременно, а процессы кислотообразования и метанообразования протекают параллельно. Уксуснокислые и метанообразующие микроорганизмы образуют симбиоз, считавшийся ранее одним микроорганизмом под названием Methanobacillus omelianskii. Процесс метанообразования — источник энергии для этих бактерий, так как метановое брожение представляет собой один из видов анаэробного дыхания, в ходе которого электроны с органических веществ переносятся на углекислый газ, который восстанавливается до метана. В результате жизнедеятельности биоценоза метатенка происходит снижение концентрации органических веществ и образование биогаза, являющегося экологически чистым топливом. Для получения биогаза мо гут использоваться отходы сельского хозяйства, стоки перерабатывающих предприятий, содержащих сахар, бытовые отходы, сточные воды городов, спиртовых заводов и т. д. Метатенк представляет собой герметичный ферментер объемом в несколько кубических метров с перемешиванием, который обязательно оборудуется газоотделителями с противопламенными ловушками. Метатенки работают в периодическом режиме загрузки отходов или сточных вод с постоянным отбором биогаза и выгрузкой твердого осадка после завершения процесса. В целом, активное использование метаногенеза при сбраживании органических отходов — один из перспективных путей совместного решения энергетических и экологических проблем, который позволяет агропромышленным комплексам перейти на автономное энергообеспечение.
Показатели загрязненности сточных вод На всех этапах очистки сточных вод ведется строгий контроль за качественным составом воды. При этом проводится детальный анализ состава сточной воды с выяснением не только концентраций тех или иных соединений, но и более полное определение качественного и количественного состава загрязнителей. Необходимость такого анализа определяется спецификой системы переработки, так как в сточных водах могут присутствовать токсические вещества, способные привести к гибели микроорганизмов и вывести систему из строя. Определение таких показателей, как органолептические (цвет, вид, запах, прозрачность, мутность), оптическая плотность, pH, температура не вызывает трудностей. Сложнее определить содержание органических веществ в сточной воде, которое необходимо знать для контроля работы очистных сооружений, повторного использования сточных вод в технологических процессах, выбора метода очистки и доочистки, окончания процесса очистки, а также оценки возможности сброса воды в водоемы. При определении содержания органических веществ широко используются два способа: химическое потребление кислорода и биохимическое потребление кислорода. В первом случае методика основана на окислении веществ, присутствующих в сточных водах, 0,25 % раствором дихромата калия при кипячении пробы в течение 2 часов в 50 % (по объему) растворе серной кислоты. Для полноты окисления органических веществ используется катализатор — сульфат серебра. Дихроматный способ достаточно прост и легко автоматизируется, что обуславливает его широкое распространение. Биохимическое потребление кислорода измеряется количеством кислорода, расходуемым микроорганизмами при аэробном биологическом разложении веществ, содержащихся в сточных водах при стандартных условиях за определенный интервал времени. Определение биохимического потребления кислорода требует специальной аппаратуры. В герметичный ферментер помещается определенное количество исследуемой сточной воды, которую засевают микроорганизмами. В процессе культивирования регистрируется изменение количества кислорода, пошедшего на окисление соединения, присутствующего в сточных водах. Лучше всего культивировать микроорганизмы из уже работающих биологических систем, адаптированных к данному спектру загрязнений. Определение лишь одного из показателей качества сточной воды (химического или биохимического потребления кислорода) не всегда позволяет оценить как ее доступность для биологической очистки, так и степень конечной очистки. Так, например, имеется целые группы соединений, определение химического потребления кислорода для которых невозможно, хотя эти соединения вполне доступны для биохимического определения кислорода и наоборот. Все это говорит о том, что для оценки чистоты сточных воды необходимо использовать одновременно оба метода. Биотехнология будет оказывать многообразное и все возрастающее влияние на способы контроля за окружающей средой и на ее состояние. Хорошим примером такого рода служит создание новых, более совершенных способов переработки отходов, однако применение биотехнологии в данной сфере отнюдь не ограничивается этим. Биотехнология будет играть все большую роль в химической промышленности и сельском хозяйстве, помогая создать замкнутые и полузамкнутые технологические циклы, решая хотя бы отчасти существующие здесь проблемы.
ЛИТЕРАТУРА К РАЗДЕЛУ «ПРОМЫШЛЕННАЯ БИОТЕХНОЛОГИЯ"
1. Альберте Б., Брей Д., Льюис Дж. и др. Молекулярная биология клетки. Т.1. М.: Мир, 1994. 2. Артамонов В.И. Занимательная физиология растений. М.: Агропромиздат, 1991. 336 с. 3. Безбородов А.М. Ферменты микроорганизмов и их применение // Биотехнология. М.: Наука, 1984. 4. Березин И.В., Клесов А.А., Швядас В.К. и др. Инженерная энзимология. М.: Высшая школа, 1987. 144 с. 5. Березин И.В., Клячко Н.Л., Левашев А.В. и др. Иммобилизованные ферменты. М.: Высшая школа, 1987. 160 с. 6. Биология наших дней. Вып. 2. -М.: Знание, 1987. 160 с. 7. Биотехнология. Принципы и применение /Хиггинс И., Бест Д., Джонс Дж. М.: Мир, 1988. 480 с. 8. Биотехнология сельскохозяйственных растений. М.: Агропромиздат, 1987. 301 с. 9. Биотехнология — сельскому хозяйству /Лобанок А.Г., Залашко М.В., Анисимова Н.И. и др. Минск: Урожай, 1988. 199 с. 10. Биотехнология растений: культура клеток. М.: Агропромиздат, 1989. 280 11. Быков В.А., Крылов И.А., Манаков М.Н. и др. Микробиологическое производство биологически активных веществ и препаратов. М.: Высшая школа, 1987. 142 с. 12. Быков В.А., Манаков М.Н., Панфилов В.И. и др. Производство белковых веществ. М.: Высшая школа, 1987. 142 с. 13. Варфоломеев С.Д., Панцхава Е.С. Биотехнология преобразования солнечной энергии. Современное состояние, проблемы, перспективы // Биотехнология. М.: Наука, 1984. 14. Голубовская Э.К. Биологические основы очистки воды. М.: Высшая школа, 1978. 270 с. 15. Грачева И.М., Гаврилова Н.М., Иванова Л.A. Технология микробных белковых препаратов, аминокислот и жиров. М.: Пищевая промышленность, 1980. 448 с. 16. Егоров Н.С. Основы учения об антибиотиках. М.: Высшая школа, 1986. 448 с. 17. Иммобилизованные клетки и ферменты. Методы / Под ред. Дж. Вудворта. М.: Мир, 1988. 215 с. 18. Казанская Н.Ф., Ларионова Н.И., Торчилин В.П. Ферменты и белковые препараты в медицине // Биотехнология. М.: Наука, 1984. 19. Каравайко Г.И. Биогеотехнология металлов // Биотехнология. М.: Наука, 1984. 20. Кефели В.И., Дмитриева Г.А. Биотехнология: курс лекций. Пущино, 1989. 96 с. 21. Клесов А.А. Применение иммобилизованных ферментов в пищевой промышленности/ /Биотехнология. М.: Наука, 1984. 22. Мартинек К. Иммобилизованные ферменты // Биотехнология. М.: Наука, 1984. 23. Методы культивирования клеток. Л.: Наука, 1988. 313 с. 24. Печуркин Н.С., Брильков А.В., Марченкова Т.В. Популяционные аспекты биотехнологии. Новосибирск: Наука, 1990. 173 с. 25. Пирузян Л.А., Михайловский Е.М. Сапротрофная микрофлора в качестве продуцента биологически активных веществ для целей микробной сапротрофной фармакотерапии // Изв. АН Серия биологическая, 1992. № 6. С. 860–866. 26. Реннеберг Р., Реннеберг И. От пекарни до биофабрики. М.: Мир, 1991. 112 с. 27. Рычков Р.С., Попов В.Г. Биотехнология перспективы развития // Биотехнология. М.: Наука, 1984. 28. Сассон А. Биотехнология: свершения и надежды. М.: Мир, 1987. 411 с. 29. Скрябин Г.К., Кощеенко К.А. Иммобилизованные клетки микроорганизмов // Биотехнология. М.: Наука, 1984. 30. Тривен М. Иммобилизованные ферменты. М.: Мир, 1983. 213 с. 27. 31. Хотянович А.В. Методы культивирования азотфиксирующих бактерий, способы получения и применение препаратов на их основе (методические рекомендации). Л., 1991. 60 с. 32. Шлегель Г. Общая микробиология. М.: Мир, 1987. 566 с.
Культуры растительных клеток[65]
КУЛЬТУРЫ КЛЕТОК ВЫСШИХ РАСТЕНИЙ
Использование культуры растительных клеток Культуры клеток высших растений имеют две сферы применения: 1. Изучение биологии клетки, существующей вне организма, обуславливает ведущую роль клеточных культур в фундаментальных исследованиях по генетике и физиологии, молекулярной биологии и цитологии растений. Популяциям растительных клеток присущи специфические особенности: генетические, эпигенетические (зависящие от дифференцированной активности генов) и физиологические. При длительном культивировании гетерогенной по этим признакам популяции идет размножение клеток, фенотип и генотип которых соответствуют данным условиям выращивания, следовательно, популяция эволюционирует. Все это позволяет считать, что культуры клеток являются новой экспериментально созданной биологической системой, особенности которой пока мало изучены. Культуры клеток и тканей могут служить адекватной моделью при изучении метаболизма и его регуляции в клетках и тканях целого растения. 2. Культивируемые клетки высших растений могут рассматриваться как типичные микрообъекты, достаточно простые в культуре, что позволяет применять к ним не только аппаратуру и технологию, но и логику экспериментов, принятых в микробиологии. Вместе с тем, культивируемые клетки способны перейти к программе развития, при которой из культивируемой соматической клетки возникает целое растение, способное к росту и размножению. Можно назвать несколько направлений создания новых технологий на основе культивируемых тканей и клеток растений. 1. Получение биологически активных веществ растительного происхождения: — традиционных продуктов вторичного метаболизма (токсинов, гербицидов, регуляторов роста, алкалоидов, стероидов, терпеноидов, имеющих медицинское применение); — синтез новых необычных соединений, что возможно благодаря исходной неоднородности клеточной популяции, генетической изменчивости культивируемых клеток и селективному отбору клеточных линий со стойкими модификациями, а в некоторых случаях и направленному мутагенезу; — культивируемые в суспензии клетки могут применятся как мультиферментные системы, способные к широкому спектру биотрансформаций химических веществ (реакции окисления, восстановления, гидроксилирования, метилирования, деметилирования, гликолизирования, изомеризации). В результате биотрансформации получают уникальные биологически активные продукты на основе синтетических соединений или веществ промежуточного обмена растений других видов. 2. Ускоренное клональное микроразмножение растений, позволяющее из одного экпланта получать от 10000 до 1000000 растений в год, причем все они будут генетически идентичны. 3. Получение безвирусных растений. 4. Эмбриокультура и оплодотворение in vitro часто применяются для преодоления постгамной несовместимости или щуплости зародыша, для получения растений после отдаленной гибридизации. При этом оплодотворенная яйцеклетка вырезается из завязи с небольшой частью ткани перикарпа и помещается на питательную среду. В таких культурах можно также наблюдать стадии развития зародыша. 5. Антерные культуры — культуры пыльников и пыльцы используются для получения гаплоидов и дигаплоидов. 6. Клеточный мутагенез и селекция. Тканевые культуры могут производить регенеранты, фенотипически и генотипически отличающиеся от исходного материала в результате сомаклонального варьирования. При этом в некоторых случаях можно обойтись без мутагенной обработки. 7. Криоконсервация и другие методы сохранения генофонда. 8. Иммобилизация растительных клеток. 9. Соматическая гибридизация на основе слияния растительных протопластов. 10. Конструирование клеток путем введения различных клеточных оганелл. 11. Генетическая трансформация на хромосомном и генном уровнях. 12. Изучение системы «хозяин — паразит» с использованием вирусов, бактерий, грибов и насекомых).
История метода Самые ранние работы по изолированию культур принадлежат Блоцишевскому (1876), Брауну и Моррису (1892), Боннэ, Саксу (1893). В этих исследованиях зародыши вычленялись из семени и выращивались в искусственных условиях. Первым исследователем, занявшимся установлением минимального размера экспланта, был Карл Рехингер (1893). Он выращивал тонкие срезы корня свеклы и одуванчика и сегменты стебля тополя на песке с применением водопроводной воды, без стерильных условий. Эти исследования показали, что каллус образуется при толщине среза не менее 1,5 мм. Еще в 19 веке X. Фёхтинг провел ряд экспериментов, доказывающих тотипотентность клетки. При этом им убедительно показана полярность как органов, так и клеток. Основы экспериментальной эмбриологии растений были заложены исследованиями Моссарта (1902), который наблюдал набухание завязей некоторых растений после обработки их спорами Licopodium, нежизнеспособными поллиниями и водными экстрактами пыльцы. В связи с этим было высказано предположение, что пыльцевая трубка не только обеспечивает передвижение спермиев к яйцеклетке, но и переносит в завязь ауксины, стимулирующие ее рост. Г. Габерланд (1902) научился культивировать отдельные клетки в течение некоторого времени. Но он выбрал для культивирования зеленые клетки, изолированные из клеток палисадной паренхимы Lamium purpureum и волосков традесканции вирджинской и медуницы мягкой, резонно рассудив, что при этом отпадет потребность в источниках углеводов. Однако упустил из виду то, что ассимилирующие зеленые клетки — зрелые и высокодифференцированные, потеряли способность к меристематической деятельности. Исходное предположение автора, что содержащие хлорофилл клетки полностью обеспечивают себя питательными веществами, необходимыми для их жизнедеятельности и роста, не было подтверждено экспериментально. Габерланд также выдвинул гипотезу о тотипотентности любой живой клетки растения, которая впоследствии была подтверждена экспериментально. Ряд ученых, в том числе и его ученики, последовали его примеру и получили отрицательные результаты. Некоторые на основании этого усомнились в гипотезе тотипотентности растительных клеток. Исследования Габерландта с фотосинтезирующими клетками были неудачны, что привело к потере интереса к культивированию тканей и клеток растений. Однако они все же положили начало поиску адекватных питательных смесей и условий, необходимых для поддержания роста органов, тканей и клеток растений. Толчком к возобновлению работ послужили исследования Гаррисона, проведенные в 1904–1907 гг. Он вырастил нейробласты лягушки в лимфатической жидкости, доказав возможность выращивания in vitro изолированных клеток. Большое влияние на направление дальнейших работ с растительными клетками оказали работы зоологов Карреля и Барроуза (1911), культивировавших ткани и клетки млекопитающих на среде сложного состава, содержащей плазму крови и экстракты эмбриональных тканей. Французский ученый Мольяр уже в 1921 культивировал сегменты корня и гипокотиля молодых побегов редьки. Они были способны к росту в условиях культуры, но при этом не происходило формирования новых тканей. В 1922 г. один из учеников Рехингера — Коттэ начал эксперименты с лишенными пигментов меристематическими тканями — изолированными кончиками корней, и добился успеха. Практически одновременно и независимо от Коттэ Роббинс подобрал состав питательной среды, обеспечивающий в культуре рост апикальной меристемы корня томатов и кукурузы. Эти опыты положили начало культивированию изолированных органов растений на питательных средах. Не всегда эти исследования были успешны. Под влиянием работ Карреля и Барроуза в 1927 году Прат начал культивировать клетки растений на средах с добавками растительных экстрактов. Результаты его экспериментов были отрицательны, так как он избрал неудачные объекты для исследований. Начало длительным и удачным исследованиям по культивированию клеток и тканей растений положили работы американского исследователя Ф. Уайта и француза Р. Готре. Они показали, что изолированные органы и ткани могут расти в культуре неограниченно долгое время, если их пересаживать на свежую питательную среду. Такую же способность наблюдал Ф. Уайт для клеток опухолевого происхождения. Результаты чужих и собственных экспериментов Уайт обобщил в монографии «Культура растительных тканей», которая была переведена на русский язык и издана в СССР в 194 9 году. В ней он выделяет несколько периодов в истории развития метода культуры клеток, тканей и органов растений: 1. 1834–1900 гг. — создание и разработка клеточной теории. 2. 1900–1922 гг. — сформулирована идея культуры тканей. 3. 1922-1934 гг. — безуспешные поиски методов, обеспечивающих длительное культивирование тканей. 4. 1934-1939 гг. — детальная разработка техники культуры растительных тканей. Период 1940–1960 гг. значительно расширил список видов, выращиваемых in vitro. В монографию Готре, вышедшую в 1959 г., включено уже 142 вида. Были разработаны составы питательных сред, изучено значение микро- и макроэлементов для поддержания нормальной ростовой активности тканей, определено влияние витаминов и стимуляторов роста. Проводились работы по выявлению значения различных натуральных экстрактов (из эндосперма кокосового ореха, каштана, кукурузы и других растений) для поддержания неорганизованного клеточного роста, а также для стимуляции органогенеза. Показано значение кинетина для пролиферации клеток in vitro и индукции стеблевого морфогенеза. Изучением этих вопросов занимались такие ученые, как Р. Хеллер, И. Нич, Ф. Скуг, Ф. Стевард, Р. Г. Бутенко. В это же время разработаны методы получения и выращивания клеточных суспензий, а также культивирования отдельной клетки, деление ко-торой индуцируется с помощью ткани-няньки. В 1960–1975 гг. положено начало методу получения изолированных протопластов из тканей корня и плодов томатов путем обработки их смесью пектолитических и целлюлолитических ферментов. Основоположник этого метода — Э. Коккинг. Такебе с сотрудниками были определены условия культивирования изолированных протопластов, при которых они образуют клеточные стенки, делятся и дают начало клеточным линиям, способным к морфогенезу. Были разработаны методы гибридизации соматических клеток путем слияния протопластов и введения в них вирусных РНК, клеточных органелл, бактерий. В лабораториях Р. Г. Бутенко, Ю. Ю. Глебы проводились исследования поведения ядерного и цитоплазматического геномов партнеров в гибридных клеточных линиях и потомстве соматических гибридов растений — регенерантов. В этот же период были разработаны методы получения безвирусных растений из меристематических тканей. Начались эксперименты по созданию установок для глубинного культивирования клеток. Начиная с 1976 г., разрабатывались методы электрослияния протопластов и селекции гибридных клеток, культивирования гаплоидных клеток и получения новых форм и сортов сельскохозяйственных растений. Удалось создать системы иммобилизованных клеток для получения различных химических соединений и их биотрансформации. Ведутся работы по переносу генов в растительные клетки и получению трансгенных растений.
Культуры соматических клеток В основе культивирования растительных клеток лежит свойство тотипотентности, благодаря которому соматические клетки растения способны полностью реализовать наследственную информацию, то есть обеспечить развитие всего растения. Следует отметить, что в отличие от животной, растительная клетка предъявляет менее жесткие требования к условиям культивирования. Изменяя условия (добавляя в состав питательной среды те или иные гормоны), можно вызватьдифференциацию недетерминированных клеток. Культура растительной ткани позволяет получить многочисленные популяции в сравнительно короткое время и в ограниченном пространстве. Клетки в условиях in vitro лишаются очень многих важных взаимодействий, которые определяют их судьбу и дифференциацию в целом организме. В определенных пределах дифференциация культивируемых клеток поддается контролю со стороны экспериментатора. Основным типом культивируемой растительной клетки является каллус. Каллусная ткань — один из видов клеточной дифференцировки, возникает путем неорганизованной пролиферации дедифференцированных клеток органов растения. У растений в природе каллусная ткань возникает в исключительных обстоятельствах (например, при травмах) и функционирует непродолжительное время. Эта ткань защищает место поранения, может накапливать питательные вещества для анатомической регенерации или регенерации утраченного органа. Образование каллуса не всегда связано с травматическим воздействием. Каллус может возникнуть и в результате пролиферации внутренних тканей экспланта без связи с поверхностью среза из-за нарушения гормонального баланса. Растущий каллус разрывает слои ткани и развивается на поверхности. Для получения культивируемых каллусных клеток фрагменты тканей различных органов высших растений — корней, листьев, стеблей, пыльников, зародышей (экспланты) помещают на искусственную среду, содержащую ауксины, в пробирки, колбы, чашки Петри (in vitro). В качестве ауксинов используют 2,4-дихлорфеноксиуксусную кислоту (2,4-Д), α-нафтилуксусную кислоту (НУК), индолил-масляную кислоту (ИМК), индолилуксусную кислоту (ИУК) в концентрации 0,5-10 мг /л, в зависимости от вида экспланта. Процессу образования каллуса предшествует дедифференцировка тканей экспланта. При дедифференцировке ткани теряют структуру, характерную для их специфических функций в растении, и возвращаются к состоянию делящихся клеток. Если эксплант, используемый для получения каллуса, является фрагментом органа, то имеет в своем составе эпидермальные клетки, клетки камбия, сосудистой системы, сердцевинной и первичной коровой паренхимы. Преимущественно пролиферируют клетки камбия, коры, сердцевинной паренхимы. Различное тканевое происхождение каллусных клеток является одной из причин гетерогенности каллусной ткани, так как некоторые функциональные особенности исходных клеток передаются в ряду клеточных поколений как стойкие модификации. В качестве примера можно привести процессы, происходящие при дедифференцировке апикальной меристемы стебля. После помещения на питательную среду меристемы стебля томатов отмечено прекращение митоза, клетки увеличиваются в размерах, теряют характерную для меристематической ткани форму, изменяется структура ядра и цитоплазмы. В готовящейся к делению клетке возрастает синтез всех форм РНК, исчезают тканеспецифичные белки-антигены и появляются белки, специфичные для делящихся клеток и для каллусной ткани. Эти наблюдения свидетельствуют об изменениях в активности генов и белкового аппарата клетки при дедифференцировке. Активаторами матричной активности ДНК хроматина или активности РНК-полимеразы являются фитогормоны. Рецепторные для фитогормонов белки, локализованные в мембранах, по-видимому, оказывают влияние в присутствии фитогормонов на структуру и функцию мембран. Возможно, это обуславливает действие фитогормонов на генную активность. Одним из важнейших гормонов, применяемых при культивировании in vitro является ауксин, который активирует деление и растяжение клеток. Проникая в клетки, ИУК связывается со специфическими рецепторами, оказывая влияние на функциональную активность мембран, полирибосом и работу ядерного аппарата. Установлено, что в плазмалемме ауксин индуцирует работу Н+-помпы, в результате чего матрикс клеточных стенок размягчается, что является необходимым условием для роста и растяжения клеток. Включенная Н+-помпа усиливает поглотительную активность тканей, обогащенных ауксином. Предполагается, что поступление ауксина в клетку способствует усилению секреции кислых гидролаз и полисахаридов, необходимых для дальнейшего роста клеточных стенок. Под влиянием ауксина уменьшается продолжительность различных периодов митотического цикла. Так, предполагается, что уменьшается продолжительность периода удвоения числа клеток, продолжительность S-периода, Gi-периода[66]. Все это приводит к значительному ускорению темпов размножения клеток. Общим моментом в действии ауксинов на деление клеток является также предварительное усиление синтеза и накопление РНК. Стимулирующее действие ауксинов на синтез РНК может быть связано с восстановлением клеток после голодания перед их вхождением в митотический цикл, но может быть также приурочено к прохождению клетками этапов митотического цикла. Особенно отчетливо необходимость синтеза РНК проявляется при прохождении клетками Gi-периода. Под влиянием ауксина усиливается синтез р-РНК, но имеет место и появление новых информационных РНК, причем на очень ранних этапах действия. Осуществление клетками подготовки к делению на всех этапах митотического цикла зависит от синтеза белков. Ауксин вызывает как общую стимуляцию их синтеза, так и появление новых белков. Это позволяет предположить существование в хроматине структурных генов, транскрипция которых специфически индуцируется ауксином. Реализация действия ауксина на хроматин и последующее деление осуществляется вследствие его проникновения в цитоплазму, образования комплекса с цитоплазматическим ауксиновым рецептором и воздействием этого комплекса на транскрипционную активность хроматина. Кроме этого ауксин усиливает окислительную и фосфорилирующую активность митохондрий, в результате чего улучшается энергетическое и субстратное обеспечение процессов синтеза РНК и белков, репликация ДНК, а также осуществление самого митоза. Этот эффект обнаруживается очень рано и, как и синтез РНК, зависит от проникновения ауксина в клетку. Для возбуждения процессов подготовки к делению достаточно начального кратковременного действия ауксина. Поэтому процессы, происходящие в клетках под влиянием ауксина, можно разделить на первичные, непосредственно индуцированные ауксином, и вторичные, являющиеся следствием первичного индуцирующего действия. Исходя из этого, можно предположить, что в митотическом цикле растительных клеток имеются кратковременные переходы, когда необходимо присутствие ауксина в клетках, и более продолжительные периоды, когда присутствие ауксинов в клетке не является необходимым.
Морфофизиологическая характеристика каллусных тканей Выделяют два типа культивируемых растительных клеток: нормальные и опухолевые. Опухолевые клетки морфологически мало отличаются от каллусных. Физиологическим различием является гормононезависимость опухолевых клеток. Благодаря этому свойству опухолевые клетки делятся и растут на питательных средах без добавок фитогормонов. Эти клетки также лишены способности дать начало нормально организованным структурам (корни, побеги) в процессе органогенеза. Иногда они образуют тератомы (уродливые органоподобные структуры), дальнейшее развитие которых невозможно. Нормальные клетки в культуре могут существовать в двух видах: в виде суспензии в жидкой питательной среде и на поверхности твердой питательной среды в виде каллуса. Поверхностное культивирование осуществляют на полужидкой агаризованной среде, среде с добавлением других желирующих полимеров, на дисках из полиуретана, на мостиках из фильтровальной бумаги, полупогруженных в жидкую питательную среду. Можно также использовать комочки ваты, пропитанные питательной средой, которые сверху покрываются кусочком фильтровальной бумаги. Каллусная ткань, выращиваемая поверхностным способом, представляет собой аморфную массу тонкостенных паренхимных клеток, не имеющую строго определенной анатомической структуры. Цвет массы может быть белым, желтоватым, зеленым, красным. В зависимости от происхождения и условий выращивания каллусные ткани бывают: — рыхлые, сильно оводненные, легко распадающиеся на отдельные клетки; — средней плотности, с хорошо выраженными меристематическими очагами; — плотные, с зонами редуцированного камбия и сосудов. Как правило, в длительной культуре на средах, содержащих ауксины, каллусные ткани теряют пигментацию и становятся рыхлыми. В цикле выращивания каллусной ткани клетки после ряда делений приступают к росту растяжением, дифференцируются как зрелая каллусная ткань и деградируют. Для того чтобы не произошло старения, утраты способности к делению и дальнейшему росту, а также отмирания каллусных клеток, первичный каллус переносят на свежую питательную среду через 28–30 дней, то есть проводят пассирование или субкультивирование каллусной ткани. Неорганизованно растущая каллусная ткань характеризуется тремя типами клеток: мелкими, средними и крупными. При пассировании ткани на среду, содержащую индукторы органогенеза, мелкие клетки приступают к делению и формируют меристематические очаги. Деление клеток меристематического очага приводит либо к формированию почек и последующему развитию из них побегов (геммогенез), либо к ризогенезу (рис. 7, 8)[67]. Клетки меристемы с самых ранних стадий развития отличаются от каллусных высоким содержанием РНК и белка. При образовании соматических эмбриоидов каллусная клетка средних размеров обособляется, ограничивается плотной оболочкой, теряет крупные вакуоли. Она содержит крупное структурированное ядро с ядрышком. Клетка делится митотически, в результате чего возникают 2 клетки проэмбрио. Последующие деления клеток приводят к формированию шаровидного зародыша, а также органа, аналогичного суспензорам в зародышевом мешке семяпочки. Дальнейшее развитие соматического эмбриона через ряд стадий ведет к регенерации целого растения с корнями и побегами (рис. 9, 10), так как в этом случае формируется биполярная структура. Каллусы с высоким морфогенетическим потенциалом обычно матовые, компактные, структурированные, имеют зеленые хлорофиллсодержащие участки, которые представляют собой зоны морфогенеза. Впоследствии там формируются побеги или растения-регенеранты. В культуре также встречаются каллусы рыхлые, не имеющие глобулярного характера. Такие каллусы либо совсем не способны к органогенезу, либо формируют только корни. Появление корней свидетельствует о сдвиге гормонального баланса в сторону ауксинов, что препятствует образованию побегов. Эти каллусы могут остаться ризогенными, и регенерировать из них растения не удастся. Неморфогенные каллусы могут быть переведены в суспензионную культуру для получения вторичных метаболитов. Переход специализированных неделящихся клеток к пролиферации связан с их дедифференциацией, другими словами — утратой специализации. В основе этого процесса, как и при дифференциации клеток в интактном растении, лежит дифференциальная активность генов. Структура и функции клеток определяются активностью генов, и если клетки различаются по своей структуре и функциям, то это обусловлено различиями в экспрессии их генов, то есть специализация обеспечивается «включением» разных генов в разных клетках. Обычно активна небольшая часть (5 %) всего пула генов, свойственных данному виду. В этот состав активных генов входят, кроме видоспецифичных и обязательных для поддержания клеточного метаболизма, гены, активные только в данном органе, ткани, клетке, а также гены, активные лишь в определенном возрасте или начавшие работать только под влиянием изменившихся внешних условий. Возникновение физиологических и структурных различий между клетками и тканями растений, связанное с их функциональной специализацией, называют процессом дифференциации. Понятие «дифференциация» отражает превращение эмбриональной, меристематической клетки в специализированную. Меристематические клетки, однотипные по структуре и функции, начинают развиваться различными путями, создавая ткани разных органов. Как это осуществляется — один из труднейших вопросов клеточной биологии. Между геномами в клетках, которые приобретают разную форму и функцию, по-видимому, нет качественных различий, и клетки эти начинают различаться только вследствие разной экспрессии генов. Вновь возникшая клетка обладает широкими потенциями и может развиваться по любому из многих путей в морфологическом и физиологическом смысле. Детерминация (определение) пути развития каждой клетки является основой физиологии развития. Вступление на тот или иной путь развития определяется особым набором белков, т. е. каждая специализированная клетка вырабатывает только ей свойственные белки, что является следствием дифференциальной активности генов — экспрессии одной группы генов при одновременной репрессии других. Способность одной-единственной зрелой соматической клетки дать начало целому организму (тотипотентность) показывает, что в процессе нормальной клеточной дифференциации у растений не происходит утраты или необратимой инактивации каких-либо генов. У растений почти всякая дифференциация обратима при условии, если дифференцированная клетка живая, в протопласте сохранилось ядро и не образовалась вторичная оболочка. Даже такие высокоспециализированные клетки, как микроспоры, с помощью ряда экспериментальных процедур можно заставить пролиферировать и дать начало целому растению. Итак, в определенных условиях многие из зрелых растительных клеток сохраняют способность делиться, а в некоторых случаях даже вступить на новый путь развития. Однако вопрос о том. как это происходит, какие события на молекулярном уровне сопровождают этот процесс, остается открытым. Таким образом, после деления перед каждой дочерней клеткой открывается одна из трех возможностей. Клетка может оставаться эмбриональной и вновь вступить в клеточный цикл с последующим митозом либо может оказаться как бы «вне цикла» (G0), перестав делиться, и наконец, приобретя компетенцию, постепенно детерминироваться и вступить на путь дифференцировки (специализации). Компетенция — способность клетки воспринимать индуцирующее воздействие и специфически реагировать на него изменением развития. Индуцирующее воздействие могут оказывать различные факторы: гормоны, продукты жизнедеятельности соседних клеток, других тканей, электрофизиологические сигналы и т. д. Детерминация — приобретение клеткой состояния готовности к реализации определенных наследственных свойств. Детерминация приводит к развитию по определенному пути с одновременным ограничением возможности развития в других направлениях. Детерминация компетентной клетки может начинаться сразу же после деления в начале роста протоплазмы. Детерминированная определенным образом клетка приобретает узкую специализацию, то есть дифференцируется и превращается в клетку какой-либо ткани. Из гормональных факторов в дифференциации и морфогенезе первостепенная роль отводится ауксинам и цитокининам. В отношении органогенеза Скугом и Мурасиге была выдвинута концепция, согласно которой можно получить образование стеблей, корней или недифференцированный рост каллуса, изменяя относительное содержание ауксинов и цитокининов. В самом простом случае (табак) индукция и образование каллуса наблюдается при сбалансированном отношении ауксинов к цитокининам, стеблевые почки образуются при повышении уровня цитокининов по отношению к ауксинам, корни формируются при высоком содержании ауксинов в среде. В большинстве случаев формирование органов в культуре клеток можно объяснить гормональной теорией регуляции, но для некоторых видов она оказывается несостоятельной. Одни и те же физиологически активные вещества регулируют и деление клеток при недифференцированном росте каллуса, и клеточные деления, связанные с дифференциацией. Если дифференциация включает изменения в экспрессии генов на уровне транскрипции, тогда должны существовать и другие специфические вещества, определяющие компетентность клетки к обработке экзогенными фитогормонами. Есть системы, не реагирующие на гормональные обработки. Клетки этих тканей не способны к дифференциации, или, пролиферирующие in vitro, не проявляют тотипотентности. Морфогенная способность клона зависит от генотипа. Имеет значение и орган, от которого взят первичный эксплант. Ткани одного и того же органа имеют разную способность к морфогенезу. Например, флоэмная ткань корня моркови дает начало корням, а ксилемная — формирует эмбриоиды. Для понимания механизмов морфогенеза необходимо рассмотреть комплекс морфофизиологических процессов, протекающих в каллусной ткани, так как эти процессы лежат в основе того, что из каллуса при определенных условиях выращивания дифференцируется растение. В процессе культивирования отмечено возникновение новых микротрубочек в клетке in vitro, изменение ее размеров, активности рибосом, состояния пластидных пигментов, запасающих веществ. Роль микротрубочек, микрофиламентов и других структур в клеточном взаимодействии пока не выяснена. Эти структурные образования появляются обычно при регенерации и каким-то образом причастны к клеточному делению, взаимодействию между клетками, структуро- и формообразованию. Изменяется также структура и активность аппарата Гольджи, который выполняет важную функцию при клеточном делении. Появляются многоядерные клетки, отмечается полиплоидизация в результате нарушения митоза. Характерное несинхронное течение митотических циклов является одним из условий морфологической гетерогенности клеток ткани. Важно, что при выращивании in vitro наблюдается генетическая гетерогенность клеток, появление мутантов с отличительными особенностями органогенеза. В основе лежат изменения состояния хромосом в виде транслокаций, делеций, другие нарушения связаны с полиплоидизацией. Клетка, введенная в культуру, претерпевает последовательные изменения: переход к дедифференцированному состоянию, эмбриональному росту и, благодаря способности каллуса к вторичной дифференциации, формообразованию. Взаимодействие между клетками выступает как решающий фактор их дифференциации и специализации. Процесс дифференциации клеток обусловлен различной степенью репрессии и дерепрессии генетической информации. В ассоциации клеток каллусной ткани одни клетки занимают определенное положение и осуществляют посредством физико-химических контактов влияние на другие, чем определяется их структурно-функциональное состояние. Межклеточные взаимодействия осуществляются с помощью соответствующих донорно-акцепторных молекул цитоплазматической мембраны. Этими молекулами могут быть низкомолекулярные белки, комплексы углеводов с белками, фитогормоны, ингибиторы, полярные соединения и другие. Но во всех случаях на основе нуклеиново-белкового, белково-углеводного и иного типа узнавания они будут способствовать слипанию или отталкиванию клеток, будут выступать как эффекторы или апорепрессоры. В клетке реципиента с помощью специальных рецепторов эти молекулы будут связываться и изменять в эпигенезе реакцию генетической информации. Таким образом, в основе дифференциации клетки лежат процессы репрограммирования, репрессии, дерепрессии генетической информации. Это приводит к образованию специализированных клеток, которые становятся способными к взаимодействию, ассоциации, образованию геометрических форм, к органо- и морфогенезу. Важнейшим условием морфогенеза является адгезия клеток, в результате которой образуется ткань и орган. Поверхностные рецепторы, а также различные структуры типа микротрубочек обуславливают узнавание, сближение, слипание клеток в процессе дифференциации, ткане- и формообразования. Вещества, активные в процессах структуро- и формообразования, синтезируются под контролем ядра при поступлении сигналов из цитоплазмы клетки, а также экзогенных импульсов, эффекторов. При этом связующим звеном между генетической информацией, ее реализацией и эффектором выступают аллостерические белки, которые собирают, накапливают внешнюю информацию и преобразуют ее, в результате чего изменяют свою конформацию и вступают во взаимодействие с опероном. Генетическая обусловленность процессов морфогенеза отражается в изменении синтеза и-РНК, белков, активных ферментов, то есть в комплексе скоординированных во времени и пространстве реакций, обуславливающих дифференциацию активности генов. Появление некоторых белков свидетельствует об их участии в морфогенезе и запуске морфогенетических реакций. Установлен специфический фактор пептидной природы, стимулирующий морфогенез. Изучая генетический контроль каллусообразования и органогенеза, ученые предположили, что интенсивность образования каллуса находится под генетическим контролем. О генетической обусловленности признака регенерации в условиях in vitro свидетельствуют следующие факты: 1. Отсутствие определенных плеч хромосом (например, в клетках Triticum timopheevii при длительном культивировании теряются плечи хромосом генома At) может приводить к снижению выхода регенерантов. 2. С помощью гибридизации можно повысить интенсивность регенерации в каллусной ткани. 3. Использование разных по составу питательных сред для регенерации способствует разному уровню экспрессии генов, которые определяют этот признак. 4. В основе генетического контроля таких признаков, как частота каллусообразования, частота образования морфогенных каллусов и количество зон регенерации для озимой пшеницы основными являются сверхдоминирование, неполное доминирование и эпистаз; для озимой твердой — эпистаз, неполное доминирование и сверхдоминирование; для яровой твердой — эпистаз. Одни генетические системы контроля для всех признаков проявляются стабильно (эпистаз), а другие (сверхдоминирование) — значительно изменяются в зависимости от признаков и генотипов. Но следует отметить, что каллусогенез и регенерация растений не являются сопряженными процессами, вероятно, они контролируются различными генетическими механизмами. Общей закономерностью для культивируемых тканей остается возрастание цитогенетической вариабельности в процессе культивирования. С этим коррелирует в большинстве случаев потеря морфогенного потенциала. Способность к морфогенезу зависит и от состояния ядра. Как правило, регенерирующие в культуре тканей растения являются диплоидными, хотя ткани, из которых они произошли, имеют разный уровень плоидности. Таким образом, для индукции морфогенеза in vitro необходимо вызвать неоднородность в клеточных популяциях и тканях. Любые воздействия, приводящие к увеличению неоднородности в культуре клеток, в пространственном распределении гормонов, будут способствовать дифференциации клеток и формообразованию в каллусе. Доказательством этого могут также служить эксперименты, проведенные с каллусной тканью пшеницы и кукурузы в космических условиях. Эти эксперименты были описаны М. Карабаевым (1994). В условиях космического полета можно выделить 2 принципиальных стадии клеточного ответа на экстремальные условия: 1. Эта стадия, или стадия адаптации, продолжается 10–12 дней и связана с адаптацией культуры к стрессу. Она сопровождается общим уменьшением жизнеспособности клеток и потерей значительного числа клеток. В этих условиях число жизнеспособных, стрессоустойчивых клеток постепенно возрастает. 2. Инициируется деление и меняется распределение клеток в популяции, уменьшается градиент элементов питательной среды, так же как и градиент продуктов жизнедеятельности клеток. Независимо от продолжительности космического поле та, развитие клеток и структур, ответственных за клеточную дифференциацию, эмбриогенез и регенерацию растений подавляется космическими условиями. Основная причина этого может быть связана со специфическим распределением клеток в клеточной популяции и слабостью межклеточных контактов под действием невесомости. Анализ этих данных позволяет заключить, что гравитация имеет большое значение для развития растений, так как условия Земли способствуют более тесному взаимодействию гетерогенных спорадично растущих клеточных структур, а это впоследствии влияет на индукцию клеточной дифференциации.
Суспензионные культуры Суспензионные культуры — отдельные клетки или группы клеток, выращиваемые во взвешенном состоянии в жидкой среде. Представляют собой относительно гомогенную популяцию клеток, которую легко подвергнуть воздействию химических веществ. Суспензионные культуры широко используются в качестве модельных систем для изучения путей вторичного метаболизма, индукции ферментов и экспрессии генов, деградации чужеродных соединений, цитологических исследований и др. Признаком "хорошей" линии служит способность клеток к перестройке метаболизма и высокая скорость размножения в конкретных условиях культивирования. Морфологические характеристики такой линии: — высокая степень дезагрегации (5-10 клеток в группе); — морфологическая выравненность клеток (небольшие размеры, сферическая или овальная форма, плотная цитоплазма); — отсутствие трахеидоподобных элементов. Клеточную суспензию получают, помещая каллусную ткань в колбу с жидкой питательной средой. Суспензия перемешивается в колбе на качалке, имеющей скорость перемешивания 100–120 об/мин. При первом переносе на свежую среду удаляют крупные кусочки исходного каллуса и крупные агрегаты, фильтруя через 1–2 слоя марли, нейлоновые сита, шприц с соответствующим отверстием. Для инициализации суспензионной культуры необходимо 2–3 г свежей массы каллусной культуры на 60-100 мл жидкой питательной среды. Однако для каждой линии культуры клеток существует минимальный объем инокулята, при меньшем размере которого культура не растет. Рост суспензионных культур клеток можно оценивать по одному или нескольким следующим параметрам: 1. Объем осажденных клеток (ООК). Переносят небольшой объем суспензионной культуры в мерную пробирку объемом 15 мл, лучше всего коническую. Центрифугируют 5 минут при 200 д. ООК — величина, которую составляет объем осадка от объема суспензии, обычно в %. 2. Число клеток. Подсчитывается в камере Фукса-Розенталя. 3. Сырая и сухая масса. Суспензия клеток фильтруется через смоченный и взвешенный фильтр, вложенный в воронку Бюхнера под слабым вакуумом. Клетки промывают дистиллированной водой, оттягивают воду под вакуумом и взвешивают снова вместе с фильтром. Сухая масса — определяется аналогично, но взвешивается сухой фильтр, а клетки сушат вместе с фильтром в термостате при 60 °C до постоянной массы. 4. Содержание белка. Для определения белка клетки собирают на фильтре из стекловолокна, дважды промывают кипящим раствором 70 % этанола, сушат ацетоном, гидролизуют 1М NaOH при температуре 85 °C полтора часа. Затем фильтруют и определяют белок по Лоури. 5. Проводимость среды. Определяют с помощью кондуктометра. Как правило, она обратно пропорциональна свежей массе клеток. 6. Жизнеспособность клеток. Оценивают, изучая движение цитоплазмы под микроскопом, а также с помощью прижизненных красителей (флюоресцеиндиацетат, соли тетразолия, синий Эванса). Перед использованием подбирают pH инкубационного буфера, концентрацию красителя, время инкубации, строят калибровочные кривые для смеси живых и убитых клеток. По полученным данным строят ростовые кривые, которые имеют S-образную форму и состоят из нескольких участков: 1 — латентная, или лаг-фаза, где видимый рост не наблюдается ни по одному из критериев; 2 — экспоненциальная, рост с ускорением; 3 — линейная, где скорость роста постоянна; 4 — фаза замедленного роста; 5 — стационарная фаза; 6 — фаза деградации клеток (рис. 11)[68]. Реальная ростовая кривая может несколько отличаться от модельной. На форму ростовых кривых влияют и генетическая характеристика популяции (вид растения), и количество инокулята, и условия выращивания (состав среды, начальное значение pH, состав газовой фазы, скорость перемешивания). Необходимо отметить, что ростовые кривые для разных критериев не идентичны. Дисбаланс между скоростями клеточного размножения (число клеток), синтеза структурных элементов клетки (сухая масса) и увеличения объема и содержания вакуолей (сырая масса) отражает специфику онтогенеза высшего растения. Для глубинного культивирования растительных клеток применимы способы, разработанные в микробиологии. Различают два вида систем культивирования: открытую и закрытую. Для закрытой системы характерен периодический режим выращивания. Клеточная масса (инокулят) помещается в определенный объем среды. Система закрыта по всем параметрам, кроме газов, до конца выращивания. Периодически подается свежая питательная среда, а старая удаляется в том же объеме. Клетки остаются в системе в течение всего цикла выращивания. Открытые (проточные) культуры характеризуются поступлением свежей питательной среды, при котором отбирается не только старая питательная среда, но и часть урожая клеточной массы. Наиболее изучено и распространено закрытое глубинное культивирование. Для аэрации и перешивания используют различную аппаратуру: роллеры, качалки, магнитные мешалки и т. д. Очень большое значение для роста и биосинтеза клеток in vitro имеют технические характеристики систем культивирования. При масштабировании от небольших по объему культур в колбах до больших многолитровых ферментеров меняются многие параметры культивирования, в частности аэрация и перемешиваемость. Для культивирования суспензий в производственных масштабах применяется аппаратура, разработанная для микробиологической промышленности, однако исследования последних лет показали, что растительные клетки в силу своих специфических особенностей требуют особых сосудов для культивирования. Клетки растений в десятки, сотни раз крупнее клеток бактерий и грибов, кроме того, их размеры меняются в процессе онтогенеза. Если в начале экспоненциальной фазы роста они мелкие и плотные, то в стационарной фазе роста они сильно увеличиваются в размерах и вакуолизируются. Чем крупнее становится клетка, тем больше возрастает опасность ее механического повреждения в процессе перемешивания. В то же время клетки растений, крупные и тяжелые, требуют эффективного перемешивания. Оседание их приводит к появлению «мертвых» зон в сосудах, в которых происходит быстрое накопление и старение клеток. Для культуры клеток женьшеня отрицательное влияние механического стресса при выращивании в ферментере с турбинными мешалками сказывалось на жизнеспособности клеток уже при скоростях мешалок свыше 100–350 об/мин, это отрицательно влияло на синтез ими антрахинонов. Устойчивость штамма к механическому стрессу является важным требованием к культуре и трудной задачей для исследователей. Мягкое перемешивание и аэрацию обеспечивает пневматический способ перемешивания потоком сжатого стерильного воздуха, подаваемого в ферментер с восходящим током воздуха. К сожалению, и этот способ имеет свой недостаток, потому что в культуральной среде возникает избыток воздуха, приводящий к кислородному голоданию[69]. От концентрации кислорода в среде зависят рост и вторичный метаболизм клеток. В микробиологических системах изучена взаимозависимость роста биомассы, выхода искомого продукта и снабжения кислородом. Для растений таких данных нет. На рост клеток, кроме кислорода, могут влиять и другие газы. Например, углекислый газ может существенно влиять на длину лаг-фазы. Высокая степень аэрации может оказывать негативное действие на рост и синтез продуктов вторичного метаболизма, поскольку удаляются углекислый газ и летучие соединения. Клетки растений in vitro по сравнению с микроорганизмами имеют низкую интенсивность дыхания, что тоже должно учитываться при конструировании сосуда для культивирования. Сравнивали рост и образование метаболитов клетками в ферментерах разных типов. Клетки моринды лимонолистной, культивируемые в ферментерах с продувкой воздуха, содержали антрахинона на 30 % больше, чем в перемешиваемых колбах, и в два раза больше, чем в ферментерах других систем. Выход биомассы клеток не менялся в зависимости от типа биореактора. Клетки барвинка розового (Catharantus roseus) также синтезировали больше индольных алкалоидов при культивировании в ферментере с продувкой воздуха, чем в биореакторах с механическим перемешиванием. Отличительная особенность суспензионных культур клеток растений — высокая плотность, необходимая для роста. Поэтому другим осложнением при культивировании клеток растений является увеличение вязкости, сопровождающее рост биомассы. Это ведет к адгезии. Адгезия (прилипание) клеток друг к другу, на поверхностях культурального сосуда и погруженных в него мешалок и датчиков вызывает затруднения. В верхней части сосуда постепенно может образовываться пена, состоящая из выделяемых клетками белков и полисахаридов. В процессе культивирования клетки слипаются и часть из них скапливается в этой пене, образуя «корку», или «безе». С увеличением биомассы клеток увеличивается и эта «корка», снижая интенсивность перемешивания, что, в конце концов, может привести культуру к гибели. Клетки растений обладают меньшей физиологической и метаболической активностью по сравнению с микроорганизмами. Время генерации (интервал времени между двумя последовательными клеточными делениями) растительной клетки в 60-100 раз превосходит время генерации микробной клетки. Пул пролиферирующих клеток не превышает 50–60 %, многие клетки быстро прекращают деление и переходят в фазу покоя. Все эти обстоятельства определяют продолжительный рост популяции клеток при накопительном, или периодическом, выращивании. Поддержание стерильности длительное время также является одной из технических проблем, особенно при непрерывном культивировании. Периодическое, или накопительное, культивирование — это самый простой способ выращивания клеток, являющийся пока традиционным. Суспензионные культуры используют для промышленного получения вторичных метаболитов. Вещества, продуцируемые растительными клетками, используются в медицине, парфюмерной промышленности, растениеводстве и других отраслях промышленности. К ним относятся: алкалоиды, терпеноиды, гликозиды, полифенолы, полисахариды, эфирные масла, пигменты, антиканцерогены (птотецин, харрингтонин), пептиды (ингибиторы фитовирусов). В настоящее время в разных странах около ста видов растений используется в биосинтетической промышленности для получения экономически важных веществ, среди них — женьшень, раувольфия змеиная, наперстянка шерстистая и пурпурная, диоскорея дельтовидная, воробейник, беладонна, паслен дольчатый, дурман обыкновенный, ландыш майский, клещевина, агава, мак снотворный и др. Получение вторичных метаболитов имеет свои особенности. Деление клеток, приводящее к увеличению клеточной биомассы, и синтез вторичных метаболитов разобщены во времени. Накопление вторичных метаболитов возрастает в фазе замедленного роста клеточной популяции и достигает максимума в стационарной фазе. Некоторые алкалоиды активно синтезируются в фазе максимальной митотической активности (экспоненциальный рост), что является исключением. Знание таких закономерностей позволяет регулировать процессы получения ценных веществ. Механизмы и условия, блокирующие активный рост клеток и клеточную пролиферацию, одновременно активируют ферменты вторичного метаболизма. Неспецифические стрессовые условия, воздействующие на клетки в конце экспоненциальной фазы, могут стимулировать переход к синтезу вторичных метаболитов и увеличивать их выход. Необходимо учитывать, что вопрос взаимодействия первичного и вторичного метаболизма, рассмотренный нами в упрощенном виде, намного сложнее.
Культивирование отдельных клеток Отдельные клетки культивируют для получения клонов, изучения их генетической и физиологической изменчивости или стабильности. Кроме того, культивирование отдельных клеток позволяет изучать условия, определяющие возникновение стимулов к делению у клеток, изолированных от влияния других клеток популяции или ткани. Отдельные клетки также важны для клоновой селекции мутантных, гибридных и трансформированных линий. Обычно в такие клетки вводят маркерные гены, которые позволяют осуществлять селекцию. Кроме того, отдельные клетки могут служить моделью для сравнительного изучения физиологических процессов в ткани и изолированной клетке. Например, для изучения фотодыхания можно сравнивать процесс фотосинтеза на уровне отдельных клеток мезофилла листа и целой ткани. Выращивание изолированных клеток складывается из двух этапов: 1) изолирование неповрежденной клетки растительной или каллусной ткани; 2) создание условий, благоприятных для роста и развития изолированной клетки. На первом этапе необходимо выделить неповрежденную и жизнеспособную клетку из ткани целого растения или каллусной ткани. Этого можно достичь путем обработки ткани пектиназами, что ведет к мацерации ее клеток. Однако не всегда после такой обработки клетки сохраняют способность к последующим делениям и образованию ткани. Лучше получать отдельные клетки из суспензионных культур или рыхлого каллуса. Идеальными отдельными клетками являются протопласты, образовавшие клеточную стенку. Далее клетки изолируют либо при помощи микроманипуляторов, либо путем ряда последовательных разведений. При первых же попытках культивирования отдельных клеток возникла важная научная проблема: как заставить делиться клетки, изолированные от влияния других клеток популяции или тканей? Отдельные клетки вели себя иначе, чем их скопления в виде агрегатов в суспензии или каллусной массы на поверхности питательной среды. При ее решении возникла гипотеза о «факторе кондиционирования». Так было названо вещество, стимулирующее деление отдельных клеток. Определено, что этот фактор имеет химическую природу, термолабилен, водорастворим, низкомолекулярен (М.К. Павлова, Р.Г. Бутенко, 1965), видонеспецифичен, не заменяет известные фитогормоны, синергичен с брассиностероидами. Было предложено несколько вариантов культивирования отдельных клеток. Впервые подобрать условия, подходящие для деления отдельных клеток, удалось в 1954 году Мьюиру, Хильденбранту и Райкеру. Этот способ получил название метода «ткани — няньки» (рис. 12).

Рис. 12. Схема использования каллуса в качестве «ткани-няньки»
Клетку изолируют при помощи микроманипулятора из рыхлого каллуса непосредственно на кусочек фильтра размером 8*8 мм, помещенный на верхушку каллусной ткани, из которой была взята клетка. Каллус должен находится в фазе активного роста. Можно также в качестве «няньки» использовать каллусную ткань другого растения родственного вида. В этом случае клетки растут и делятся. По мере старения каллуса — няньки фильтр с клетками переносится на молодой каллус. Когда ткань из клетки достигает размеров 0,5–1 мм, то ее можно высаживать непосредственно на питательную среду. Проводились также эксперименты по высаживанию клетки непосредственно на агаризованную среду, но обязательно рядом с фильтром, который в течение нескольких дней контактировал с молодой, интенсивно растущей каллусной тканью. Поскольку эти работы показали, что постоянный контакт клетки через фильтр с каллусной массой не является обязательным для деления клетки, то было предложено использовать старую культуральную среду для стимуляции одиночной клетки к делению. Можно также использовать метод «кормящего слоя». Для этого берут суспензию клеток того же вида, что и одиночная клетка, или близкого вида. Клеточная суспензия должна находиться в ранней экспоненциальной фазе ростового цикла. В 1959 году Бергман предложил фильтровать суспензионную культуру (в его экспериментах это были табак и фасоль) стерильно через один слой батиста (ячейки 0,3*0,1 мм). В результате получали суспензию, на 90 % состоящую из отдельных клеток. Эту суспензию смешивали с агаризованной питательной средой того же состава, что использовался при культивировании суспензии (среда содержала 0,6 % агара). Смесь разливали тонким слоем (1 мм) в чашки Петри. Агар разделял клетки, но не препятствовал обмену химическими сигналами между ними, а толщина слоя позволяла смотреть за их поведением под микроскопом. Индукция делений отдельных клеток возможна при применении очень богатой питательной среды, например, среды Као и Михайлюка. При этом объем среды, в которую помещаются клетки, должен был минимальным (микрокапли объемом до 20 мкл). Все эти способы культивирования позволяют клетке «ощущать» фактор кондиционирования. Он либо вырабатывается в достаточном количестве клетками «кормящего слоя», «ткани — няньки», либо содержится в суспензии, где ранее культивировались клетки, либо не теряется в большом объеме среды. Таким образом, фактор, вызывающий деление клеток, вырабатывается самими клетками, но в небольшом количестве. И только увеличивая число клеток, вырабатывающих этот фактор, чтобы он не рассеивался в больших объемах питательной среды, или же уменьшая объем среды, в котором будет выращиваться клетка, можно заставить ее делить-
Культуры гаплоидных клеток Большой интерес для селекционеров представляют гаплоидные растения. Гаплоиды получают двумя способами. Первый способ классический — отдаленная гибридизация, когда в зиготе отдаленного гибрида хромосомы одного из видов элиминируют. Второй способ основан на методиках культивирования in vitro, где из неоплодотворенных половых клеток с редуцированным набором хромосом можно регенерировать целые растения. Обычно они стерильны, так как у них нарушено формирование мужских и женских гамет. При культивировании in vitro, однако, может произойти спонтанное удвоение хромосом, или его можно вызвать искусственно, например, обработав колхицином клетки илирастения. Дигаплоиды фертильны и вполне жизнеспособны. Гаплоиды и дигаплоиды имеют ряд преимуществ в селекционной работе: • гаплоидные растения имеют один набор хромосом, характерный для гамет, что дает селекционерам возможность наблюдать мутации сразу же в ходе осмотра гаплоидных растений, поскольку все рецессивные генные мутации в гаплоидных организмах не маскируются доминантными аллелями; • если гаплоидные клетки подвергнуть полиплоидизации с помощью колхицина, то возникнут дигаплоиды, характеризующиеся абсолютной гомозиготностью. Скрещивание гомозиготных линий дает, как правило, высокопродуктивное потомство. С другой стороны, в настоящее время картофель не размножают семенами из-за пестроты потомства, а создание с помощью гаплоидов гомозиготных линий устранит этот недостаток; • гомозиготные растения используются селекционерами и в других целях: количественный генетический анализ, изучение взаимодействия генов, изучение генетической изменчивости, определение групп сцепления, установление числа генов, действующих на количественные признаки, определение локализации полигенов и т. д. • гаплоидные растения лишены летальных или сублетальных мутаций, ведущих к гибели или ослаблению потомства. Гаплоиды высших растений можно получить из эксплантов, взятых на любой стадии развития гаметофита после редукционного деления клеток спорогенной ткани пыльника. Наиболее распространены следующие методы индуцирования гаплоидов: 1. Индуцированный андрогенез в культуре пыльников и пыльцы. 2. Селективная элиминация хромосом в гибридном зародыше. Этот метод чаще всего используется в селекции злаковых. 3. Псевдогамия — развитие гаплоидного зародыша после оплодотворения инородной пыльцой без оплодотворения яйцеклетки или же развитие изолированной семяпочки (гиногенез). В клеточной инженерии чаще применяется первый метод. Впервые гаплоидные растения были получены в 1964 году индийскими исследователями С. Гуха и С. Махешвари при культивировании пыльников дурмана. С тех пор таким методом получены гаплоидные растения более чем у 200 видов, в том числе у пшеницы, ячменя, ржи, риса, картофеля и других культур. Для культуры пыльников используют целые пыльники, стерильно выделенные из бутонов в определенной фазе развития. Их помещают на твердую питательную среду, либо на поверхность жидкой питательной среды. В редких случая культивируют бутоны или соцветия. Получение гаплоидных растений из изолированных пыльников может идти по двум направлениям: прямая регенерация соматических зародышей и косвенная — через каллусогенез. В первом случае внутри пыльников из отдельных пыльцевых зерен формируются проэмбриональные структуры, которые при определенных условиях культивирования развиваются в эмбриоиды, дающие начало гаплоидным растениям. Эмбриоиды — зародышеподобные структуры. Во втором — пыльца делится, но клетки, возникшие в результате делений, быстро увеличиваются в размерах и, разрывая оболочку пыльцевого зерна, образуют каллус. В результате дальнейшего морфогенеза из этих каллусных клеток регенерируют растения. При этом растения могут иметь разную степень плоидности — ди, поли, анеуплоидные. Последние часто стерильны, но после обработки растений колхицином происходит удвоение числа хромосом, в результате чего можно получить фертильные гомозиготы. Культура пыльцы представляет собой культивирование микроспор, освобожденных от соматических тканей пыльника, в жидкой среде. Пыльцу от соматической ткани пыльника отделяют несколькими способами: 1. Спонтанное высвобождение (пассивный способ) — пыльники определенным образом обрабатываются, инкубируются на жидкой среде, где лопаются, а пыльца высвобождается и всплывает наверх. 2. Гомогенизация и фильтрация. Пыльники, культивируемые в жидкой среде, разрушают, надрезая скальпелем и осторожно надавливая, затем фильтруют (поры фильтра 50 -100 мкм) и центрифугируют. Осадок промывают и суспендируют в жидкой среде. 3. Разрезание — разрезают стенку пыльника. Этот метод применяется редко, так как трудоемок и длителен. Пыльцевой эмбриогенез обусловлен функциональной и структурной детерминацией пыльцевого ядра и клеток гаметофита, поэтому в развитии могут принимать участие: — лишь вегетативные клетки, — лишь генеративные клетки, — оба типа клеток, если вегетативные и генеративные клетки сольются, при этом образуется диплоидный эмбриоид. Для пасленовых характерен только эмбриогенез, для злаковых — образование, как каллусов, так и эмбриоидов. Среди гаплоидов много альбиносов (особенно у злаков). Наибольший выход регенерантов-альбиносов в культуре пыльцы, что вызвано, по-видимому, нарушениями развития пыльцы. Причина не установлена, возможно, это результат мутаций в микроспорах при культивировании. При отдаленной гибридизации некоторых видов установлено явление селективной элиминации хромосом одного из родителей на ранней стадии развития гибридного зародыша. Это явление хорошо изучено у ячменя. При скрещивании диплоидных ячменей Hordeum vulgare (культурный) и Н. bulbosum (многолетний луковичный дикий) на стадии роста зародыша и эндосперма (через 5 дней после оплодотворения) происходит выпад хромосом дикого вида. Возникает гаплоид с набором хромосом Н. vulgare. Через 15 суток после оплодотворения рост гибридного зародыша на материнском растении прекращается, но при культивировании in vitro из таких зародышей развиваются проростки. Частота и количество образовавшихся растений при этом способе очень высоки. Кроме того, растения-альбиносы не образуются. С помощью этого метода были выведены сорта Исток и Одесский-115 — за 4 года вместо 10–12 лет обычной селекции. В Канаде так были получены сорта Минго и Родео. Элиминация хромосом встречается и у других родов. Если в качестве опылителя использовать дикий ячмень, то можно индуцировать гаплоиды у ржи и пшеницы. Работы по получению гаплоидов в культуре женского гаметофита начались в 50-е годы. В последнее время интерес к ним возрос. У растений с мужской стерильностью культивирование неоплодотворенных семяпочек является единственной возможностью получения гаплоидов. Женский гаметофит может быть источником получения гаплоидов и у растений с низким морфогенетическим потенциалом каллусной ткани, либо если каллусная ткань регенерирует растения-альбиносы. У некоторых растений, например у ячменя и риса, индукция зеленых растений намного выше при гиногенезе по сравнению с андрогенезом. В зависимости от того, какая клетка зародыша даст начало новому организму, различают партеногенез и апогамию. Партеногенез — развитие яйцеклетки без оплодотворения. При апогамии зародыш развивается из синергиды или антиподы. В ранних работах наблюдалась пролиферация соматических тканей зародышевого мешка. Впервые гаплоидный каллус из неоплодотворенной семяпочки был получен в 1964 году Тулеком в культуре гингко, но органогенез индуцировать не удалось. Это случилось лишь в 1976 году, когда Сан Ноум при работе с культурой неоплодотворенных завязей ячменя получил нормальные зеленые гаплоидные растения. Гиногенез может идти двумя путями — через эмбриогенез и через каллусогенез. В работах Сан Наума с ячменем было показано, что гаплоидные эмбриоиды преимущественно образовывались из антипод, а каллус — из синергид. У риса и эмбиогенез, и каллусогенез давали синергиды, а антиподы в итоге дегенерировали. У табака гаплоидный эмбриогенез характерен для яйцеклеток, у скерды — для антипод.
НОВЫЕ ЭКСПЕРИМЕНТАЛЬНЫЕ СИСТЕМЫ ДЛЯ ИЗУЧЕНИЯ СИНТЕЗА ВТОРИЧНЫХ МЕТАБОЛИТОВ С ИСПОЛЬЗОВАНИЕМ КУЛЬТУРЫ ТКАНЕЙ РАСТЕНИЙ
Иммобилизация растительных клеток: необходимость, основные методы Культуры клеток и тканей растений считаются потенциальным источником специфических вторичных метаболитов, к которым относятся такие соединения, как алкалоиды, стероиды, масла и пигменты. Многие из этих веществ все еще получают путем экстракции из растений. Не ко всем видам растений в настоящее время применимы методы микробиологической промышленности. За исключением некоторых видов растений, суспензионные и каллусные культуры клеток синтезируют вторичные метаболиты в меньших количествах, чем целые растения. При этом рост биомассы в ферментере может быть значительным. Новым подходом, направленным на увеличение выхода вторичных метаболитов, является иммобилизация клеток и тканей растений. Первая удачная попытка зафиксировать целые клетки была осуществлена в 1966 г. Мосбахом. Он зафиксировал клетки лишайник[70] Umbilicaria pustulata в полиакриламидном геле. На следующий год ван Вецель выращивал клетки эмбрионов животных, иммобилизованных на микрошариках ДЭАЗ (диэтиламиноэтил сефадекса, на основе декстрана). После этого клетки были иммобилизованы на разных субтратах. В основном это были клетки микроорганизмов. Методы иммобилизации клеток делят на 4 категории: 1. Иммобилизация клеток или субклеточных органелл в инертном субстрате. Например, rktnrb Catharanthus roseus, Digitalis lanata в альгинатных, агарозных шариках, в желатине и т. д. Метод предполагает обволакивание клеток одной из различных цементирующих сред — альгинат, агар, коллаген, полиакриламид. 2. Адсорбция клеток на инертном субстрате. Клетки прилипают к заряженным шарикам из альгината, полистирола, полиакриламида. Метод применялся в экспериментах с животными клетками, а также клетками Saccharomyces uvarum, S. cerevisiae, Candida tropicalis, E. coli. 3. Адсорбция клеток на инертном субстрате с помощью биологических макромолекул (таких, как лектин). Применяется редко, есть сведения об экспериментах с различными линиями клеток человека, эритроцитами крови барана, адсорбированными на покрытой белком агарозе. 4. Ковалентное связывание с другим инертным носителем типа КМЦ. Очень редко применяется, известна удачная иммобилизация для Micrococcus luteus. В основном проводились эксперименты по иммобилизации клеток животных и микроорганизмов. В последнее время интерес к иммобилизации клеток растений значительно возрос, это связано с тем, что иммобилизованные клетки имеют определенные преимущества перед каллусными и суспензионными культурами при использовании их для получения вторичных метаболитов.
Физиологические основы преимущества иммобилизованных растительных клеток перед традиционными способами культивирования В литературе имеются многочисленные данные о том, что существует положительная корреляция между накоплением вторичных метаболитов и степенью дифференцировки в культуре клеток. Кроме того, лигнин, например, откладывается в трахеидах и сосудистых элементах ксилемы только после завершения процессов дифференцировки, что было показано в экспериментах как in vivo, так и in vitro. Полученные данные свидетельствуют о том, что дифференциация и накопление вторичных продуктов обмена веществ происходит в конце клеточного цикла. При снижении роста процессы дифференциации ускоряются. Изучение содержания алкалоидов, накапливаемых многими растениями in vitro, показало, что компактные, медленно растущие культуры клеток содержат алкалоиды в больших количествах, чем рыхлые, быстро растущие культуры. Организация клеток необходима для их нормального метаболизма. Наличие организованности в ткани и ее последующее действие на различные физические и химические градиенты — четкие показатели, по которым различаются высоко- и низкопродуктивные культуры. Очевидно, что иммобилизация клеток обеспечивает условия, приводящие к дифференциации, упорядочивает организацию клеток и способствует тем самым высокому выходу вторичных метаболитов. 1. Клетки, иммобилизованные в или на инертном субстрате, образуют биомассу гораздо медленнее, чем растущие в жидких суспензионных культурах. Какова же связь между ростом и метаболизмом? При чем здесь клеточная организация и дифференцировка? Предполагают, что эта взаимосвязь обусловлена двумя типами механизмов. Первый механизм основан на том, что рост определяет степень агрегации клеток, оказывая косвенное влияние на синтез вторичных метаболитов. Организация в данном случае является результатом агрегации клеток, а достаточная степень агрегации может быть получена только в медленно растущих культурах. Второй механизм связан с кинетикой скорости роста и предполагает, что «первичный» и «вторичный» пути метаболизма по-разному конкурируют за предшественники в быстро и медленно растущих клетках. Если условия среды благоприятны для быстрого роста, то в первую очередь синтезируются первичные метаболиты. Если быстрый рост блокирован, то начинается синтез вторичных метаболитов. Таким образом, низкая скорость роста иммобилизованных клеток способствует высокому выходу метаболитов. 2. Кроме медленного роста иммобилизация клеток позволяет им расти в тесном физическом контакте друг другом, что благоприятно отражается и на химических контактах. В растении любая клетка окружена другими клетками, но ее положение меняется в ходе онтогенеза в результате деления как этой, так и окружающих клеток. От положения клетки в растении зависит степень и тип дифференциации этой клетки. Следовательно, физическое окружение клетки влияет на ее метаболизм. Каким образом? Регуляция синтеза вторичных метаболитов находится как под генетическим, так и под эпигенетическим (внеядерным) контролем, то есть любые изменения в цитоплазме могут привести к количественным и качественным изменениям в образовании вторичных метаболитов. В свою очередь, цитоплазма представляет собой динамическую систему, находящуюся под влиянием окружающей среды. Из внешних условий на метаболизм существенное влияние оказывают 2 важных фактора: концентрация кислорода и углекислого газа, а также уровень освещения. Свет играет роль и в процессе фотосинтеза, и в таких физиологических процессах, как деление клеток, ориентация микрофибрилл, активация ферментов. Интенсивность и длина световой волны определяется положением клетки в массе других клеток, то есть зависят от степени организованности ткани. В организованной структуре существуют центробежные градиенты концентрации О2 и СО2, которые играют исключительно важную роль в процессе дифференциации. Таким образом, вторичный метаболизм в крупных агрегатах клеток с небольшим отношением площади к объему (S/V) отличается от такового изолированных клеток и мелких групп клеток в результате действия градиентов концентрации газов. Аналогично действуют градиенты регуляторов роста, питательных веществ, механического давления. Условия окружения у диспергированных клеток и клеток в виде агрегатов различны, поэтому пути метаболизма у них также различаются. 3. Регулировать выход вторичных метаболитов можно также, изменяя химический состав окружающей среды. Изменение состава среды для каллусной и суспензионной культуры сопровождается определенными физическим манипуляциями с клетками, что может привести к повреждению или загрязнению культур. Эти трудности можно преодолеть, используя циркуляцию больших объемов питательной среды вокруг физически неподвижных клеток, что позволяет осуществлять последовательные химические воздействия. 4. В некоторых случаях возникают проблемы с выделением идиолитов. При использовании иммобилизованных клеток относительно легко осуществляется обработка их химическим веществами, индуцирующими высвобождение требуемых продуктов. Это также снижает ингибирование по типу обратной связи, которое ограничивает синтез веществ вследствие накопления их внутри клетки. Культивируемые клетки некоторых растений, например, Capsicum frutescens выделяют вторичные метаболиты в окружающую среду, а система иммобилизованных клеток позволяет отбирать продукты без повреждения культур. Таким образом, иммобилизация клеток способствует легкой изоляции идиолитов.
Системы культивирования иммобилизованных клеток Существует 2 типа систем культивирования иммобилизованных клеток: 1. Система культуры с плоской основой, клетки выращиваются в горизонтально расположенном сосуде. 2. Система колоночной культуры, где клетки выращиваются в вертикальном сосуде. В обеих системах жидкая среда циркулирует вокруг физически неподвижных клеток. Система культуры с плоской основой (рис. 13)
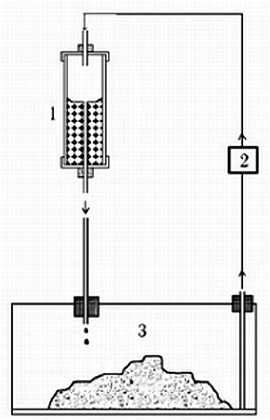
Рис. 13. Система культуры с плоской основой
Питательная среда капает под действием силы тяжести из цилиндрического сосуда объемом 70 мл (1) в стеклянный сосуд для культивирования (3) объемом 350 мл, где находятся клетки (40–50 г сырой массы), посаженные на субстрат — подстилку из нетоксичной полипропиленовой ткани. Питательная среда проникает сквозь ткань под действием капиллярных сил, снабжая клетки. После этого использованная питательная среда откачивается из сосуда для культивирования с помощью перистальтического насоса (2) обратно в резервуар и используется повторно. Исследования, проведенные с этой системой, показали, что клетки Solanum niger, культивируемые на плоской основе (среда Мурасиге-Скуга с добавлением 2,4-Д и кинетина) способны потреблять питательные вещества и быстро реагируют на недостаток ортофосфата. Увеличение сырой массы идет гораздо медленнее, чем в суспензионных культурах. Количество жизнеспособных клеток такое же и достигает 70–80 %. Количество алкалоидов через 7 суток культивирования достигает 12 мг/г сухой массы, в суспензии же через 18 дней культивирования оно составляет 10 мг/г сухой массы. В местах, где питательная среда капает прямо на клетки, через 3–4 дня культивируемая ткань становится темной и отличается от остальной, которая у S. niger окрашена в светло-бежевый цвет. Клетки в зонах капания часто бывают более компактны, содержат больше алкалоидов. Если клетки изолировать из зон капания и поместить в чашки Петри с агаром, содержание алкалоидов падает, при возвращении в прежние условия культивирования вновь аккумулируются высокие количества метаболитов. Кроме того, в клетки вводились предшественники вторичных метаболитов. Отмечено, что синтез капсацина клетками Capsicum frutencens увеличивается в тысячи раз при добавлении в среду 5 мл изокаприновой кислоты. При этом капсацин не накапливается внутри клеток, а секретируется в питательную среду. Установлено, что клетки, иммобилизованные на плоской основе, способны потреблять питательные вещества из среды, в том числе и кислород, имеют низкую скорость роста, каллусоподобное расположение, способны к тесному межклеточному контакту. В целом, они имеют более высокий уровень синтеза вторичных метаболитов. Несмотря на эти преимущества, промышленное культивирование имеет существенный недостаток: горизонтальная конструкция аппарата создает неудобства при работе и требует большой площади. Эти недостатки устраняются в другой системе. Система колоночной культуры (рис. 14)
Рис. 14. Система культуры в колонке
В такой системе возрастает число клеток, на которые капает среда, что увеличивает "зону капания", где происходит накопление больших количеств вторичных метаболитов. Сосуд для культивирования из горизонтального превращается в вертикальный. В сосуде с питательной средой (1) 50 мл жидкой среды, которая под действием силы тяжести капает в вертикальную стеклянную колонку, содержащую иммобилизованные клетки (3). Среду собирают со дна колонки и вновь используют в цикле, перекачивая с помощью перистальтического насоса в резервуар со средой. Как разместить клетки внутри колонки, чтобы капающая среда не спрессовала их в плотную массу на дне колонки? Закрепить в нейлоновую сеть, корзиночки, используя инертный, проницаемый и стабильный гель. Между корзиночками образуется воздушное пространство, сеточка структурирует гель, а клетки могут прорастать через корзиночки, контактируя друг с другом. Подходящим материалом для размещения клеток оказались нейлоновые мочалки. Материал нетоксичен, легко режется, выдерживает автоклавирование. В качестве субстратов для погружения клеток используют агар и альгинат кальция. Агар традиционно используется в работе с культурами клеток и тканей. 2 % (масса к объему) раствор агара в дистиллированной воде автоклавируют при 1 атмосфере в течение 20 минут и охлаждают на водяной бане до 35–40 °C. Клетки суспензионной культуры в стационарной фазе роста пропускают через сито с диаметром пор 1 мм и смешивают в пропорции 1:1 (V: V) с незастывшим агаром. Стерильные кусочки мочалки 2–3*1*1 см с помощью стерильного пинцета опускают в смесь клеток с агаром, а когда агар начинает застывать, помещают в стерильные колонки с внутренним диаметром 15*2,5 см. В каждую колонку помещают примерно 10 сеточек, таким образом, на колонку приходится 5–7 г сырой массы клеток. Аналогично иммобилизуют клетки с использованием альгината кальция. В этом инертном субстрате, например, успешно были иммобилизованы А. Альферманном с сотрудниками клетки наперстянки шерстистой (Digitalis lanata). 2 % раствор альгината натрия автоклавируют, охлаждают до комнатной температуры, смешивают с клетками и переносят в стерильный раствор 0,05 М СаСl2 в дистиллированной воде на 10 минут, чтобы образовывающийся альгинат кальция затвердел внутри и вокруг кусочков мочалки, обволакивая клетки. Молекулы альгината поперечно сшиваются катионами кальция, при этом происходит его стабилизация. Затвердевший материал трижды промывают в стерильной дистиллированной воде и вносят в стеклянные колонки. Как показали дальнейшие исследования, рост клеток в альгинате был лучшим, чем в агаре, что связано, вероятно, с негативным действием расплавленного агара, температура которого составляет около 40 °C, на клетки в процессе иммобилизации. Поэтому эксперименты по изменению условий окружающей среды проводились в колонках, где клетки Datura innoxia и Capsicum frutencens были иммобилизованы в альгинате кальция. Скорость потребления ортофосфата, нитратов, аммония и сахарозы в клетках колоночной культуры ниже, чем в горизонтальной системе. Жизнеспособность клеток, по сравнению с суспензионными культурами, заметно не снижается (60–65 %), содержание алкалоидов через 8-10 суток составляет 12–13 мг/г сухой массы клеток, алкалоиды в питательную среду не выделяются, а pH среды после 8 суток культивирования снижается на 0,4 единицы, до 5,4. Определение скорости потребления кислорода показывает предположительную нехватку его на 4–8 сутки культивирования. Культуры, освещаемые люминесцентными лампами, потребляют питательные вещества интенсивнее, рост клеток лучше также в освещенных культурах. В светокультуре повышается жизнеспособность клеток и содержание алкалоидов. Так как клетки не зеленеют, для поддержания нормального клеточного метаболизма необходимы и другие световые эффекты. Добавление предшественников не всегда однозначно влияет на рост, потребление питательных веществ и синтез метаболитов. Например, действие орнитина на клетки Datura innoxia скорее отрицательно, а изокаприновой кислоты на клетки Capsicum frutencens — скорее положительно, так как сырая масса увеличивается, но выход капсомицина остается прежним. В обоих случаях синтез метаболитов не ингибируется альгинатным гелем. Полученные результаты свидетельствуют о сохранении жизнеспособности клеток многих видов растений при иммобилизации. Иммобилизованные клетки, полученные из рыхлых, быстро растущих суспензионных культур, замедляют рост и приобретают сходство с клетками каллусной культуры. Эти клетки более взаимосвязаны, чем в жидкой культуре, что ведет к возникновению определенного физического и химического градиентов. Иммобилизованные клетки аккумулируют больше алкалоидов, чем в жидкой культуре, что ведет к возникновению определенных физических и химических градиентов. Это способствует переходу клеток в стационарную фазу, что приводит к ускорению образования алкалоидов и других веществ вторичного происхождения. Общие рекомендации к культивированию клеток на основе полученных результатов. 1. Клетки должны выращиваться физически стационарно, в тесном контакте друг с другом, чтобы стимулировалось развитие физических и химических градиентов и обеспечивалась частичная дифференцировка культуры. Некоторые виды растений необходимо культивировать при освещении и индуцировать образование хлоропластов, чтобы обеспечить уровень метаболизма, близкий к происходящему в клетках интактного растения. 2. Состав питательной среды и уровень кислорода необходимо регулировать для замедления роста культуры. Рекомендуется использовать регуляторы роста для имитации процессов дифференциации, происходящих in vivo. 3. Клетки необходимо снабжать предшественниками, но в низких концентрациях. Предшественники должны быть максимально близки в цепочке превращений к исходному продукту. 4. Желательно использовать клетки, которые секретируют необходимые метаболиты в питательную среду или клетки, у которых такую секрецию можно индуцировать.
ПРОТОПЛАСТЫ РАСТИТЕЛЬНЫХ КЛЕТОК КАК ОБЪЕКТ БИОЛОГИЧЕСКОГО КОНСТРУИРОВАНИЯ
Применение изолированных протопластов Протопласты являются уникальной моделью для изучения фундаментальных физиологических проблем у растений. Они незаменимы при изучении состава, структуры и функционирования плазмалеммы в норме и при воздействии на нее гормонами, ингибиторами, фитототоксинами, а также при взаимодействии самих протопластов в популяции. Кроме того, протопласты могут использоваться для определения состава и архитектоники первичной клеточной стенки и изучения механизма ее репарации после разрушения. На схеме (рис. 15) представлены основные направления физиологических исследований с использованием культуры изолированных протопластов.
Рис. 15 (ниже). Изолированные протопласты — объект и модель в физиологических исследованиях (по Р.Г. Бутенко, 1981)
Таким образом, изолированные протопласты имеют ряд областей применения, как теоретического, так и прикладного характера: 1. Изучение химии и структуры клеточной стенки (и при разрушении, и при синтезе «de novo»). 2. Изучение свойств плазмалеммы, трансмембранных перемещений. 3. «Мягкое» выделение органелл. 4. Наблюдение за закономерностями дифференцировки клеток при слиянии протопластов, отслеживание взаимодействия ядра и цитоплазмы в полученной гибридной клетке, изучение соматических гибридов. 5. Введение чужих органелл. 6. Введение чужеродных генов в растительную клетку (трансгенез).
Способы получения и культивирования протопластов
Получение протопластов Протопласт — клетка, лишенная целлюлозной оболочки, окруженная цитоплазматической мембраной, сохраняющая все свойства, присущие растительной клетке. Впервые протопласты в 1892 г. выделил Дж. Клеркер, который использовал механический способ. При этом способе у плазмолированных клеток разрезают клеточную стенку, протопласты выходят в среду. В настоящее время метод претерпел модификации, улучшен, но имеет ряд ограничений: — невысокая производительность, — можно использовать ткани только с экстенсивным плазмолизом, трудоемкость и длительность, Другой метод выделения протопластов — энзиматический, с использованием ферментов. В 1952 году Салтон с помощью фермента лизоцима впервые разрушил клеточную стенку бактерий. В 1960 году Коккинг обработал кончики корней томата гидролитическим ферментом из культуральной жидкости плесневых грибов (Myrothecium verrucaria) и впервые получил изолированные протопласты высших растений энзиматическим способом. Преимущества энзиматического метода по сравнению с механическим: — одновременно выделяется большое количество протопластов (до 10 млн. из грамма ткани или клеток), — клетки не подвергаются сильному осмотическому стрессу, — клетки не повреждаются, — метод сравнительно быстрый. Для удаления клеточной стенки используют ферменты трех типов: целлюлазы, гемицеллюлазы и пектиназы. Комбинация ферментов и их соотношение специфично для каждого типа клеток. Выделение протопластов проводят в три этапа: — обработка ферментами, — выделение протопластов из клеточных стенок, — отделение интактных протопластов от клеточных осколков. Стандартная методика протопластов (по Такебе) из тканей листа Nicotiana tabacum: Зрелый, сформировавшийся лист отделяют от взрослого растения в возрасте 60–80 дней, окунают в 70 % этанол, а затем помещают на 15–20 минут в 10 % раствор гипохлорита кальция и многократно промывают дистиллированной водой. С помощью пинцета нижний эпидермис снимают, очищенные от эпидермиса листья разрезают скальпелем на небольшие кусочки площадью 4 кв. см. Для лучшего снятия эпидермиса листья должны немного подвянуть, можно также ограничить снабжение водой перед срезанием листьев. Далее ткань обрабатывают последовательно или одновременно пектиназой, вызывающей мацерацию, и целлюлазой, разрушающей клеточные стенки. Оптимальная концентрация ферментов, как и время обработки, индивидуальны для разных тканей. Протопласты должны находиться в растворе ферментов минимальное количество времени, после чего следует тщательная промывка. Ферменты стерилизуют через бактериальные фильтры. Регуляция водообмена клетки связана с наличием клеточной стенки. Когда протопласт "голый", один из компонентов регуляции водообмена теряется, поэтому важное значение приобретают осмотические свойства среды выделения и культивирования. Среда должна быть немного гипертонической, чтобы протопласты находились в слегка плазмолизированном состоянии. Эти условия тормозят метаболизм и регенерацию клеточной стенки. В качестве осмотических стабилизаторов используют сахара (глюкозу, маннит, сорбит, ксилозу), ионные осмотики (СаСl2, КСl) в концентрации 0,3–0,8 моль/литр. Концентрации подбираются индивидуально для каждого растительного объекта. Удобнее обрабатывать ткани ферментами в чашке Петри, которую держат под углом 15°. Смесь ферментов с протопластами переносят в центрифужные пробирки. Отделить протопласты от ферментативной смеси можно двумя способами: либо фильтрация с центрифугированием, либо флотация. При фильтрации смесь пропускают через фильтры с размерами пор 40 мкм. На фильтре при этом остаются комки клеток и их большие осколки. При дальнейшем центрифугировании оседают протопласты, осколки остаются в супернатанте. При повторном центрифугировании идет отмывка от фермента, после чего протопласты переносятся в среду для культивирования. Метод флотации предложен О. Гамборгом с сотрудниками в 1981 году, и предназначается для ослабленных протопластов. Он основан на том, что протопласты имеют более низкую плотность, чем органеллы или остатки клеточных стенок. К исходной смеси добавляют раствор сахарозы и центрифугируют при скорости от 40–80 до 350 д. Чистые протопласты плавают, осколки оседают на дно. Протопласты можно выделять также из суспензионных и клеточных культур. Лучше всего — в поздней стадии логарифмического роста, когда клеточные стенки легче поддаются разрушению, протопласты наиболее жизнеспособны. Далее протопласты культивируют в тех же условиях, что и клетки. Состав солей может быть несколько изменен. Среда состоит из осмотического стабилизатора, неорганических соединений, источника углерода, азота, витаминов, фитогормонов. Условия культивирования: pH среды 5,4–5,8, температура 22–28 °C, невысокая освещенность (не более 2000 лк).
Способы культивирования протопластов Существуют два способа культивирования протопластов: метод жидких капель и метод платирования. В первом случае суспензию протопластов в виде капель помещают на пластиковые чашки Петри. Вариацией этого способа является культивирование единичных изолированных протопластов в микрокаплях объемом 1 мкл, предложенное Ю. Глебой в 1978 г. Во втором — суспензию протопластов наливают в пластиковые чашки Петри, добавляют равный объем той же среды с 1 % агаром при температуре не выше 45 °C. После остывания чашки Петри переворачивают и культивируют при 28 °C. В данном случае протопласты фиксированы в одном положении и физически отделены друг от друга. Это дает возможность наблюдать за развитием интактного протопласта: формированием клеточной стенки, делением, ростом и развитием растения. Вариантом этой техники является использование кормящих протопластов или клеток, подвергнутых воздействию рентгеновского или γ-излучения, что блокирует их способность к делению. Такие протопласты или клетки смешивают с жизнеспособными протопластами и они поддерживают и стимулируют их рост. Сразу после удаления раствора фермента начинается образование клеточной стенки. Труднее добиться деления клеток и регенерации растений. Регенерация растений осуществляется либо через эмбриогенез, либо через развитие каллуса с дальнейшей индукцией морфогенеза. Добиваются этого добавлением в среду ауксинов или сочетания ауксинов с цитокининами. На пролиферацию клеток, возникших из протопластов, влияет 4 фактора: — видовая специфичность и физиологическое состояние исходной ткани растения, — способ и условия выделения протопластов, — плотность высева протопластов, — состав питательной среды.
Слияние протопластов — парасексуальная гибридизация Изолированные протопласты, еще не образовавшие клеточной стенки, могут сливаться между собой. Слияние протопластов — своеобразный метод гибридизации, так называемая парасексуальная, или соматическая гибридизация. В отличие от обычной, где сливаются половые клетки (гаметы), в качестве родительских при парасексуальной гибридизации используются диплоидные клетки растений. Внеядерные генетические детерминанты у большинства высших растений наследуются в половом процессе строго одноядерно и матерински. Техника парасексуальной гибридизации может позволить: — скрещивание филогенетически отдаленных видов растений (организмов), — получение асимметричных гибридов, несущих генный набор одного из родителей наряду с несколькими хромосомами, органеллами или цитоплазмой другого, — слияние трех и более клеток, — получение гибридов, представляющих сумму генотипов родителей, — перевод мутаций в гетерозиготное состояние, что позволяет получать жизнеспособные формы при слиянии протопластов, поскольку мутагенез довольно часто дает дефектное по морфогенезу растение, — получение растений, гетерозиготных по внеядерным генам и др. Парасексуальная гибридизация важна для анализа как ядерных генов, так и внеядерных геномов. Цитоплазматический геном кодирует ряд признаков — скорость фотосинтеза, устойчивость к патогенам, абиотическим факторам и т. д. Наличие косегрегация генов (признаки, контролирующие внеядерный геном, сегрегируют совместно) свидетельствует о физическом сцеплении генов. Слияние бывает спонтанным (чаще у протопластов из молодых тканей или суспензионных культур) и индуцированным. Для стимуляции слияния протопластов предложен ряд методов, как физических, так и химических. При физическом способе слияния протопластов, разработанном Г. Циммерманом с сотрудниками в 1981 году, протопласты помещают в камеру с неоднородным электрополем. На электродах образуются агрегаты из 2–3 протопластов, либо цепочки из 5–6 протопластов между электродами. Дополнительный единичный импульс постоянного тока приводит к образованию пор в сильно сжатых мембранах, происходит перетекание цитоплазмы, так как переменный ток удерживает протопласты вместе некоторое время, и протопласты в таких агрегатах сливаются. Затухающий ток приводит к возвращению сферической формы у слившихся протопластов. В основе слияния лежит различное действие постоянного и переменного электрического тока на плазмалемму. Постоянное эклектическое поле сжимает мембраны, ведя к их локальному разрушению, а переменное электрополе вызывает латеральную диффузию белков мембраны, образуя свободные от гликопротеидов липидные области, где противоположные мембраны могут установить контакт. Чаще для индукции слияния протопластов используют методику "ПЭГ — высокие значения pH — высокая концентрация Са2+", которая дает до 50 % слившихся протопластов (pH 9-11, концентрация Са2+ 100–300 ммоль/л). В присутствии полиэтиленгликоля наблюдается сильная адгезия протопластов, после удаления полиэтиленгликоля и добавления кальция — их слияние. Предполагают, что pH и ионы кальция увеличивают текучесть мембран, что связано с их жидкостномозаичной структурой. При слиянии протопластов различных растений, например, А и В, могут с равной вероятностью образовываться комбинации АА, ВВ и АВ. Желаемый продукт слияния — АВ, поэтому разрабатываются способы увеличения частоты слияния именно такого типа и избирательного выделения только продукта слияния АВ. Один из таких методов заключается в следующем. Поверхность протопласта обычно несет отрицательный заряд. Путем обработки ее фосфолипидом, несущим положительный заряд, можно временно придать поверхности протопласта положительный заряд. Если теперь протопласты А, имеющие положительный заряд, смешать с необработанными протопластами В, несущими отрицательный заряд, то будут в основном образовываться комбинации АВ в результате притяжения разноименных зарядов. Разработаны также методы маркирования протопластов того или иного растения с помощью разных флуоресцентных красителей. Если обработать протопласты одного растения флуоресцеинизотиоцианатом (FITC), а протопласты другого растения родаминизотиоцианатом (RITC), то можно, не изменяя активности клеток, пометить их желто-зеленой (FITC) или красной (RITC) флуоресценцией. Гибриды, образовавшиеся путем слияния разных типов клеток, будут иметь оба цвета флюоресценции — желто-зеленый и красный. Протопласты могут сливаться как попарно, так и в большем количестве. Многоядерные продукты слияния, как правило, разрушаются. Первое сообщение о получении соматических гибридов на уровне растений появилось в 1972 году (Карлсон и коллеги), в нашей стране подобное осуществили в лаборатории Бутенко Р.Г. в 1975 году. Судьба геномов (ядерного и цитоплазматического) после слияния протопластов может быть различной: 1. Ядерные генетические детерминанты наследуются как дву-, так и однородительски. В последнем случае ядра не сливаются и впоследствии сегрегируют в процессе клеточных делений. 2. Внеядерные генетические детерминанты наследуются двуродительски. При этом в межвидовых комбинациях прослеживается тенденция к соматическому выщеплению и элиминации одного из родительских цитоплазматических геномов. 3. Возникновение гибридных клеток и растений в результате слияния более чем двух родительских клеток. Таким образом, слияние протопластов приводит либо к образованию гибрида, либо к образованию цибрида. Соматический гибрид — продукт слияния и цитоплазмы, и ядра обоих протопластов. Цибрид (цитоплазматический гибрид) — растение-регенерант, содержащее цитоплазму обоих родителей и ядро одного из них. Цибриды получают, облучая перед слиянием один из протопластов у-лучами для разрушения ядра. Скрининг таких клеток проводится по генам — маркерам ядерного и цитоплазматических (митохондриального и хлоропластного) геномов. Есть указания на рекомбинацию ДНК митохондрий и хлоропластов в гибридных клетках (Ю.Ю. Глеба, К.М. Сытник, 1984). При слиянии могут образовываться и так называемые асимметричные гибриды — продукты слияния, имеющие полный хромосомный набор одного из партнеров и часть хромосом другого партнера. Такие гибриды часто возникают при слиянии клеток организмов, филогенетически удаленных друг от друга. В этом случае вследствие неправильных делений клетки, обусловленных некоординированным поведением двух разнородных наборов хромосом, в ряду поколений теряются частично или полностью хромосомы одного из родителей. Асимметричные гибриды бывают устойчивее, плодовитее и жизнеспособнее, чем симметричные, несущие полные наборы генов родительских клеток. В целях асимметричной гибридизации возможна избирательная обработка клеток одного из родителей для разрушения части его хромосом. Возможен прицельный перенос в клетку нужной хромосомы. Гибриды могут быть получены путем слияния трех и более родительских клеток. Из таких гибридных клеток могут быть выращены растения — регенеранты.
Виды соматических гибридов Впервые зрелый межвидовой гибрид, полученный в результате парасексуальной гибридизации протопластов 2 сортов табака (Nicotiana glauca, с 24 хромосомами и N.langsdorfii с 18 хромосомами), описан Карлсоном в 1972 г. Каллус амфиплоидного гибрида мог расти на безгормональной среде. Гибридное растение цвело. С тех пор были получены жизнеспособные внутривидовые, межвидовые, межродовые гибриды. Осуществлено слияние протопластов культурного картофеля сорта Приекульский ранний (Solanum tuberosum) с протопластами дикого картофеля (S. chacoense). Известно, что у дикого картофеля клубни очень мелкие. Вместе с тем, растение устойчиво ко многим заболеваниям. Картофель сорта Приекульский ранний образует крупные клубни, но растения этого сорта восприимчивы к болезням. Размеры протопластов у этих растений разные. Соматические гибриды по форме листьев и кустов, размеру клубней занимали промежуточное положение между культурными и дикими растениями. Вместе с тем гибрид, полученный в результате соматической гибридизации, оказался устойчивым к вирусу «У», чем отличался от полового гибрида. Первая попытка по созданию межродовых гибридов принадлежит Г. Мельхерсу, создавшему в 1978 году гибрид картофель + томат, так называемый томатофель. Гибрид был стерилен, морфологически аномален: толстые корни, отсутствие типичных столонов, махровые цветки. Было еще несколько попыток получения таких гибридов, но все растения стерильны. Эти эксперименты показали ограниченность применения парасексуальной гибридизации для прикладной селекции. Японскими исследователями (X. Кисака с соавт., 1997) путем электрослияния протопластов ячменя и риса был полученмежродовой соматический гибрид. Протопласты риса получали из суспензионной культуры, а протопласты ячменя были изолированы из молодых листьев. Часть полученных каллусов сформировали зеленые участки и побеги. Только один побег сформировал корни, и это растение было успешно перенесено в почву. По морфологии было близко к растениям риса. Цитологический анализ показал, что растение имело и маленькие хромосомы от риса, и большие от ячменя. Были проанализированы также митохондриальная и хлоропластная ДНК. Растение содержало новые последовательности и в митохондриальной, и в хлоропластной ДНК, которые не обнаруживались ни в одном из родителей. Ю. Ю. Глебой с сотрудниками проводились многочисленные эксперименты по созданию межтрибных гибридов. Триба — таксономическая единица между семейством и родом. Получены удачные гибриды между Arabidopsis и Brassica (турнепс) Arabidobrassica. У гибридных линий индуцировали морфогенез корней и растения. Растения генетически и морфологически униформны, не цвели. На вид — уродливы, очень много тератомоподобных образований, похожих на цветки. Была осуществлена гибридизация 2-х родов пасленовых Datura innoxia + Atropa belladonna. Удалось регенерировать растения. Во всех случаях выявлены хромосомы обоих родительских видов. Амфиплоиды оказались неспособны к стеблевому морфогенезу, в линиях с полиплоидным и анеуплоидным наборами хромосом получали аномальные стебли. Регенерировавшие растения были стерильны, похожи на дурман, но содержали небольшое количество хромосом красавки. В других экспериментах сливали протопласты красавки с каллусными клетками китайского табака. Получили 12 клонов. В клетках всех клонов обнаружили хромосомные типы обоих родителей, через год только у двух клонов происходила полная элиминация хромосом красавки. Морковь + сныть: из образовавшейся каллусной ткани через полгода регенерировали аномальные растения. Одно из них цвело, но у цветка отсутствовали пыльники и пестик. Интересные эксперименты были проведены в этой же лаборатории по гибридизации хлорофиллдефектного табака с красавкой. После слияния получили 40 фотосинтезирующих колоний, из них 4 клеточных линии дали нормальные растения красавки, 4 — аномальные по морфологии гибриды табак + красавка, остальные — зеленые, иногда пестролистные растения, идентичные табаку, которые цвели, давали семена. Они содержали хромосомы табака и пластиды красавки. Это были первые фертильные межтрибные гибриды. Первые работы по получению межсемейственных гибридов проведены К.Као и В.Веттером в 1976-77 гг. (соя + табак). Позднее в лаборатории Ю.Ю.Глебы провели аналогичные эксперименты пасленовые + бобовые и лилейные (горошек + табак и лук + табак). И.Ф.Каневскому удалось индуцировать морфогенез стеблеподобных тератом в культуре межсемейственных гибридов N.tabacum + Vicia faba. Практически во всех случаях наблюдалась видоспецифичная элиминация хромосом одного из родителей. В культурах межсемейственных гибридов наблюдалось много многоядерных клеток, клеток с мини ядрами, в метафазах делений встречались гигантские хромосомы. Отмечена асинхронность в расхождении родительских хромосом в анафазе. Морфогенез у такого материала отмечен не был. Для отдаленных гибридов характерно: 1. Относительная стабильность гибридного состояния, при котором не наблюдается полной элиминации генетического материала одного из родителей. 2. Генетические перестройки (реконструкция и частичная элиминация хромосом). 3. Генетическая разнокачественность клонов гибридных клеток. 4. Ограниченная морфогенетическая способность. Изучение межцарственных гибридов клеток "животное + растение[71]" показало, что на этапе слияния видоспецифичность не проявляется, поэтому можно слить даже животную и растительную клетки. На более поздних этапах онтогенеза эти различия сказываются, что было установлено в экспериментах по слиянию протопластов арабидопсиса и табака с лимфоцитами человека. При этом происходило слияние цитоплазмы, ядра не сливались. Эдвард Коккинг параллельно проводил изучение ультраструктуры таких гибридов, работая с клетками амфибий и протопластами моркови. После объединения клеток ядра амфибии были окружены тонким слоем собственной цитоплазмы, но уже через 48 часов отмечалось полное смешивание цитоплазмы и регенерация клеточной стенки вокруг гетерокариона.
КОНСТРУИРОВАНИЕ КЛЕТОК
Протопласты широко используются также в качестве реципиентов для клеточных органелл. В 1973 году И. Потрикусс и Ф. Хоффман успешно трансплантировали изолированные ядра петуньи в протопласты табака. Каким образом можно ввести ядро или другие клеточные органеллы в протопласт? При использовании сэндвич-метода трансплантацию проводят в условиях слабого деплазмолиза протопласта. Перед введением протопласты и ядра обрабатывают лизоцимом, который модифицирует клеточную мембрану. Далее осуществляют попеременное осаждение ядер и протопластов центрифугированием. В результате центрифугирования в пробирках формируется несколько чередующихся слоев. Центрифужные пробирки заполняют раствором осмотика (маннита) без лизоцима и центрифугируют полчаса при 140 д, оставляют на 2 часа при температуре +4 °C. Осадок ресуспендируют и просматривают под микроскопом. Ядра можно обнаружить и в цитоплазме, и в вакуолях. В некоторых случаях при поглощении ядра образуются жизнеспособные гибриды, в других — в клеточном цикле происходит потеря интеграции между чужеродным ядром и ядром хозяина. Клеточные органеллы можно также переносить посредством липосом. Широкий спектр одно- и многоламелльных частиц, которые сливаются с мембранами протопластов, получен с применением таких веществ, как фосфатидилсерин, холистерол и т. д. Можно также переносить органеллы путем микроинъекций. Этот метод широко используется для введения в клетку ДНК, РНК, а недавно был успешно использован и для переноса клеточных органелл у растений. Кроме ядра трансплантируют и другие органеллы, такие как митохондрии и хлоропласты. Выбор этих органелл объясняется их полуавтономностью, то есть наличием собственной ДНК и способность делиться самостоятельно, независимо от деления самой клетки. Кроме того, эти органеллы контролируют важнейшие физиологические процессы растительной клетки, такие как фотосинтез и дыхание. Например, перенос хлоропластов может использоваться для выведения новых форм хозяйственно важных сортов растений. Трансплантация высокоэффективных хлоропластов может способствовать активации фотосинтеза и повышению продуктивности растения. Одним из важных моментов является сохранение клеточных органелл, поэтому для переноса их в последнее время используются субпротопласты — фрагменты, полученные из протопластов. Они могут содержать большую часть цитоплазмы, но без ядра (цитопласты), ядро с небольшим количеством цитоплазмы (кариопласты), часть хромосом с небольшим количеством цитоплазматического материала (микропротопласты). Биологическое конструирование на уровне клетки может оказаться полезным и перспективным для создания клеток, клеточных систем и целых растений, удовлетворяющих потребности человека. Под биологическим конструированием следует понимать не только введение отдельных органелл (рис. 16). Аналогичным образом в клетку можно вводить и чужеродный генетический материал как в виде фрагментов ДНК, так и в виде отдельных хромосом. Кроме того, в изолированные протопласты можно вводить клетки микроорганизмов, создавая таким образом искусственные ассоциации.
Рис. 16. Конструирование клеток (по Р.Г. Бутенко, 1987)
КЛЕТОЧНАЯ СЕЛЕКЦИЯ
Методы клеточной селекции Одно из направлений клеточных технологий — это использование их в селекции, которое облегчает и ускоряет традиционный селекционный процесс в создании новых форм и сортов растений. Существующие методы культивирования изолированных клеток и тканей in vitro условно можно разделить на две группы. Первая группа — это вспомогательные технологии, которые не подменяют обычную селекцию, а служат ей. К ним можно отнести: оплодотворение in vitro (преодоление прогамной несовместимости), культивирование семяпочек и незрелых гибридных зародышей (преодоление постгамной несовместимости), получение гаплоидов путем культивирования пыльников и микроспор, криосохранение изолированных клеток, тканей и органов, клональное микроразмножение отдаленных гибридов. Вторая группа методов ведет к самостоятельному, независимому от традиционных методов селекции, получению новых форм и сортов растений: клеточная се лекция с использованием каллусной ткани, соматическая гибридизация (слияние изолированных протопластов и получение неполовых гибридов), применение методов генной инженерии. В отдаленной гибридизации находят применение такие методы культуры изолированных тканей, как оплодотворение in vitro, эмбриокультура (выращивание изолированных зародышей на искусственных питательных средах), клональное микроразмножение ценных гибридов, а также получение гаплоидов in vitro и криосохранение. Оплодотворение in vitro (преодоление прогамной несовместимости) проводится в том случае, когда невозможно осуществить оплодотворение между выбранными парами в естественных условиях. Это вызвано несколькими причинами: 1) физиологические (несоответствие во времени созревания пыльцы и т. д.); 2) морфологические (короткая пыльцевая трубка или блокирование роста ее на разных этапах развития и т. д.). Оплодотворение in vitro можно осуществить двумя способами: а) культивирование на искусственной агаризованной питательной среде завязи с нанесенной на нее готовой пыльцой; б) завязь вскрывается и на питательную среду переносятся кусочки плаценты с семяпочками, вблизи которых или непосредственно на ткани плаценты культивируется готовая пыльца. Визуально определить, прошло оплодотворение in vitro или нет, можно по быстро увеличивающимся в размерах семяпочкам. Сформировавшийся зародыш, как правило, не переходит в состояние покоя, а сразу прорастает и дает начало гибридному поколению. Плацентарное оплодотворение in vitro позволило преодолеть несовместимость в скрещивании сортов культурного табака N. tabacum с дикими видами N. rosulata и N. debneyi и сделало возможным получение межвидовых гибридов табака в опытах М.Ф. Терновского и др. (1976), Шинкаревой (1986). Постгамная несовместимость при отдаленной гибридизации возникает после оплодотворения. Часто при этом образуются щуплые невсхожие семена. Причиной может быть расхождение во времени развития зародыша и эндосперма. Из-за слабого развития эндосперма зародыш бывает неспособен к нормальному прорастанию. В таких случаях из зрелой щуплой зерновки изолируют зародыш и выращивают его в питательной среде. Выращивание зародышей в искусственной питательной среде называется эмбриокультурой. Среда для выращивания зрелого зародыша может быть простой, без добавок физиологически активных веществ (например, среда Уайта) или любая другая, содержащая минеральные соли и сахарозу. При более отдаленных скрещиваниях нарушения в развитии зародыша могут наблюдаться уже на ранних этапах, что выражается в отсутствии дифференцировки, замедленном росте. В этом случае культура зародыша состоит из двух этапов — эмбрионального роста зародыша, во время которого продолжается его дифференцировка, и прорастания подросшего зародыша. Для первого этапа требуется более сложная по составу среда с повышенным содержанием сахарозы, с добавками различных аминокислот, витаминов и гормонов. Применение эмбриокультуры в селекции приобретает в последнее время большое значение для получения отдаленных гибридов зерновых, злаковых и других сельскохозяйственных культур. Показана возможность увеличения выхода пшенично-ржаных гибридов путем доращивания незрелых зародышей, а также использования эмбриокультуры для преодоления постгамной несовместимости при гибридизации пшеницы с колосняком. Метод эмбриокультуры находит все более широкое применение в межвидовой гибридизации овощных растений. Для лука разработаны приемы выращивания in vitro абортивных зародышей от гибридных семян с разных этапов эмбриогенеза, выращивание зародышей от частично фертильных межвидовых гибридов. Культура изолированных зародышей используется в селекции томатов и других овощных растений. Исследована гормональная регуляция роста и развития зародышей томата in vitro. Обсуждается возможность применения эмбриокультуры для получения отдаленных гибридов подсолнечника, изучаются факторы, контролирующие рост и развитие in vitro зародышей подсолнечника, выделенных в разные сроки после опыления. Культура изолированных зародышей как вспомогательный метод при отдаленной гибридизации применяется не только для преодоления постгамной несовместимости, но также с целью микроразмножения ценных гибридов. В этом случае микроразмножение идет путем каллусогенеза, индукции морфогенеза и получения растений — регенерантов из каллусной ткани. Техника клонирования незрелых зародышей позволяет размножать ценные генотипы растений на ранних стадиях жизненного цикла. Еще одна возможность применения культуры зародышей — использование ее в клеточной селекции.
Генетические основы применения культуры клеток растений в селекционных целях Клетки в культуре in vitro отличаются по морфологии, по биохимическим свойствам, по физиологическому состоянию и генетически. Разнообразие (вариабельность) среди клеточных линий или растений-регенерантов называют сомаклональной вариабельностью. Генетическая природа и механизм возникновения сомаклональной изменчивости пока мало изучены. Однако четко можно выделить зависимость возникновения сомаклональных вариантов, прежде всего, от генетической гетерогенности соматических клеток исходного экспланта, генетической и эпигенетической изменчивости, индуцируемой условиями культивирования in vitro, а также от генотипа и исходного экспланта. Полиморфизм культивируемых клеток можно объяснить видовыми и возрастными особенностями, уровнем плоидности, влиянием состава питательной среды и условий культивирования, отсутствием коррелятивных связей. Последний фактор, ведущий к нарушению жесткой регуляции, существовавшей в целом растении, видимо, является основной причиной спонтанной изменчивости клеток in vitro. Любой фрагмент растения представляет собой мозаику различных тканей, и в зависимости от того, какая ткань даст начало каллусу, возникшие даже из одинаковых эксплантов каллусы будут гетерогенными и отличающимися друг от друга. Одинаковых, в полном смысле, эксплантов в природе быть не может, следовательно, неоднородность исходного материала (видовая, возрастная, физиологическая) предопределяет разнокачественность клеток в культуре. Физиологическая гетерогенность состоит в том, что клетки в популяции находятся в разном физиологическом состоянии, т. е. делятся, растут, стареют, погибают. Такая культура называется асинхронной. Заставить популяцию клеток высших растений проходить фазы клеточного цикла одновременно, т. е. синхронизировать их почти невозможно. Потому что та часть клеток, которая способна в данный момент к делению, составляет 2–4 %. Неблагоприятные условия (низкая температура, исключение важных компонентов питания), задерживающие деление, в какой-то степени способствуют накоплению числа клеток, готовых к делению. Более эффективны некоторые химические вещества, блокирующие определенные стадии подготовки к делению. В лучших случаях синхронизация может быть достигнута у 10–30 % клеток, но при последующих делениях популяция опять быстро утрачивает синхронность. Следует подчеркнуть, что физиологическая вариабельность клеток в суспензионной культуре меньше по сравнению с культурой каллусной ткани на агаре, что связано с более однородными условиями питания, аэрации и удаления токсических метаболитов из клеточного окружения в жидкой перемешиваемой среде. Гетерогенность культивируемых клеток обусловлена генетической, эпигенетической и модификационной изменчивостью. Генетические, или мутационные, изменения приводят к изменению генотипа, которое может быть унаследовано. Мутации (изменения количества или структуры ДНК) происходят на генном, хромосомном и геномном уровнях. Генная, или точечная, мутация означает изменение структуры ДНК в одном локусе. Генные мутации приводят к сильным или слабым изменениям морфологических, биохимических и физиологических свойств клетки. Мутации, возникающие в результате изменения макроструктуры хромосом, называются хромосомными мутациями, или хромосомными аберрациями (перестройками). Структурные перестройки хромосом возникают в результате инверсии, делеции, дупликации, транслокации и транспозиции. Геномные мутации связаны с изменением числа хромосом в ядре, т. е. с изменениями в кариотипе. Все виды названных генетических изменений имеют место у клеток in vitro. Наиболее подробно исследована хромосомная изменчивость клеток in vitro. Даже клетки одной и той же ткани, выращиваемые в одном сосу-де, могут значительно различаться между собой по хромосомным наборам (диплоидные, полиплоидные, анеуплоидные). Причины генетической изменчивости многообразны: 1) нарушение коррелятивных связей при выделении первичного экспланта из растения, т. е. отсутствие организменного контроля; 2) действие компонентов среды; 3) влияние продуктов метаболизма, накапливающихся в среде; 4) гетерогенность исходного материала и селекция клеток определенного типа. Хромосомная изменчивость является результатом нарушений митоза, называемых эндомитозом и эндоредупликацией. При эндомитозе происходит спирализация хромосом и начинается митоз, но нарушается веретено деления, сохраняется оболочка ядра, хромосомы не расходятся и деспирализуются внутри ядерной оболочки. Это приводит к возрастанию числа хромосом, увеличению размеров ядра и клеток. Эндоредупликация не сопровождается образованием хромосом и делением ядра, хотя содержание ДНК в ядре тоже увеличивается. К образованию полиплоидных и анеуплоидных клеток также приводят нарушения в митозе, связанные с неправильным распределением хромосом. Клетки различного уровня плоидности различаются по скорости деления и роста, по устойчивости к неблагоприятным воздействиям, начинают конкурировать, и одни из них начинают преобладать. Такой процесс возрастающего доминирования в популяции клеток определенного типа называется клеточной селекцией. Доминирование может быть вызвано преимущественной пролиферацией одних клеток или успешной элиминацией (удалением) других. Такую селекцию правильнее называть автоселекцией, потому что она протекает спонтанно, без специального воздействия какими-либо стрессовыми факторами. В процессе автоселекции формируется наиболее приспособленный к данным условиям кариотип. Вероятно, клетки приспосабливаются к новым условиям существования путем отбора более жизнеспособных полиплоидных клеток. Интересно, что изменение условий выращивания меняет направление отбора. Показано, например, что высокие концентрации 2,4-Д и кинетина увеличивают возможность полиплоидизации. То, что условия выращивания играют важную роль в формировании цитогенетической гетерогенности, хорошо видно из опытов с тканью гаплопаппуса. В лаборатории Р. Г. Бутенко в течение двух лет культивировались меристематические клетки этого растения, пассажи проводились раз в месяц. В итоге исходные диплоидные клетки на 95 % приобрели другие уровни плоидности. Шведский исследователь Т. Эриксон, работая с этой же тканью, пересаживал ее на свежую питательную среду через каждые 2 дня. При этом штамм сохранил стабильную диплоидную характеристику. Однако способ выращивания не может полностью гарантировать генетическую стабильность в популяции клеток, так как генетической гетерогенностью может обладать сам исходный материал. У многих растений дифференцированные ткани имеют клетки разной плоидности. Специализированные клетки, например клетки зеленой ассимилирующей паренхимы листа, запасающих тканей мясистых корней, клубней, зачастую являются полиплоидными. Спонтанное или индуцированное каким-либо фактором образование различных вариантных форм растений можно использовать для улучшения уже существующих сортов сельскохозяйственных культур. Как было отмечено, клетки in vitro становятся разнокачественными также благодаря эпигенетическим изменениям, т. е. изменениям в программе считки генов или потенции к их активации. Эти изменения генной активности являются наследуемыми. К ненаследуемым изменениям у клеток в культуре относятся модификационные изменения, которые в большинстве носят адаптивный, приспособительный характер. Эти изменения не затрагивают генетических структур клетки, они соответствуют физиологической адаптации, при которой границы изменений не превышают норму реакции, обусловленной генотипом. Гетерогенность клеток in vitro возрастает с увеличением продолжительности их культивирования. Различные типы морфогенеза, соматический эмбриогенез или органогенез, также могут по-разному сказываться на генетических изменениях и, соответственно, на фенотипе растений. Экспериментально установлено, что при соматическом эмбриогенезе время прохождения цикла клетка — растение значительно короче, чем при органогенезе, поэтому степень сходства получаемого материала и исходного родительского генотипа может быть значительно выше. Сомаклональные варианты имеют, несомненно, практическое применение в сельскохозяйственной практике, в силу появления форм, отличающихся от родительских по различным биохимическим, качественным и количественным признакам, а также цитогенетическим характеристикам. Например, получены сомаклоны картофеля сорта Зарево, отличающиеся высокой урожайностью, повышенной устойчивостью к заболеваниям, более высоким содержанием в клубнях протеина и крахмала. Причем наследование важных признаков при размножении клубнями сохранялось в течение трех лет полевых испытаний (В.В. Сидоров и др., 1984, 1985). Для растений табака получены через каллусную культуру сомаклоны, устойчивые к вирусу табачной мозаики. В настоящее время метод культуры тканей начал широко использоваться в селекции не только кормовых и технических культур, но и декоративных и лекарственных растений. Примером тому может служить новый сорт пеларгонии Velvet Rose, полученный через каллусную культуру. Таким образом, полученные положительные результаты свидетельствуют о необходимости более эффективного внедрения различных приемов получения сомаклональных вариантов в практику селекционной работы, и наиболее реальным является применение сомаклональной изменчивости для улучшения или «доработки» уже существующих сортов или линий по отдельным недостающим признакам. Несмотря на то, что существует генетическая нестабильность культур изолированных тканей и клеток растений, она не может обеспечить потребности селекционеров в генетическом разнообразии. В связи с этим для ускорения селекционного процесса в культуре клеток используются химические и физические мутагены. Обработка ткани раувольфии змеиной азотистым ипритом в концентрации 2,5∙10-3 М привела к повышению уровня аберраций хромосом в первом пассаже до 32 %, вызвала сдвиг популяции в сторону увеличения триплоидов. В результате удалось получить штамм с более высокой биосинтетической активностью по сравнению с исходной тканью. Спонтанный и индуцированный мутагенез в культуре клеток, тканей и протопластов позволяет получать растения, представляющие практический интерес для селекционеров. Важное практическое значение имеет создание форм растений, устойчивых к неблагоприятному действию факторов внешней среды, таких как низкие температуры, засоление почв, загрязнение природной среды токсическими веществами, поражение вредителями и возбудителями болезней. Эти факторы могут быть использованы в качестве селективного фона в процессе клеточной селекции. Клетки, сохранившие при этом жизнеспособность, могут быть регенерированы в целые растения. Принципиальной разницы в результатах клеточной селекции при спонтанном и индуцированном мутагенезе нет. Для повышения частоты мутаций в соматических клетках обычно используют такие мутагены, как нитрозогуанидин, нитрозометилмочевина, метилметансульфонат. Реже применяют облучение ультрафиолетом, γ-квантами 60Со и нейтронами. В качестве селективных агентов используют антибиотики, ингибиторы синтеза нуклеиновых кислот, аналоги пуриновых и пиримидиновых оснований, фито- и патотоксины, агенты, вызывающие солевой и водный стресс, аналоги аминокислот и Для проведения клеточной селекции используют следующие приемы: — прямая (позитивная) селекция, при которой выживает лишь определенный искомый мутантный тип клеток; — непрямая (негативная) селекция, основанная на избирательной гибели делящихся клеток дикого типа и выживания метаболически неактивных клеток, но требующая дополнительной идентификации у них мутационных изменений; — тотальная селекция, при которой индивидуально тестируются все клеточные клоны; — визуальная селекция и неселективный отбор, когда вариантная линия может быть идентифицирована среди всей популяции клеток визуально или при использовании биохимических методов (тонкослойная или жидкостная хроматография, радиоиммунный анализ, микроспектрофотометрия и др.). Из перечисленных выше приемов клеточной селекции прямая селекция является наиболее распространенным методом и используется главным образом для выделения растений-регенерантов, устойчивых, например, к гербицидам, антибиотикам, токсинам, тяжелым металлам, солям и другим антиметаболитам.
Виды культур растительных клеток, используемые в клеточной селекции Для проведения работ по клеточной селекции растений в условиях in vitro в качестве объекта исследования могут быть использованы каллусные, суспензионные культуры или изолированные протопласты. Выбор объекта зависит от наличия разработанных технологий применительно к различным видам растений, а также от конечных целей исследования. Каллусная ткань представляет собой легко доступный материал, который наиболее часто используют для клеточной селекции. Как правило, работу проводят на первичной или пересадочной каллусной ткани, которая не утрачивает способности к регенерации на протяжении ряда субкультивирований. Однако при работе с каллусными культурами многие исследователи отмечают существенные недостатки данного объекта: медленный рост ткани, неравноценное для всех клеток действие токсических веществ, которые применяются в качестве селективного фактора, а также потеря регенерационной способности в процессе культивирования каллусных клеток. Несомненно, проводить селекцию целесообразно на уровне одиночных клеток (суспензионная культура, протопласты). Однако для многих видов растений не разработаны эффективные технологии и способы культивирования одиночных клеток. Поэтому, несмотря на перечисленные выше недостатки использования каллусных культур, этот способ селекции остается для некоторых видов растений пока единственным. Получение стабильно устойчивых линий — процесс длительный. Как правило, селекция начинается с получения достаточного количества каллусной массы из изолированных растительных эксплантов, использующейся в дальнейшем для определения концентрации селективного фактора (построение дозовой кривой), при которой наблюдается одновременно рост каллусной ткани, и в то же время часть каллусных колоний погибает. Выбранная концентрация селективного фактора признается оптимальной и используется в дальнейших экспериментах. Так как первично полученные на средах с селективными факторами колонии клеток могли возникнуть вследствие физиологической адаптации или определенного состояния дифференцировки клеток и не быть генетически устойчивыми, то в течение последующих 4–6 субкультивирований на селективной среде проверяется стабильность устойчивости полученных клонов. Затем их переносят на среду без селективного фактора и субкультивируют еще 2–3 пассажа. И только после повторного возвращения в селективные условия отбирают стабильные клоны, из которых пытаются получить растения-регенеранты. Однако работы, проведенные с получением растений, устойчивых к повышенным солям, а также к токсинам, выделенным из грибов — возбудителей болезней, показали, что устойчивость клетки и растения к исследуемому селективному фактору может совпадать и не совпадать. Прямая корреляция между устойчивостью растений и клеток in vitro отмечена лишь для низких температур, устойчивостью к гербицидам, высоким концентрациям алюминия и другим факторам. Большое число работ по культивированию каллуса, с целью получения нового селекционного материала, проведено на пшенице, ячмене, рисе, сорго, а также на картофеле, томатах, люцерне и, крайне редко, на древесных. Уже достигнуты первые положительные результаты по получению растений пшеницы, риса, картофеля, устойчивых к NaCl или Na2SO4. Получены клетки, а из них растения моркови, которые синтезируют в 20 раз больше метионина, в 30 раз — триптофана, в 5 раз — лизина путем добавления в питательную среду токсичных аналогов аминокислот. Для картофеля получены растения, устойчивые к кольцевой гнили. Что касается древесных, то для них работы в этом направлении крайне редки и часто имеют поисковый характер. Таким образом, использование каллусной культуры в селекционных целях открывает огромные возможности в создании новых форм растений, несущих ценные признаки, необходимые для человечества. Наряду с перечисленными выше объектами (каллусная, суспензионная культура, изолированные протопласты), в качестве исходного материала для селекции могут быть использованы культуры соматических или андрогенных эмбриоидов, такие органогенные экспланты, как сегменты листьев или различные меристематические и стеблевые части растений, а также культура изолированных зародышей. Например, путем культивирования и селекции in vitro зародышей из семян получены растения ячменя, устойчивые к аналогам аминокислот, с улучшенным содержанием белка. Для картофеля разработан эффективный метод обработки побегов и черенков мутагеном, приводивший к получению химерных мутантов хлорофиллдефектности и антибиотикоустойчивости. При культивировании пыльников яровой пшеницы сорта Саратовская-29 и Московская-35 на питательных средах с повышенным содержанием солей хлорида натрия получены соматические эмбриоиды, а в дальнейшем растения-регенеранты, проявившие повышенную солеустойчивость (Беккужина, 1993). Таким образом, проведение селекции на клеточном уровне позволяет создавать новые формы растений в 2–4 раза быстрее по сравнению с традиционными способами селекции.
Преимущества метода клеточной селекции in vitro По сравнению с экспериментальным мутагенезом на уровне целых растений метод мутагенеза на уровне клеток имеет ряд преимуществ: — экономится площадь, так как в одной чашке Петри диаметром 10 см можно культивировать 107-108 клеток, а для такого же количества растений необходима площадь свыше тысячи гектаров; — мутантные признаки на уровне отдельных клеток проявляются довольно быстро; — возможно получение новых типов мутаций, в том числе и биохимического характера; — экономится время и трудозатраты на получение нового желаемого признака. Основным требованием для успешного использования клеточного мутагенеза является хорошо разработанная система регенерации растений. Важным условием является также возможность получения гаплоидов у того или иного вида растений. В дальнейшую селекционную работу включаются только те генотипы, у которых мутации проявляются на уровне целого растения. Растения с измененными признаками, полученные в результате мутагенеза на клеточном уровне, называются вариантами (термин «мутант» используется тогда, когда мутация подтверждается генетическими или молекулярно-генетическими методами). Рекомендуются следующие обозначения: R0 — растения-регенеранты, полученные из соответствующих клеточных клонов, R1, R2 и т. д. — первое и последующее поколения после самоопыления. Общая схема получения мутантных форм путем селекции на клеточном уровне состоит из нескольких этапов (рис. 17):
Рис. 17. Схема получения мутантных форм путем клеточной селекции (по В.И. Артамонову, 1989)
Измененные при мутагенной обработке клетки могут быть выделены в условиях культивирования in vitro путем прямого и непрямого отборов, а также при тестировании отдельных клеточных колоний. Прямой отбор состоит в добавлении к питательным средам отдельных компонентов, к которым обычные, неизмененные клетки не устойчивы. Непрямой отбор (негативная селекция) заключается в создании условий культивирования, при которых рост неизмененных клеток либо задерживается, либо эти клетки погибают (например, культивирование при низких или высоких температурах на средах с недостатком отдельных компонентов и т. д.). Существует ряд факторов, ограничивающих селекцию in vitro. Многие хозяйственно важные признаки, такие, как урожайность, количество зерна, устойчивость к пестицидам и другие трудно или практически невозможно получить при культивировании in vitro поскольку они не проявляются на клеточном уровне. Недостаточно также биохимических и молекулярных маркеров, которые коррелировали бы с этими признаками на уровне целых растений. Не все селектируемые признаки, проявляющиеся на уровне клеток, сохраняются на уровне растений — регенерантов. Тому несколько причин: некоторая часть изменений не затрагивает генетический аппарат клетки, поэтому не сохраняется у потомков; генетические изменения могут элиминироваться в процессе дифференциации и мейоза; функция мутированного гена может быть ограничена состоянием дифференцируемых и культивируемых клеток; мутация одного гена может сопровождаться активацией различных генов, кодирующих изоферменты; часть генотипов неспособна регенерировать нормальные фертильные растения.
КЛОНАЛЬНОЕ МИКРОРАЗМНОЖЕНИЕ И ОЗДОРОВЛЕНИЕ РАСТЕНИЙ
Преимущества микроклонального размножения перед традиционными способами размножения растений. История метода. В природе существует два способа размножения растений: половой (семенной) и вегетативный. Оба эти способа имеют как свои преимущества, так и недостатки. К недостаткам семенного размножения относятся генетическая пестрота семенного материала и длительность ювенильного периода. При вегетативном размножении генотип материнского растения сохраняется, а также сокращается длительность ювенильного периода. Однако большинство видов плохо размножается вегетативным способом, к ним относятся многие древесные породы. Например, эффективность размножения, даже на ювенильной стадии, дуба, сосны, ели, орехоплодных не очень высока. Кроме того, с помощью черенкования невозможно размножать многие виды древесных растений в возрасте старше 10–15 лет. Трудно получить стандартный посадочный материал, так как существует возможность накопления и передачи инфекции. Операции по размножению с помощью прививок сложны и трудоемки. Достижения в области культуры клеток и тканей привели к созданию принципиально нового метода вегетативного размножения — клонального микроразмножения. Клональное микроразмножение — получение in vitro, неполовым путем, генетически идентичных исходному экземпляру растений. В основе метода лежит уникальная способность растительной клетки реализовывать присущую ей тотипотентность. Термин "клон" был предложен в 1903 году Уэбстером (от греческого klon — черенок или побег, пригодный для размножения растений). В соответствии с научной терминологией клонирование подразумевает получение идентичных организмов из единичных клеток. Этот метод имеет ряд преимуществ перед существующими традиционными способами размножения: — получение генетически однородного посадочного материала; — освобождение растений от вирусов за счет использования меристемной культуры; — высокий коэффициент размножения (105-106 — для травянистых, цветочных растений, 104-105 — для кустарниковых древесных растений и 104 — для хвойных); — сокращение продолжительности селекционного процесса; — ускорение перехода растений от ювенильной к репродуктивной фазе развития; — размножение растений, трудно размножаемых традиционными способами; — возможность проведения работ в течение всего года; — возможность автоматизации процесса выращивания. Пионером клонального микроразмножения считается французский ученый Жан Морель, который в 50-х годах нашего столетия получил первые растения — регенеранты орхидей. В это время техника культивирования апикальных меристем in vitro была уже хорошо разработана. Как правило, исследователи в качестве первичного экспланта использовали верхушечные меристемы травянистых растений: гвоздики, хризантемы, подсолнечника, гороха, кукурузы и т. д. В нашей стране работы по клональному микроразмножению были начаты в 30-х годах в лаборатории культуры тканей и морфогенеза ИФРа. Под руководством Р.Г.Бутенко были изучены условия микроразмножения картофеля, сахарной свеклы, гвоздики, герберы и др. растений и предложены промышленные технологии. В дальнейшем исследования по клональному микроразмножении охватили и древесные растения. Первые работы по культуре тканей древесных растений были опубликованы в середине 20-х годов нашего столетия и связаны с именем Готре, который показал, что камбиальные ткани некоторых растений способны к каллусогенезу in vitro. Но первые растения — регенеранты осины, доведенные до почвенной культуры, были получены лишь в середине 60-х годов Матесом. Культивирование тканей хвойных пород in vitro долгое время редко использовалось как объект исследования. Это было связано со специфическими трудностями культивирования тканей, изолированных из растения. Известно, что древесные, и особенно хвойные растения характеризуются медленным ростом, трудно укореняются, содержат большое количество вторичных соединений (фенолы, терпены и т. д.), которые в изолированных тканях активируются. Окисленные фенолы обычно ингибируют деление и рост клеток, что ведет к гибели первичного экспланта или уменьшению способности тканей древесных растений к регенерации адвентивных почек, которая с возрастом растения-донора исчезает практически полностью. В настоящее время, несмотря на перечисленные трудности, насчитывается более 200 видов древесных растений из 40 семейств, которые были размножены in vitro (каштан, дуб, береза, клен, сосна, ель, секвойя и др.).
Факторы, влияющие на процесс клонального микроразмножения На эффективность микроклонального размножения влияет масса факторов различной природы. Это физиологические особенности вводимого в культуру растения, химические и физические условия культивирования. Наиболее важным моментом является выбор материнского растения и экспланта. При выборе материнского растения необходимо учитывать его физиологические, сортовые и видовые особенности. Исходные растения должны быть здоровы, не поражены грибными, бактериальными и вирусными болезнями. Кроме того, они должны находится в состоянии интенсивного роста (выход из фазы покоя и переход к активному росту). Луковицы, корневища и клубни в состоянии покоя непригодны, перед введением в культуру их предварительно обрабатывают высокими или низкими температурами. Способность к размножению также детерминирована генетически. Например, земляника размножается всеми способами, облепиха — ни одним, хотя в природе черенкуется. Двудольные обладают большей регенерационной способностью, чем однодольные и древесные. При выборе экспланта необходимо учитывать его возраст, строение и происхождение. Для обеспечения максимальной стабильности клонируемого материала, во избежание появления аномальных растений в качестве экспланта желательно использовать молодые, слабодифференцированные ткани. Кроме того, экспланты от ювенильных растений лучше укореняются, чем от зрелых, особенно это касается древесных пород. Лучше всего использовать кончики стеблей, пазушные почки, зародыши, молодые листья, черенки, соцветия, чешую луковиц, то есть экспланты, содержащие меристемы. Опыты с эмбрионами кукурузы, проведенные Грином и Филипсом в 1975 году, показали, что при извлечении эмбрионов из зрелых семян они образуют каллус и корни. Если же изолировать их через 2–3 недели после опыления, то образуются и каллус, и растения. Вероятно, это связано с разворачиванием генетической программы в онтогенезе растения. Следует отметить, что не всегда молодые ткани являются удачным объектом для размножения. У эхеверии на эксплантах из молодых листьев возникают только корни, из старых — только побеги, из средних по возрасту — и побеги, и корни. Чем меньше размер экспланта, тем меньше его регенерационная способность. С другой стороны, в крупном экспланте увеличивается возможность появления в его клетках вирусов и других патогенов, что препятствует оздоровлению тканей. Длительность культивирования также влияет на эффективность микроразмножения. Физиологическое состояние экспланта меняется в течение пассажей, при длительном культивировании частота укореняемости побегов возрастает. Возможно, что при этом эксплант приобретает признаки ювенильности, что ведет к повышению его морфогенетического потенциала. Успех введения в культуру часто определяется эффективностью стерилизации. Выбор стерилизующего агента определяется особенностями экспланта. Для нежных тканей концентрация стерилизующего агента должна быть снижена, чтобы сохранить жизнеспособность экспланта. Часто внутреннее заражение исходных эксплантов бывает намного сильнее, чем поверхностное, поэтому экспланты предварительнообрабатывают фунгицидами и антибиотиками против грибной и бактериальной инфекций. Хорошие результаты дает обработка растений бензоатом натрия. В зависимости от вида растений необходимо испытывать как твердые, так и жидкие питательные среды. Иногда жидкие среды имеют преимущество, так как обеспечивают большую подвижность трофических элементов. Например, при размножении роз более успешным было культивирование побегов на двухслойной питательной среде: нижний слой — агаризованный, верхний — жидкий. На эффективность размножения могут также влиять расположение экспланта (горизонтальное или вертикальное), тип пробок (ватные, пластмассовые, стеклянные, металлические и т. д.), а также соотношение объема эксплантов и количества питательной среды для оптимального освещения и газообмена эксплантов. Состав питательной среды необходимо подбирать для каждого вида растений. На клональное микроразмножение влияют гормоны, минеральные соли, витамины и углеводы. При микроразмножении in vitro часто используют среды Мурасиге и Скуга, Линсмайера и Скуга, Шенка и Хильдебрандта, Нича, Гамборга, Хеллера и другие. Обычно используют среду Мурасиге-Скуга, которая содержит много неорганического азота, что стимулирует процессы органогенеза и соматического эмбриогенеза. В наших экспериментах (Кузьмина Н.А., Внукова В.В., 1997) выход морфогенных каллусов твердой пшеницы был выше на среде Мурасиге-Скуга по сравнению со средой Гамборга, которая одержала окисленные формы азота. Среда Мурасиге-Скуга также способствовала стабилизации хромосомного набора клеток твердой пшеницы при высоком содержании ауксина в среде. Вообще вопрос оптимального соотношения NH4/NO3 остается открытым, так как литературные данные весьма противоречивы и универсального рецепта для всех видов растений нет. В качестве источника углеродного питания используют различные углеводы типа сахарозы, глюкозы, фруктозы, галактозы. Разные культуры требуют различной концентрации углеводов на разных этапах клонального микроразмножения. К физическим факторам выращивания относятся температура и условия освещения. На первых двух этапах освещенность колеблется от 1000 до 3000 Лк, фотопериод 14–16 часов, но эти параметры зависят от культуры. Высокая интенсивность света может вызывать хлорозы и задерживать развитие, но при переносе в почву эти растения чувствуют себя лучше и растут энергичнее. Спектральный состав также играет немаловажную роль. Некоторые исследователи (Катаева Н.В., Аветисов В.А, 1981) указывают на синий свет как основной компонент морфогене за. Красный свет стимулирует образование почек у табака, у салата — образование побегов, у березы — укоренение. В работах Т.Н. Константиновой с соавторами (1987) показано, что синий свет усиливает закладку вегетативных почек у побегов табака в условиях in vitro, а красный стимулирует развитие цветочных почек. Однако при добавлении цитокининов и ауксинов в различной концентрации соотношение процессов дифференциации цветочных и вегетативных почек меняется, в некоторых случаях наблюдается даже противоположный эффект. В исследованиях Р. А. Карначук и Е. С. Гвоздевой (1998) наибольший выход морфогенных каллусов пшеницы, формирующих растения и побеги, отмечен на зеленом свету. Важное значение играет также сочетание спектрального состава света и гормональных факторов среды. Температура культивирования обычно варьирует в интервале 22–26 °C днем и 18–22 °C ночью. В некоторых случаях понижение температуры ведет к повышению эффективности размножения. Для повышения коэффициента размножения необходимо каждому виду с учетом его естественного ареала произрастания подбирать индивидуальные условия культивирования. Относительная влажность воздуха — 65–70%
Этапы клонального микроразмножения Процесс клонального микроразмножения можно разделить на 4 этапа: 1. Выбор растения-донора, изолирование эксплантов и получение хорошо растущей стерильной культуры. 2. Собственно микроразмножение, когда достигается получение максимального количества меристематических клонов. 3. Укоренение размноженных побегов с последующей адаптацией их к почвенным условиям, а при необходимости депонирование растений-регенерантов при пониженной температуре (+2 °C, +10 °C). 4. Выращивание растений в условиях теплицы и подготовка их к реализации или посадке в поле. Для культивирования тканей на каждом из четырех этапов требуется применение определенного состава питательной среды. На первом этапе необходимо добиться получения хорошо растущей стерильной культуры. В тех случаях, когда трудно получить исходную стерильную культуру экспланта, рекомендуется вводить в состав питательной среды антибиотики (тетрациклин, бензилпенициллин и др.) в концентрации 100–200 мг/л. Это в первую очередь относится к древесным растениям, у которых наблюдается тенденция к накоплению внутренней инфекции. На первом этапе, как правило, используют среду, содержащую минеральные соли по рецепту Мурасига и Скуга, а также различные биологически активные вещества и стимуляторы роста (ауксины, цитокинины) в различных сочетаниях в зависимости от объекта. В тех случаях, когда наблюдается ингибирование роста первичного экспланта, за счет выделения им в питательную среду токсичных веществ (фенолов, терпенов и других вторичных соединений), снять его можно, используя антиоксиданты. Это возможно двумя способами: либо омывкой экспланта слабым его раствором в течение 4-24 ч, либо непосредственным добавлением в питательную среду. В качестве антиоксидантов используют: аскорбиновую кислоту (1 мг/л), глютатион (4–5 мг/л), дитиотриэтол (1–3 мг/л), диэтилдитиокарбомат (2–5 мг/л), поливинилпирролидон (5000-10000 мг/л). В некоторых случаях целесообразно добавлять в питательную среду адсорбент — древесный активированный уголь в концентрации 0,5–1 %. Продолжительность первого этапа может колебаться от 1 до 2 месяцев, в результате которого наблюдается рост меристематических тканей и формирование первичных побегов. 2 этап — собственно микроразмножение. На этом этапе необходимо добиться получения максимального количества мериклонов, учитывая при этом, что с увеличением субкультивирований увеличивается число растений-регенерантов с ненормальной морфологией и возможно наблюдать образование растений-мутантов. Как и на первом этапе, используют питательную среду по рецепту Мурасига и Скуга, содержащую различные биологически активные вещества, а также регуляторы роста. Основную роль при подборе оптимальных условий культивирования эксплантов играют соотношение и концентрация внесенных в питательную среду цитокининов и ауксинов. Из цитокининов наиболее часто используют БАЛ в концентрациях от 1 до 10 мг/л, а из ауксинов — ИУК и НУК в концентрациях до 0,5 мг/л. При долгом культивировании растительных тканей на питательных средах с повышенным содержанием цитокининов (5-10 мг/л) происходит постепенное накопление их в тканях выше необходимого физиологического уровня, что приводит к появлению токсического действия и формированию растений с измененной морфологией. Вместе с тем, возможно наблюдать такие нежелательные для клонального микроразмножения эффекты, как подавление пролиферации пазушных меристем, образование витрифицированных (оводненных) побегов и уменьшение способности растений к укоренению. Отрицательное действие цитокининов возможно преодолеть, по данным Н.В. Катаевой и Р.Г. Бутенко, путем использования питательных сред с минимальной концентрацией цитокининов, обеспечивающих стабильный коэффициент микроразмножения, или путем чередования циклов культивирования на средах с низким и высоким уровнем фитогормонов. 3 и 4 этапы — укоренение микропобегов, их последующая адаптация к почвенным условиям и высадка в поле являются наиболее трудоемкими этапами, от которых зависит успех клонального микроразмножения. На третьем этапе, как правило, меняют основной состав среды: уменьшают в два, а иногда и в четыре раза концентрацию минеральных солей по рецепту Мурасига и Скуга или заменяют ее средой Уайта, уменьшают количество сахара до 0,5–1 % и полностью исключают цитокинины, оставляя один лишь ауксин. В качестве стимулятора корнеобразования используют β-индолил-3-масляную кислоту (ИМК), ИУК или НУК. Укоренение микропобегов проводят двумя способами: 1) выдерживание микропобегов в течение нескольких часов (2-24 ч) в стерильном концентрированном растворе ауксина (20–50 мг/л) и последующее их культивирование на агаризованной среде без гормонов или непосредственно в подходящем почвенном субстрате (импульсная обработка); 2) непосредственное культивирование микропобегов в течение 3–4 недель на питательной среде, содержащей ауксин в невысоких концентрациях (1–5 мг/л в зависимости от исследуемого объекта). В последнее время предложен метод укоренения пробирочных растений в условиях гидропоники. Этот метод позволяет значительно упростить этап укоренения и одновременно получать растения, адаптированные к естественным условиям. Для картофеля возможно использовать безсубстратную гидропонику для получения мини-клубней. Затенение нижней части культуральных сосудов плотной черной материей или добавление в питательную среду активированного угля способствует укоренению микропобегов. Пересадка растений-регенерантов в субстрат является ответственным этапом, завершающим процесс клонального микроразмножения. Наиболее благоприятное время для пересадки пробирочных растений — весна или начало лета. Растения с двумя-тремя листьями и хорошо развитой корневой системой осторожно вынимают из колб или пробирок пинцетом с длинными концами или специальным крючком. Корни отмывают от остатков агара и высаживают в почвенный субстрат, предварительно простерилизованный при 85–90 °C в течение 1–2 ч. Для большинства растений в качестве субстратов используют торф, песок (3:1); торф, дерновую почву, перлит (1:1:1); торф, песок, перлит (1:1:1). Исключение составляют семейство орхидных, для которых готовят субстрат, состоящий из сфагнового мха, смеси торфа, листьев бука или дуба, сосновой коры (1:1:1). Приготовленным заранее почвенным субстратом заполняют пикировочные ящики или торфяные горшочки, в которых выращивают растения-регенеранты. Горшочки с растениями помещают в теплицы с регулируемым температурным режимом (20–22 °C), освещенностью не более 5 тыс. лк и влажностью 65–90 %. Для лучшего роста растений создают условия искусственного тумана. В тех случаях, когда нет возможности создать такие условия, горшочки с растениями накрывают стеклянными банками или полиэтиленовыми пакетами, которые постепенно открывают до полной адаптации растений. Через 20–30 дней после посадки хорошо укоренившиеся растения подкармливают растворами минеральных солей Кнудсона, Мурасига и Скуга, Чеснокова, Кнопа (в зависимости от вида растений) или комплексным минеральным удобрением. По мере роста растений их рассаживают в большие емкости со свежим субстратом. Дальнейшее выращивание акклиматизированных растений соответствует принятой агротехнике выращивания для каждого индивидуального вида растений. Процесс адаптации пробирочных растений к почвенным условиям является наиболее дорогостоящей и трудоемкой операцией. Нередко после пересадки растений в почву наблюдается остановка в росте, опадение листьев и гибель растений. Эти явления связаны, в первую очередь, с тем, что у пробирочных растений нарушена деятельность устьичного аппарата, вследствие чего происходит потеря большого количества воды. Во-вторых, у некоторых растений в условиях in vitro не происходит образования корневых волосков, что приводит, в свою очередь, к нарушению поглощения воды и минеральных солей из почвы. Поэтому целесообразно на третьем или четвертом этапах клонального микроразмножения применять искусственную микоризацию растений (для микотрофных), учитывая их положительную роль в снабжении растений минеральными и органическими питательными веществами, водой, биологически активными веществами, а также в защите растений от патогенов. Индийскими учеными предложен простой метод предотвращения быстрого обезвоживания листьев растений, выращенных in vitro, во время их пересадки в полевые условия. Метод заключается в том, что листья в течение всего акклиматизационного периода следует опрыскивать 50 %-ным водным раствором глицерина или смесью парафина, или жира в диэтиловом эфире (1:1). Применение этого метода помогает избежать длинных и затруднительных процессов закаливания пробирочных растений и обеспечивает 100 %-ную их приживаемость.
Методы клонального микроразмножения Существует много методов клонального микроразмножения, а также различных их классификаций. Согласно одной из них, предложенной Мурасиге в 1977 году, процесс можно осуществлять следующими путями: 1. Активация пазушных меристем. 2. Образование адвентивных побегов тканями экспланта. 3. Возникновение адвентивных побегов в каллусе. 4. Индукция соматического эмбриогенеза в клетках экспланта. 5. Соматический эмбриогенез в каллусной ткани. 6. Формирование придаточных эмбриоидов в ткани первичных соматических зародышей (деление первичных эмбриоидов). H. В. Катаева и Р. Г. Бутенко (1983) выделяют два принципиально различных типа клонального микроразмножения: 1. Активация уже существующих в растении меристем (апекс стебля, пазушные и спящие почки стебля). 2. Индукция возникновения почек или эмбриоидов de novo: а) образование адвентивных побегов непосредственно тканями экспланта; б) индукция соматического эмбриогенеза; в) дифференциация адвентивных почек в первичной и пересадочной каллусной ткани. Основной метод, использующийся при клональном микроразмножении растений — активация развития уже существующих в растении меристем. Он основан на снятии апикального доминирования (рис. 18).
Рис. 18. Схема размножения растений методом активации уже существующих меристем (по А. Р. Родину, Е. А. Калашниковой, 1993): 1 — путем удаления верхушечной меристемы; 2 — добавлением цитокининов в среду (б/г — среда без гормонов, Ц — цитокинин, А — ауксин)
Этого можно достичь двумя путями: а) удалением верхушечной меристемы стебля и последующим микрочеренкованием побега in vitro на безгормональной среде; б) добавлением в питательную среду веществ цитокининового типа действия, индуцирующих развитие многочисленных пазушных побегов. Как правило, в качестве цитокининов используют 6-бензиламинопурин (БАЛ) или 6-фурфуриламинопурин (кинетин) и зеатин. Полученные таким образом побеги отделяют от первичного экспланта и вновь самостоятельно культивируют на свежеприготовленной питательной среде, стимулирующей пролиферацию пазушных меристем и возникновение побегов более высоких порядков. Часто в качестве экспланта используют верхушечные или пазушные почки, которые изолируют из побега и помещают на питательную среду с цитокининами. Образующиеся пучки побегов делят, при необходимости черенкуют и переносят на свежую питательную среду. После нескольких пассажей, добавляя в питательную среду ауксины, побеги укореняют in vitro (рис. 19), а затем переносят в почву, где создают условия, способствующие адаптации растений (рис. 20).
Рис. 19. Образование корней побегами розы при добавлении в питательную среду 2 мг/л 2,4-Д
Рис. 20. Адаптация пробирочных роз к почвенным условиям
В настоящее время этот метод широко используется в производстве посадочного материала сельскохозяйственных культур, как технических, так и овощных, а также для размножения культур промышленного цветоводства (например, гвоздики, рис. 21), тропических и субтропических растений, плодовых и ягодных культур, древесных растений. Для некоторых культур, таких как картофель, технология клонального размножения поставлена на промышленную основу. Применение метода активации развития существующих меристем позволяет получать из одной меристемы картофеля более 100000 растений в год, причем технология предусматривает получение в пробирках микроклубней — ценного безвирусного семенного материала.
Рис. 21. Пробирочная гвоздика
Второй метод — индукция возникновения адвентивных почек непосредственно тканями экспланта. Он основан на способности изолированных частей растения при благоприятных условиях питательной среды восстанавливать недостающие органы и таким образом регенерировать целые растения. Можно добиться образования адвентивных почек почти из любых органов и тканей растения (изолированного зародыша, листа, стебля, семядолей, чешуек и донца луковиц, сегментов корней и зачатков соцветий). Этот процесс происходит на питательных средах, содержащих цитокинины в соотношении с ауксинами 10:1 или 100:1. В качестве ауксина используют ИУК или НУК. Таким способом были размножены многие представители семейства лилейных, томаты, древесные растения (из зрелых и незрелых зародышей). Достаточно хорошо разработана технология клонального размножения земляники, основанная на культивировании апикальных меристем. Меристематические верхушки изолируют из молодых, свободных от вирусных болезней растений, и выращивают на питательной среде МС, содержащей ВАЛ в концентрации 0,1–0,5 мг/л. Через 3–4 недели культивирования меристема развивается в проросток, в основании которого формируются адвентивные почки, быстро растущие и дающие начало новым почкам. В течение 6–8 недель образуется конгломерат почек, связанных между собой соединительной тканью и находящихся на разной стадии развития. Появляются листья на коротких черешках, в нижней части которых формируются новые адвентивные почки. Эти почки разделяют и пересаживают на свежую питательную среду. На среде без регуляторов роста за 4–5 недель формируются нормальные растения с корнями и листьями. От одного материнского растения таким образом можно получить несколько миллионов растений-регенерантов в год. Третий метод, практикуемый при клональном микроразмножении, основывается на дифференциации из соматических клеток зародышеподобных структур, которые по своему виду напоминают зиготические зародыши (рис. 22).
Рис. 22. Соматический эмбриогенез в каллусной ткани
Этот метод получил название соматического эмбриогенеза. В отличие от развития in vivo, соматические зародыши развиваются асексуально вне зародышевого мешка и по своему внешнему виду напоминают биполярные структуры, у которых одновременно наблюдается развитие апикальных меристем стебля и корня. Согласно Стеварду, соматические зародыши проходят 3 стадии развития: глобулярную, сердцевидную, торпедовидную и в конечном итоге имеют тенденцию развития в проросток. На рисунке 3 показан конечный результат развития — растение пшеницы. Наиболее впечатляющим применением метода соматического эмбриогенеза стало размножение гвинейской масличной пальмы (Elaeis guineensis), масло которой широко используется при производстве маргарина и пищевого масла. Масличная пальма в природе не образует побегов и боковых ростков, что затрудняет ее вегетативное размножение. Культивирование черенков in vitro также невозможно. Было решено получить скопления клеток недифференцированной ткани (каллусы) путем дедифференцировки специфических тканей, а затем культивировать их до регенерации целых проростков. В первой культуральной среде каллусы из фрагментов листьев развивались в течение 90 дней, при переносе во вторую и третью культуральные среды превращались в "эмбриоиды". Эмбриоиды размножались самопроизвольно, в течение месяца число эмбриоидов возрастало втрое, а за год из 10 эмбрионов можно было получить потомство численностью 500000 растений. Формирование эмбриоидов в культуре тканей осуществляется в несколько этапов. Сначала происходит дифференциация клеток под влиянием ауксинов, добавленных в питательную среду (2,4-Д) и превращение их в эмбриональные. Получить эмбриоиды из этих клеток можно уменьшая концентрацию ауксинов или исключая их из питательной среды. Соматические зародыши представляют собой полностью сформированные зародыши, из которых путем соответствующего капсулирования можно получить искусственные семена. Четвертый метод клонального микроразмножения — дифференциация адвентивных почек в первичной и пересадочной каллусной ткани (рис. 23).
Рис. 23. Дифференциация придаточных почек в каллусной ткани
Практически он мало используется с целью получения посадочного материала in vitro. Это связано с тем, что при частом пассировании каллусной ткани может изменяться плоидность регенерируемых растений, наблюдаются структурные перестройки хромосом и накопление генных мутаций. Наряду с генетическими изменениями отмечаются и морфологические: низкорослость, неправильное жилкование листьев, образование укороченных междоузлий, пониженная устойчивость к болезням и вредителям. В то же время, некоторые недостатки этого метода в селекционной работе оборачиваются преимуществами. Кроме того, в некоторых случаях он является единственно возможным способом размножения растений в культуре тканей. Через каллусную культуру успешно размножаются сахарная свекла, злаковые (рис. 24), представители рода Brassica, подсолнечник и другие культуры.
Рис. 24. Формирование побегов каллусной тканью пшеницы
Оздоровление посадочного материала от вирусов Основное преимущество клонального микроразмножения — получение генетически однородного, безвирусного посадочного материала. Предположение о возможности отсутствия вирусов в меристематических тканях больных растений впервые было высказано в 1936 г. Чунгом, а позднее, в 1943 г., и Уайтом. В 1949 г. этот факт был подтвержден экспериментально. В 1952 г. Морелю и Мартену из Национального агрономического института (Франция) удалось получить безвирусные георгины из зараженных растений. Структурной основой используемого на практике явления служит специфика строения точки роста растений: дистальная ее часть, представленная апикальной меристемой, у разных растений имеет средний диаметр 200 мкм и высоту от 20 до 150 мкм. В нижних слоях дифференцирующиеся клетки меристемы образуют прокамбий, дающий начало пучкам проводящей системы. Известно, что успех клонального микроразмножения зависит от меристематического экспланта. При этом отмечается закономерность: чем больше листовых зачатков и тканей, тем легче идут процессы морфогенеза, заканчивающиеся образованием целого растения. Вместе с тем, при таком развитии конуса нарастания увеличивается риск быстрой транспортировки вируса по проводящей системе. Кроме того, даже небольшой меристематический эксплант может содержать вирусы, проникшие в клетки в результате медленного распространения через плазмодесмы. В целом, эффективность применения апикальной меристемы в качестве метода оздоровления зараженных вирусами растений может оказаться довольно низкой. Снизить риск попадания вирусов в здоровые ткани можно путем применения предварительной термо- или химиотерапии исходных растений. Метод термотерапии применяется как в условиях in vivo, так и in vitro и предусматривает использование горячего сухого воздуха. Для объяснения механизма освобождения растений от вирусов в процессе термотерапии существуют различные гипотезы. Согласно одной их них при высоких температурах разрушаются белковая оболочка и нуклеиновая кислота вируса. Вторая гипотеза предполагает действие высоких температур на вирусы через метаболизм растений. При такой температуре начинает преобладать деградация вирусных частиц, а синтез их, наоборот, уменьшается. Растения, подвергающиеся термотерапии, помещают в термокамеры, где температура в течение первой недели повышается с 25 до 37 °C путем ежедневного увеличения температуры на 2 градуса. Все остальные режимы обязательно поддерживаются в оптимальном состоянии: освещенность, высокая относительная влажность воздуха, определенный фотопериод. Продолжительность термостатирования зависит от состава вирусов и их термостойкости. Если для гвоздики достаточно 10–12 недельного воздействия теплом, то для хризантемы этот период превышает 12 недель. Помимо положительного действия высоких температур на освобождение от вирусов, выявлено аналогичное влияние их на точку роста и процессы морфогенеза некоторых цветочных культур (гвоздики, фрезии) в условиях in vitro. Высокие температуры увеличивают коэффициент размножения на 50–60 %, повышаю адаптацию пробирочных растений к почвенным условиям и позволяют получить больше безвирусных маточных растений. Другой способ оздоровления — химиотерапия. В питательную среду, на которой культивируют апикальные меристемы, добавляют препарат вирозола в концентрации 20–50 мг/л. Это противовирусный препарат широкого спектра действия. Применение его позволяет увеличить число безвирусных растений с 40 % до 80-100 %.
СОЗДАНИЕ ИСКУССТВЕННЫХ АССОЦИАЦИЙ КУЛЬТИВИРУЕМЫХ КЛЕТОК ВЫСШИХ РАСТЕНИЙ С МИКРООРГАНИЗМАМИ
Цели создания ассоциаций Создание искусственных ассоциаций — новое, сравнительно молодое направление клеточной инженерии по получению новых клеток и клеточных систем путем введения микроорганизмов в клетки или в популяции культивируемых клеток растений. Экспериментальные клеточные системы называются ассоциациями. Ассоциации могут быть как внутриклеточные (эндосимбиотического типа), так и межклеточные (экзосимбиотического типа). В первом случае микроорганизмы вводят в изолированные протопласты высших растений. Во втором — совместно культивируют клетки и ткани растений с микроорганизмами. При создании ассоциаций предполагается, что клетки и их популяции должны приобретать новые свойства, обусловленные присутствием в них микроорганизмов. Цели создания популяций: 1. Экспериментальная проверка гипотезы теории симбиотического происхождения эукариотической клетки, которое предположительно проходило через стадии эндо- и экзосимбиоза. Реконструкция отдельных стадий эволюционного процесса симбиогенеза. 2. Моделирование природных симбиотических отношений растений и микроорганизмов, играющих огромную роль в процессе фиксации атмосферного азота (обеспечение связанным азотом природных экосистем, а также агроценозов). 3. Повышение продуктивности растительных клеток-продуцентов экономически важных веществ. 4. Получение растений с новыми свойствами, при условии, что отношения, складывающиеся между клетками партнеров при совместном выращивании, сохраняются в растениях-регенерантах. В литературе обсуждаются возможность улучшения таким способом сельскохозяйственных растений, а также получение растений со способностью к автономной фиксацией азота. Первые две цели имеют значение для решения теоретических вопросов биологии, последние две носят ярко выраженный прикладной характер.
Повышение продуктивности сельскохозяйственных растений Одна из целей культивирования растительных клеток — получение важных для медицины и ряда отраслей промышленности веществ. Для того чтобы производство было рентабельным, необходимо культивировать их на простых по составу пита тельных средах. В то же время, среды достаточно сложны по составу и включают в себя витамины, гормоны, источники углеродного питания, так как клетки в культуре являются гетеротрофами или обладают ограниченной способностью к фотосинтезу. Совмещение в культивируемых клетках способности к фотосинтезу и биосинтезу специфических продуктов — маловероятно. Поэтому введение в такие культуры микроорганизмов, синтезирующих субстраты для роста растительных клеток или предшественники для биосинтеза полезных веществ представляется весьма заманчивым. В микробиологии опыт смешанного культивирования есть. Он показывает, что системы микроорганизмов более эффективны, чем монокультуры. Их используют для очистки сточных вод, получения ферментов, биологически активных веществ (ауксины, витамины, антибиотики). Считается, что в биотехнологии найдут применение смешанные популяции, включающие в себя как сочетания нескольких штаммов микроорганизмов, так и представителей царств растений и животных. Улучшение сельскохозяйственных растений предполагает получение растений, способных к фиксации молекулярного азота. При внесении удобрений используется от 30 до 50 % внесенного азота. Другой путь поступления азота — биологическая фиксация молекулярного азота. Большая часть осуществляется азотфиксирующими симбионтами, но этот процесс характерен лишь для некоторых видов высших растений и микроорганизмов. Для повышения доли биологической фиксации азота используют 3 подхода: 1. Инокуляция азотфиксирующими микроорганизмами (бактериальные удобрения). Недостаток — низкая выживаемость интродуцируемых чистых культур и вытеснение их естественной почвенной микрофлорой. 2. Создание азотфиксирующих растений методами генной инженерии. При этом предлагается вводить гены nif в протопласты высших растений. Препятствия на этом пути: процесс требует большого количества энергии, которой нет в растительной клетке, нет также систем транспорта, запасов железа и молибдена, необходимых для синтеза нитрогеназы, нет систем защиты нитрогеназы от инактивации кислородом. 3. Введение целых азотфиксирующих организмов в растения. Такие ассоциации должны учитывать особенности организации природных азотфиксирующих симбиозов: — целостность обоих партнеров, — интеграция партнеров в пределах организма макросимбионта, — относительная обособленность макросимбионта. С помощью клеточной инженерии можно осуществить жизнедеятельность азотфиксирующих организмов в клетках и тканях культурных растений. При этом возможна проверка большого числа сочетаний партнеров. В процессе культивирования возможна также адаптация партнеров друг к другу. Кроме того, бактериальные симбионты могут быть интегрированы в ткани растений с сохранением их интактности, что позволит оградить нитрогеназу от кислорода, выделяемого растением в процессе фотосинтеза. В настоящее время эти положения могут быть подтверждены рядом экспериментов.
Создание эндосимбиотических ассоциаций В изолированные протопласты растений вводили микроорганизмы следующих систематических групп: бактерии, дрожжи, цианобактерии, цианеллы. Существует несколько способов введения микроорганизмов в протопласты (рис. 25):
Рис. 25. Введение микроорганизмов в изолированные протопласты высших растений (по L. Folke, O.Gamborg, 1981)
1. Эндоцитоз (инвагинация плазмалеммы), при этом везикула с микроорганизмом высвобождается в цитоплазме протопласта. 2. Интеграция (слияние) мембран протопласта и микроорганизма, органеллы водорослей высвобождаются в цитоплазму протопласта, но при этом они не окружены плазмалеммой протопласта. 3. Заключение микроорганизма в искусственные мембраны — липосомы. Например, в протопласты лука вводили цианобактерии, заключенные в липидные капли. Каждый из методов имеет свои достоинства и недостатки. В первом случае везикулы окружены цитоплазматической мембраной протопласта, в результате чего может произойти слияние с лизосомами и разрушение микроорганизма. Во втором случае происходит нарушение целостности микроорганизма: в цитоплазме оказываются органеллы. а) эндоцитоз (поглощение путем инвагинации мембраны) б) слияние мембран в) слияние мембраны протопласта с липосомой В третьем случае в цитоплазме оказываются интактные микроорганизмы, что является одним из условий воспроизведения естественного симбиоза. Такого же результата можно достичь используя метод микроинъекций (например — цианобактерий) в пространство между стенкой и плазмолированной клеткой лука. Этот способ можно рассматривать либо как стадию перед попаданием бактерий в цитоплазму, либо как получение ассоциаций межклеточного типа. К сожалению, жизнеспособных систем на основе изолированных протопластов в настоящее время не получено, так как в процессе культивирования происходило разрушение либо микроорганизма, либо протопласта. Необходимы дальнейшие исследования.
Экзосимбиотические ассоциации Партнеры те же, в основном — азотфиксаторы (Rhizobium, Azotobacter, Azospirillum), Chlorella и различные виды грибов. Общий принцип создания таких ассоциаций — совместное культивирование клеток растений с микроорганизмами. Способы ведения микроорганизмов в культуру могут быть различны: внесение микроорганизмов непосредственно в культуру; высев на поверхности агара рядом с каллусом, но без соприкосновения; разделение бактериальных клеток и культуры мембранами, обеспечивающими обмен продуктами метаболизма, но не допускающими контакта между партнерами. Рассмотрим результаты попыток создания ассоциаций с различными партнерами.
Ассоциации с клубеньковыми бактериями Сначала создавались упрощенные модельные системы для изучения симбиоза бобовых растений с клетками растений. Каллусную ткань сои инфицировали клетками Rhizobium. Были получены подобия инфекционных нитей и бактероидов в каллусной клетке. В таких ассоциациях клетки бактерий проявляли нитрогеназную активность (НГА, отсутствующую в чистой культуре. Это свидетельствовало о процессе азотфиксации. Далее были созданы и азотфиксирующие ассоциации Rhizobium с культурами тканей небобовых растений. В таких ассоциациях клетки бактерий были локализованы на поверхности растительных клеток, проникали вглубь каллуса по межклетникам и изредка обнаруживались внутри клеток. В основном сохраняли палочковидную форму без образования бактероидов. В большинстве таких экспериментов бактериальные клетки угнетали рост растительных и усиливали их отмирание. Сообщения о положительном взаимном влиянии единичны. Есть одна удачная попытка регенерации растений табака из ассоциации каллусной ткани с Rhizobium.
Ассоциации со свободноживущими азотфиксаторами Azotobacter, Azospirillum живут в ризосфере растений и иногда образуют с ними ассоциации. Главная задача проведения экспериментов с этими бактериями — получение эффективных азотфиксирующих систем, растущих на среде без связанного азота, с перспективой регенерации растений. В таких экспериментах была обнаружена видовая специфичность. Каллусные культуры табака, проса быстро "обрастали" Azospirillum, но через 4 недели погибали. Ткань сахарного тростника субкультивировали в аналогичных условиях 18 месяцев, при этом стабильные ассоциации формировались только на среде с низким содержанием связанного азота или без него. Бактерии проявляли НГА во всех случаях. И Azotobacter, и Azospirillum в соответствующих ассоциациях были локализованы на поверхности или в межклетниках и никогда не проникали внутрь клетки. Растения-регенеранты пока не получены.
Ассоциации с зелеными водорослями Каллус моркови инокулировали одним из штаммов Chlorella, культивировали на свету на среде с дефицитом азота, каллус жил несколько месяцев. Контрольные же растения погибали. Клетки водоросли не проникали внутрь растительных.
Ассоциации с грибами Ткань руты раздельно культивировали с различными грибами, при этом на клетки растений влияли диффундирующие через агар выделения гриба. В некоторых случаях добавляли культуральную жидкость грибов. Совместное культивирование с Botritis allii увеличивало синтез алкалоидов в 10 раз по сравнению с контролем, а добавление культуральной жидкости — в 50 раз.
Цианобактерии в искусственных ассоциациях с растительными клетками Цианобактерии как партнеры в искусственных ассоциациях имеют ряд особенностей: — предполагают, что древние цианобактерии сыграли роль в формировании эукариотической клетки; — чаще других фототрофов вступают в симбиотические отношения с другими организмами в природе; — в симбиозах осуществляют различные метаболические функции: с автотрофами играют роль азотфиксаторов, с гетеротрофами обеспечивают партнеров продуктами фотосинтеза; — способны выделять в среду углеводы, аминокислоты, пептиды, витамины, гормоны; — в процессе фотосинтеза выделяют кислород, которые растения в процессе дыхания потребляют; — находят практическое применение в улучшении обеспечения растений связанным азотом. Препятствием могут служить различные требования к условиям культивирования: у клеток растений оптимум pH 5–5.5, а цианобактерий нейтрально-щелочной (7-10); для растительных клеток температурный оптимум 24–27 °C, для бактерий — 30–40 °C; концентрация минеральных солей в среде для выращивания растительных клеток выше, чем требуется для цианобактерий. В исследованиях, проведенных на кафедре клеточной физиологии и иммунологии МГУ, проверялись различные сочетания партнеров, с использованием как каллусных, так и суспензионных культур. Был отмечен ряд физиологических реакций: от ингибирования одного партнера другим до взаимной стимуляции процессов жизнедеятельности. В суспензионных культурах клеток табака и Anabaena nidulans отмечено следующее: — взаимная стимуляция роста, когда в условиях, неоптимальных для роста каждого компонента системы, происходила взаимная стимуляция; — двухфазность ростовых процессов: за пиком роста цианобактерий следует пик роста растительных клеток, сопровождающийся частичной деградацией цианобактерий. Предполагается, что в первой фазе роста отношения партнеров носят характер мутуализма; — цианобактерии адсорбируются на поверхности клеток, проникают вглубь клеточных агрегатов, но часть партнеров остается в суспензии в свободном состоянии; — растительные клетки потребляют продукты фотосинтеза цианобактерий; — между клетками растений и микроорганизмов устанавливаются метаболические взаимодействия, которые могут привести к стимуляции видоспецифических биосинтезов растительных клеток. Ассоциации каллусных тканей с цианобактериями получали двумя способами: — использовали изолированные протопласты, выделенные из дефицитных по хлорофиллу участков мезофилла листа, — использовали каллус, выделенный из бесхлорофильного участка листа, к которому подсевали суспензию A.variablis. При этом каллусные культуры стабильно росли не менее 2 лет. Антагонизма не было. Добавляя в среду ауксины и цитокинины, удалось вызвать органогенез у каллуса табака, ассоциированного с цианобактерией. Сформировались белые побеги регенерантов табака с участками сине-зеленого цвета, содержащими бактерии. В регенерантах из каллуса цианобактерии имели внутритканевую локализацию и обнаруживались только в листьях. Их также находили на поверхности листа, стебля (рис. 26), в сосудистой системе, устьицах.
Рис. 26. Цепочки Anabaena variabilis в углублениях складчатой поверхности стебля табака (по Р. Г. Бутенко и др., 1987)
Результаты по получению ассоциаций цианобактерий с растениями — регенерантами представляют интерес в связи с проблемой повышения доли биологически связанного азота в азотном питании растений.
МЕТОДЫ СОХРАНЕНИЯ ГЕНОФОНДА
При получении клеточных линий с полезными признаками встает проблема сохранения этих признаков. Растения могут хранить генетическую информацию в семенах, однако этот источник не вполне надежен, так как со временем из-за мутаций всхожесть семян падает. Кроме того, некоторые растения размножаются только вегетативно. Этим обусловлена необходимость сохранения части материала in vitro. С другой стороны, в некоторых случаях удается получить новые клеточные линии, синтезирующие большее количество вторичных метаболитов, то есть более продуктивные, которые тоже нуждаются в сохранении. Для исследования физиологических и биохимических процессов, протекающих в тканях, также требуются стандартные исходные культуры, чем вызвана необходимость сохранять материал в течение определенного промежутка времени, когда идут серийные эксперименты. Все это делает проблему сохранения генофонда весьма актуальной. Можно, конечно, пассировать и перевивать клеточные культуры. Однако при этом возникает опасность сомаклональной изменчивости, накопления мутаций, контаминаций (заражения чужеродным генетическим материалом). Это также требует определенных финансовых и трудовых затрат (необходимость частых пересадок, расходы, связанные со средой и т. д.). Цель исследователей состоит в увеличении интервала между пересадками. Существует разные подходы к сохранению куль- — криосохранение, — замедление роста, — сушка (распылительная и лиофильная) — для клеток микроорганизмов.
Криосохранение Криосохранение — замораживание при сверхнизких температурах. Обычно его проводят в жидком азоте, при температуре -196 °C. Успех низкотемпературной консервации зависит от ряда факторов: — вид и тип клеток, — их концентрация в суспензии, — состав среды для консервирования, — вид и концентрация криопротектора, — режим охлаждения и отогрева, — способ реабилитации клеток после отогрева. Существенную роль в успешном замораживании клеток играет их морфофизиологическое состояние: клетки, находящиеся в стационарной фазе роста, менее устойчивы к повреждающему действию низкотемпературной консервации, чем клетки, находящиеся в экспоненциальной фазе роста. Клетки для замораживания отбирают в середине экспоненциальной фазы ростовой кривой. Немаловажное значение имеет и плотность замораживаемой суспензии. Оптимальные результаты по восстановлению клеток были получены при замораживании клеточнойсуспензии плотностью 1∙105-5∙106 клеток в 1 мл. Для растительных клеток часто требуется предварительное культивирование в особых условиях. В среду добавляют различные вещества, например: — 2–6 % маннит или сорбит для уменьшения размера вакуолей; — аминокислоты, в первую очередь пролин, который служит для связывания воды в клетке (концентрация до 1 моля или 11,5 %), аспарагин, γ-аминомасляную кислоту слоту; — диметилсульфоксид (ДМСО), который добавляют к среде для предкультивирования в концентрации от 2,5 до 10 % на 48 часов для увеличения проницаемости цитоплазматической мембраны; — кроме того, применяют искусственное закаливание к холоду, когда снижают температуру культивирования, имитируя естественный осенний процесс подготовки к периоду зимнего покоя (применим только для растений умеренного климата). Клеточные культуры выдерживают несколько суток при температуре +8 — +10 °C, а затем при +2 — +5 °C в течение 1–6 недель. Процесс замораживания растительных клеток от животных отличает, в основном, наличие этапа предварительного культивирования. Криопротекторы — вещества, позволяющие снизить повреждающее действие физико-химических факторов при криоконсервировании. К ним относятся сахароза, декстран, этиленгликоль, поливинилпирролидон, диметилсульфоксид (ДМСО), глицерин. Для определения токсичности криопротектора клетки выдерживают при комнатной температуре в различных его концентрациях в течение 30–50 минут, после чего определяют их жизнеспособность. Дополнительно оценивают его протективные свойства путем пробного замораживания и оттаивания культур. Наиболее часто в качестве криопротекторов используют глицерин и ДМСО. Перед добавлением криопротектора суспензию клеток концентрируют путем центрифугирования, надосадочную жидкость сливают. Криопротекторы вносят в культуру за час до замораживания, что приводит к изменению проницаемости мембраны, изменению точки замерзания и оттаивания. Программы охлаждения могут быть различными, но для всех них характерна медленная скорость охлаждения. При замораживании происходит образование льда внутри и снаружи клеток. Характер этих изменений зависит от изучаемого образца и обработки криопротекторами, но главным образом, от скорости охлаждения. При медленном охлаждении происходит образование внеклеточного льда, приводящее к обезвоживанию клетки до того, как будет достигнута точка замерзания цитоплазмы. При быстром охлаждении клетки быстрее замораживаются изнутри, медленнее обезвоживаются, что приводит к образованию кристаллов льда внутри клетки. В этом случае клетки повреждаются. Обычно охлаждение проводят в два этапа (рис. 27):
Рис. 27. Замораживание клеток: а) быстрое, б) медленное, поэтапное
1. От +20 до -28 °C со скоростью 1 градус в минуту (для растительных клеток скорость замораживания 0,5 градуса в минуту до -35 °C), выдерживают при этой температуре 15 минут. 2. Погружение в жидкий азот (мгновенное охлаждение до — 196 °C) Замораживание производят в специальных аппаратах. При их отсутствии — на спиртовой бане (0,5–1 литр спирта наливают в термос с металлической колбой, погружают в него ампулы на 15 минут и добавляют при помешивании жидкий азот или сухой лед; доводят температуру до -32 °C (температура должна быть не выше -28 и не ниже -32 °C). Далее переносят ампулы в жидкий азот. При размораживании ампулы пинцетом переносят в водяную баню с температурой +37 — +40 °C, ампула объемом в 1 мл размораживается в течение 0,5–1 минуты. После размораживания клетки отмывают либо в ростовой среде (животные), либо в поддерживающей среде. Растительные клетки также можно отмывать 3-10 % раствором сахарозы. Далее клетки проверяют на жизнеспособность с помощью витальных красителей, окрашивающих мертвые клетки. Окончательным критерием служит четкое возобновление роста на стандартных питательных средах, используемых для данной культуры. Перевиваемые культуры животных клеток после размораживания имеют повышенную чувствительность к вирусам, которая проявляется в течение первых двух пассажей. Далее чувствительность возвращается к исходной.
Замедление роста Замедления роста можно добиться следующими методами: 1. Хранение под слоем минерального масла (для бактериальных и грибных культур). 2. Изменение газового состава и атмосферного давления внутри культурального сосуда. 3. Изменение светового режима. 4. Охлаждение до температуры прекращения активного роста. 5. Применение гормональных и осмотических ингибиторов. Из гормональных ингибиторов наиболее часто используют хлорхолинхлорид (для растительных клеток), из осмотических — маннит в концентрации 3–6 %. 6. Замена СаСL2 на Са(NО3)2 в питательных средах. Для картофеля в качестве способа, позволяющего сохранить генофонд, рекомендуется клубнеобразование в пробирках.
БЕСКЛЕТОЧНЫЕ СИСТЕМЫ
Мембраны хлоропластов Американский ученый М. Кальвин, чьи исследования в области изучения механизма фотосинтеза были отмечены Нобелевской премией, в 1972 году выдвинул идею создания фотоэлемента, в котором в качестве источника электрического тока служили мембраны хлоропластов. Основной компонент таких мембран — хлорофилл, способный при освещении отдавать и принимать электроны. В качестве проводника, контактирующего с хлорофиллом, Кальвин использовал оксид цинка. Мембраны, содержащие хлорофилл, помещали в раствор ферментов, действующих как катализаторы ЭТЦ. На свету происходит фотолиз воды: Н2O —> Н2 + 1/2 O2. При освещении этой системы в ней также возникал электрический ток плотностью 0,1 мкА на см2. Такой фотоэлемент функционировал недолго, поскольку хлорофилл вскоре терял способность отдавать электроны. Для того чтобы продлить время действия фотоэлемента, был использован дополнительный источник электронов — гидрохинон. В такой системе хлорофилл отдавал не только свои электроны, но и электроны гидрохинона. Полученный таким образом фотоэлемент площадью 10 м2 может обладать мощностью 1 кВт. Японский ученый Фудзио Такахаси для получения электроэнергии использовал хлоропласты из листьев салата. Транзисторный приемник, к которому была присоединена такая солнечная батарейка, успешно работал. Если из системы убрать проводник и индуцировать образование водорода и кислорода, то система может служить также прототипом фотореактора, при помощи которого энергия Солнца запасается в ценном топливе — водороде. Преимущества системы: — наличие избытка субстрата — воды, — нелимитируемый источник энергии — Солнце, — продукт (водород) можно хранить, не загрязняя атмосферу, — продукт имеет высокую теплотворную способность (29 ккал/г) по сравнению с углеводородами (3,5 ккал/г), — процесс протекает при нормальной температуре без образования промежуточных токсических веществ, — процесс циклический, так как при окислении продукта образуется субстрат — вода.
Мембраны хлоропластов можно иммобилизовать, закрепляя их в геле. Получение фотогальванических элементов с использованием бактериальных мембран Другой механизм превращения энергии существует у галофильных бактерий. Halobacterium halobium используют энергию света, поглощаемую пурпурным пигментом бактериородопсином, находящимся в мембране клеток. Этот белок с необычными свойствами был выделен и описан в 1973 году У. Стохениусом и Д. Остерхельтом. С его помощью бактерии улавливают энергию Солнца. Поглощение света вызывает химические и физические превращения в молекуле пигмента, приводящие к переносу протонов с одной стороны мембраны на другую, при этом создаётся электрохимический градиент. Разность потенциалов может быть использована для генерирования электрического тока. Бактериородопсин несложно выделить из бактерий. Для этого бактерии помещают в воду, где они переполняются водой[72] и лопаются. Мембраны, содержащие бактериородопсин, не разрушаются в воде из-за прочной упаковки молекул пигмента, которые образуют белковые кристаллы — так называемые фиолетовые бляшки. В них молекулы бактериородопсина объединены в триады, а триады — в правильные шестиугольники. Бляшки крупные, легко отделяются центрифугированием. После промывания осадка получается паста фиолетового цвета. На 75 % она состоит из бактериородопсина и на 25 % из фосфолипидов, заполняющих промежутки между белковыми молекулами. Бактериородопсин устойчив к факторам внешней среды: не утрачивает активность при нагревании до 100 °C, хранится в холодильнике годами, устойчив к кислотам и химическим окисляющим агентам. Устойчивы и фосфолипиды фиолетовых бляшек. H.halobium можно культивировать в водоемах с высокой концентрацией хлористого натрия и других минеральных солей. Из 10 литров бактериальной культуры получают 0,5 грамма мембран, содержащих 100000 молекул пигмента. Бактериородопсин осаждают с помощью катионов кальция или другим способом. Пигмент можно фиксировать на подложках, обладающих физическими и химическими свойствами для транспорта протонов, и создавать на их основе системы, генерирующие электрический ток. При освещении таких систем на мембране обнаруживается электрический потенциал, то есть бактериородопсин функционирует как генератор электрического тока. В лаборатории В. П. Скулачева были созданы фотогальванические элементы для генерирования тока силой 800 мкА. В них применялись мембранные фильтры, пропитанные фосфолипидами с бактериородопсином и хлорофиллом. Такие фильтры, соединенные последовательно, могут служит в качестве электрической батареи.
Бесклеточные белоксинтезирующие системы (ББСС) ББСС используются для изучения матричной активности иРНК и анализа транслируемых с них полипептидов. В их состав входят: рибосомы, матрица (искусственная или природная РНК), белковые факторы трансляции, аминоацил т-РНК, АТФ, одновалентные и двухвалентные катионы (К, Са), буферный раствор для поддрежания гомеостаза, аминокислоты. В генной инженерии бесклеточные белоксинтезирующие системы используются для исследования кодирующего потенциала и механизмов экспрессии клонированных генов in vitro, и на промежуточных этапах конструирования рекомбинантных генов для идентификации мРНК или фрагментов ДНК по кодируемым белкам. По происхождению компонентов ББСС можно классифицировать как прокариотические и эукариотические. Наиболее распространенные прокариотические белоксинтезирующие системы — на основе экстрактов из кишечной палочки (E.coli). Из эукариотических белоксинтезирующих систем для трансляции матриц эукариот применяют 2 основные системы: из ретикулоцитов кролика и из зародышей пшеницы. Эти системы являются универсальными, в них можно транслировать любые матрицы. Для проведения анализа готовят реакционную смесь, состоящую из ретикулоцитного лизата или экстракта зародышей и смеси аминокислот, меченых аминокислот, АТФ, буфера, матричной РНК и других компонентов. Смесь инкубируют, после чего проверяют включение метки во вновь синтезированные белки (по радиоактивности) и тестируют белки с помощью электрофореза. Трансляция in vitro полезна при уточнении роли отдельных компонентов системы синтеза белка, так как их можно удалять и добавлять по мере необходимости. Ее использование помогает при расшифровке генетического кода. В некоторых бесклеточных системах транслируют предварительно очищенную мРНК или используют эндогенную мРНК, присутствующую в полисомах. В других белоксинтезирующих системах — системах сопряженной транскрипции и трансляции, — синтез мРНК и ее трансляция рибосомами идут одновременно. В качестве матричной РНК также используются искусственные полинуклеотиды известного состава. В настоящее время механизмы трансляции in vitro применяются и для определения механизмов распределения белка по различным внутренним компартаментам. Подробнее можно почитать здесь: бесклеточные системы синтеза белка.
ЛИТЕРАТУРА К РАЗДЕЛУ «КУЛЬТУРЫ РАСТИТЕЛЬНЫХ КЛЕТОК»
1. Артамонов В.И. Биотехнология — агропромышленному комплексу. М.: Наука, 1989. 160 с. 2. Артамонов В. И. Сельские профессии биотехнологии. М.: Изд-во МСХА, 1992. 127 с. 3. Атанасов А. Биотехнология в растениеводстве. Новосибирск: ИЦ и Г СО РАН, 1993. 241 с. 4. Баулина О.И., Лобакова Е.С., Корженевская Т.Г., Бутенко Р.Г., Гусев М.В. Ультраструктура клеток женьшеня и цианобактерии Chlorogloepsis fritschii в ассоциации при культивировании в темноте. // Вестник Московского университета. Сер. Биология. 1995. № 2. С. 3–16. 5. Бекер М.Е., Лиепиньш Г.К., Райпулис Е.П. Биотехнология. М.: Агропромиздат, 1990. 334 с. 6. Беккужина С.С., Никифорова И.Д. Клональное размножение и тестирование на солеустойчивость растений — регенерантов яровой пшеницы. // Новые методы биотехнологии растений. Тез. II российского симпозиума. Пущино, 1993. С. 113. 7. Биотехнология. / Под ред. А.А. Баева. М.: Наука, 1984. 309 с. 8. Биотехнология сельскохозяйственных растений. М.: Агропромиздат, 1987. С.241–244. 9. Блинов А.Г. Бесклеточные белоксинтезирующие системы. // Методы молекулярной генетики и генной инженерии. Новосибирск: Наука, 1990. С. 80–86. 10. Борисюк Н.В. Молекулярно — генетическая конституция соматических гибридов. // Биотехнология. Итоги науки и техники ВИНИТИ АН СССР. М., 1988. Т. 9. С. 73 -113. 11. Борнман X. Реконструкция клеток растений: Brassica в качестве объекта изучения. // Биология культивируемых клеток и биотехнология растений. М.: Наука, 1991. С. 85–95. 12. Бутенко Р.Г. Изолированные протопласты растений — объект и модель для физиологических исследований. // Культура клеток растений. М.: Наука, 1981. С. 69–84. 13. Бутенко Р.Г. Клеточные технологии для получения экономически важных веществ растительного происхождения. // Культура клеток растений и биотехнология. М.: Наука, 1986. С. 3–20. 14. Бутенко Р.Г. Культура изолированных тканей и физиология морфогенеза растений. М.: Наука,1964. 272 с. 15. Бутенко Р.Г. и др. Клеточная инженерия. — М.: Высшая школа, 1987. 16. Валиханова Г. Ж. Биотехнология растений. Алматы: Конжык, 1996. 272 с. 17. Валиханова Г.Ж., Рахимбаев И.Р. Культура клеток и биотехнология растений. Учебное пособие. Алма-Ата: изд. КазГУ, 1989. 80 с. 18. Высоцкая О.Н. Длительное сохранение in vitro коллекции растений земляники. // Физиология растений. 1994. Т. 41, № 6. С. 935–941. 19. Высоцкий В.А. Клональное микроразмножение растений. // Культура клеток растений и биотехнология. М.: Наука, 1986. С. 91 — 102. 20. Глеба Ю.Ю. Гибридизация соматических клеток растений. // Культура клеток растений. М.: Наука, 1981. С. 85–91. 21. Глеба Ю.Ю., Зубко М. К. Теоретические и прикладные аспекты клеточной инженерии растений // Биотехнология. Итоги науки и техники ВИНИТИ АН СССР. М., 1988. Т. 9. С. 3–72. 22. Глеба Ю.Ю., Сытник К.М. Слияние протопластов и генетическое конструирование высших растений. Киев: Наук, думка, 1982. 104 с. 23. Дмитриева Н.Н. Проблема регуляции морфогенеза и дифференциации в культуре клеток и тканей растений. // Культура клеток растений. М.: Наука, 1981. С. 113–123. 24. Зоринянц С.Э. Цитофизиологические характеристики и реорганизация генома длительно культивируемой суспензии клеток Triticum timopheevii. Автореф. дисс… к.б.н. М., 1996. 24 с. 25. Калинин Ф.Л., Сарнацкая В.В., Полищук В.Е. Методы культуры ткани в физиологии и биохимии растений. Киев: Науковадумка, 1980. 488 с. 26. Карабаев М. Культивируемые клетки пшеницы и кукурузы: физиологические и биотехнологические аспекты. Автореф. дисс… д.б.н. М., 1994. 49 с. 27. Карначук P.А., Гвоздева Е.С. Влияние света на баланс фитогормонов и морфогенез в культуре ткани зародышей пшеницы. // Физиология растений. 1998. Т. 45, № 2. С. 289–295. 28. Катаева Н.В., Аветисов В.А. Клональное размножение в культуре ткани. // Культура клеток растений. М.: Наука, 1981. С. 137–149. 29. Катаева Н.В., Бутенко Р.Г. Клональное микроразмножение растений. М.: Наука, 1983. 96 с. 30. Кефели В.И., Дмитриева Г.А. Биотехнология: курс лекций. Пущино, 1989. 96 с. 31. Кирай 3., Барабаш 3. Результаты и перспективы использования биотехнологии в растениеводстве и защите растений. // Международный агропромышленный журнал. 1990. № 3. С. 7–10. 32. Константинова Т.Н., Аксенова Н.П., Сергеева Л.И., Чайлахян М.Х. Взаимное влияние света и гормонов на регуляцию морфогенетических процессов в культуре in vitro. // Физиология растений. 1998. Т. 34, № 4. С. 795–802. 33. Корженевская Т.Г., Бутенко Р.Г., Гусев М.В. Ассоциации цианобактерий с культивируемыми клетками и тканями высших растений. // Культура клеток растений и биотехнология. М.: Наука, 1986. С. 225–242. 34. Кузьмина Н.А., Внукова В.В. Влияние состава питательной среды на хромосомные числа твердой пшеницы (Triticum durum Desf.) in vitro. // Естественные науки и экология. Вып. 2. Омск: изд-во ОмГПУ, 1997. С. 28–33. 35. Кучко А.А. Гибридизация соматических клеток растений методом слияния изолированных протопластов. // Культура клеток растений и биотехнология. М.: Наука, 1986. С. 144–159. 36. Левенко Б.А., Сергеева Л.Е., Виноградов В.А., Ларькина Н.И., Миронов Е.К. Отбор и анализ устойчивых к солевому и водному стрессу клеточных линий табака и регенерантов из них. // Экспериментальная генетика растений в ускорении селекционного процесса. — Киев: Наукова думка, 1989. - с. 101–110. 37. Мамаева С.Е. Хромосомный анализ культивируемых клеток. // Методы культивирования клеток. Л.: Наука, 1988. С. 78 98. 38. Методы клеточной биотехнологии растений. Киев, 1987. 53 с. 39. Методы молекулярной биологии, биохимии и биотехнологии растений. Алма-Ата: Наука, 1988. 168 с. 40. Милов Д., Изворска Н. Состояние и дальнейшее развитие работы с культурами in vitro. // Международный агропромышленный журнал. № 2. 1990. С. 72–78. 41. Новак Ф.Й. Индукция гаплоидов в культуре тканей и их значение в селекции растений. // Культура клеток растений и биотехнология. М.: Наука, 1986. С. 171–195. 42. Носов A.M. Регуляция синтеза вторичных соединений в культуре клеток растений. // Биология кульивируемых клеток и биотехнология. М.: Наука, 1991. С. 5–19. 43. Попов А.С. Криоконсервация клеток растений. // Методы культивирования клеток. Л.: Наука, 1988. С. 70–77. 44. Родин А.Р., Калашникова Е.А. Использование методов клеточной и генной инженерии для получения посадочного материала древесных пород. Учебное пособие. М.: МГУЛ, 1993. 90 с. 45. Сельскохозяйственная биотехнология: Учеб. /В. С. Шевелуха, Е. А. Калашникова, С. В. Дегтярев и др.: Под ред. В. С. Шевелухи. М.: Высш. школа, 1998. 416 с. 46. Смоленская И.Н., Носов А.В. Методы получения и культивирования протопластов. // Методы культивирования клеток. Л.: Наука, 1988. С. 164–175. 47. Уайт Ф.Р. Культура растительных тканей. М.: Иностранная литература. 1949. 160 с. 48. Урманцева В.В. Культивирование каллусных тканей на твердых средах. // Методы культивирования клеток. Л.: Наука, 1988. С. 232–241. 49. Цуцаева А.А., Петренко Т.Ф. Криоконсервация культивируемых клеток животных. // Методы культивирования клеток. Л.: Наука, 1988. С. 63–69. 50. Шегебаев 0.0., Абдуллаев К.К., Жамбакин К. Ж. Генетико-битехнологические исследования в селекции пшеницы. // Биотехнология в селекции селькохозяйственных культур. Алматы, 1993. С. 28–48. 51. Эмбриология растений: использование в генетике, селекции, биотехнологии. М.: Агропромиздат, 1990. Т.2. 463 с. 52. Karabaev М., Shegebaev О. Cultivated cells of wheat and maiz: biotechnology and breeding. // Abstracts 5th International Wheat Conference June 10–14, Turkey, Ankara, 1996. P. 366. 53. Kisaka H., Kisaka М., Kanno A., Kameya T. Intergeneric somatic hybridization of rice (Oryza sativa L.) and barley (Hordeum vulgare L.) by protoplast fusion. // Plant Cell Reports. Vol. 17, № 5. 1998. P. 362–367. 54. Kumlehn J., Schieder О., Lorz H. In vitro development of wheat (Triticum aestivum L.) from zygote to plant via ovule culture. // Plant Cell Reports. Vol. 16. № 10.1997. P. 663–667.
Культуры животных клеток
КУЛЬТИВИРОВАНИЕ КЛЕТОК
История метода Признание идеи о том, что клетки тканей высших животных можно выделить из организма и затем создать условия для роста и воспроизводства их in vitro, датируется первым десятилетием XX века. После того как стало известно, что подобные процессы реальны, наступил второй этап работ, начало которому положила демонстрация возможности выращивания и репродукции в таких клетках фильтрующихся инфекционных агентов — вирусов. Третий этап истории начинается со времени, когда была показана практическая возможность получения в клетках животных больших количеств вирусного материала для применения в вакцинных препаратах, и простирается до времени, когда: 1) стало возможным вставить в клетки специфические экзогенно полученные гены и получить их экспрессию и 2) подтверждена возможность выращивания в культуре из одиночной клетки целой популяции. Когда такие популяции получали из клетки, выделявшей в окружающую среду антитела, то все молекулы антител в надосадочной жидкости были одинаковыми. Причины и следствия этих двух феноменов в настоящее время интенсивно исследуются и они знаменуют собой начало четвертого этапа работ в данной области. Чтобы показать способность клеток животных расти и делиться в культуре, потребовалось овладеть рядом подходов и методик. Особенности их приведены ниже. 1. Методики получения клеток, свободных от экзогенных прокариотов и грибов. 2. Методики разработки среды, в которых рост «вырезанных из ткани» или изолированных клеток не подавляется. 3. Методики наблюдения за клетками в динамике их развития. 4. Методики непрерывного культивирования культур клеток животных in vitro и поддержания их свободными от других биологических агентов. Научную основу для разработки этих методик составляет представление о клетке как основном структурном элементе живых организмов животного и растительного происхождения. Идея о том, что клетки тканей животных можно выделить из организма и затем создать условия для роста и воспроизводства их in vitro возникла на базе концепции, принадлежащей Клоду Бернару. Он предположил, что не только живые организмы способны сохранять постоянство внутренних условий, вне зависимости от изменений в окружающей среде. Клетка вне организма животного тоже будет стремиться поддерживать свои внутренние условия. Если различия между внутренними и внешними условиями будут незначительными, то высока вероятность роста и деления клетки. Такое понимание явления приводит к необходимости разработки сред, способных поддерживать и стимулировать рост клеток вне организма. Чуть позже, в 1885 году, У. Ру (W. Roux) показал возможность сохранения вне организма живых тканей на практике. Он сохранял в жизнеспособном состоянии оболочку куриного эмбриона в теплом физиологическом растворе. Впоследствии он стал автором, активно публиковавшимся по проблемам эмбриологии in vitro. Позднее, в 1897 г., Лёб (Loeb) поддерживал в жизнеспособном состоянии клетки крови и соединительной ткани в пробирках с сывороткой и плазмой крови. Льюнгрен (1898) показал возможность поддержания эксплантатов кожи человека в жизнеспособном состоянии в кислой среде с сохранением способности к реимплантации. Дополнительные эксперименты были проведены Джолли (1903), наблюдавшим деление клетки в висячей капле, содержащей лейкоциты саламандры, а Биб и Эвинг (1906) подтвердили это при пересадке лимфосаркомной ткани собаки. Продолжая работы Ру, Росс Харрисон усовершенствовал методику «висячей капли». Он использовал небольшие кусочки ткани, отторгнутые от медуллярного сосуда лягушки к внедренные в ее лимфатический тромб, и выдерживал их в виде капли на нижней стороне покровного стекла, расположенного поверх углубления в предметном стекле. В 1907 г. ему удалось наблюдать с помощью такой «камеры» рост нервных клеток в течение нескольких недель; он установил, что скорость роста этих клеток составляет 20 мкм за 25 мин. В то время как эксперименты Харрисона были направлены на то, чтобы получить ответы на вопросы, относящиеся к физиологии нервных клеток лягушки, методика, которой он пользовался, была применена Барроузом для других клеток тканей теплокровных животных. Этот исследователь в 1910 г. вместо лимфатического тромба использовал тромб плазмы курицы. В 1913 г. Алексис Каррель применил плазму крови, обогащенную экстрактом эмбриона. Добавка такого экстракта ускоряла рост тканей. Примененная методика обеспечивала значительно большую вероятность успеха, чем та, которую использовали Левис (1911) и Рид (1908 г.). Рид готовила культуры клеток из костного мозга морской свинки и пыталась выращивать эксплантаты на среде определенного химического состава. Работа Карреля привлекла большое внимание, так как она была опубликована под интригующим названием — культивирование «бессмертных» клеток. Инкубация клеток сердца куриною эмбриона была начала 17 января 1912 г. Пересев клеток продолжил Эблинг, как он сам заявлял, работая с ними 34 года. Поскольку Каррель был хирургом и весьма сведущим в вопросах асептики, он смог внести существенный вклад в культивирование клеток животных in vitro. В то же время организация и технические условия проводимых экспериментов были очень громоздкими. Ассистенты Карреля были одеты в длиннополые резиновые халаты темного цвета с капюшонами для полного прикрытия головы. Процедуры были длительными и отягощенными многими деталями. В результате тех требований, которые выдвигались автором в отношении сложных мер предосторожности для предотвращения контаминации, вокруг данного предмета создалась атмосфера таинственности и исключительности, что скорее тормозило прогресс, чем способствовало ему. Тем не менее, им было достигнуто многое. В частности, даже при отсутствии антибиотиков он добился успеха в пересадке клеток, используя хирургическую технику для отторжения отдельных колоний и переноса их в новые условия роста. Каррель также продемонстрировал своим коллегам научное значение тех наблюдений, которые могут быть сделаны в процессе пересадки клеток. В ходе проделанных работ был внесен ряд поправок в рецептуру среды культивирования. В частности, Тирод модифицировал раствор Рингера и в дополнение к куриной сыворотке и эмбриональному экстракту стал использовать коагулят фибрина. Для наблюдения за делящимися клетками животных Канти в 1928 г. разработал метод кинофотомикрографии. В этот же период был разработан дополнительный и очень существенный подход в технике работы с клетками. Имеется в виду применение трипсина для высвобождения клеток из тканевой матрицы, в которой они находятся. Однако эта методика не находила признания до тех пор, пока в 1937 г. Симмс и Стидлман использовали ее для пассирования клеток между культурами плазмы. Эта методика дает возможность успешно применять в культурах индивидуальные клетки, а не ткани. Впервые клоны клеток в культуре из одиночной клетки были получены Эрлом с сотрудниками в 1948 году. Игл (1955) систематически исследовал пищевые потребности клеток в условиях. До тех пор пока в 1961 г. Хейфлик и Мурхед не выделили линию диплоидных клеток человека (НДС) WI-38, считалось, что один раз установившаяся клеточная линия имеет неограниченное время жизни. Относительно линии WI-38 было показано, что период ее существования в культуре ограничивается приблизительно 50 удваиваниями популяции. Перед отмиранием популяции для клеток этой линии характерен феномен старения. Однако при отмирании эти клетки оставались диплоидными и не имели признаков злокачественных изменений. Клетки, выделенные из раковых опухолей или трансформированные в ходе культивирования, характеризуются «бессмертностью» и коррелируют с гетероплоидностью. Первые суспензионные культуры клеток животных, как правило, основывались на клетках злокачественных тканей. Это — клетки HeLa, выделенные из раковой опухоли шейки матки человека. Перевиваемая линия карциномы шейки матки была выделена еще в 1952 году Джеем с сотрудниками, она используется и в настоящее время во многих лабораториях мира. Последующий этап в истории культивирования диплоидных клеток человека связан с установлением факта, что они являются генетически стабильными и свободными от всех известных латентных и онкогенных вирусов. Поэтому линии диплоидных клеток человека разрешено применять для получения продуктов, предназначаемых для людей. Эта догма остается действующей и в настоящее время, хотя новейшие открытия отчетливо показали присутствие в клетках, выделенных из нормальных тканей, потенциальных онкогенов, идентичных тем, которые найдены в таких известных онкогенных вирусах, как вирус саркомы Рауса и вирус саркомы Молони. Раус еще в 1910 году индуцировал опухоль, использовав профильтрованный экстракт куриной опухоли. Эта опухоль была индуцирована РНК-вирусом (вирус саркомы Рауса). Позднее было установлено, что ряд вирусов способен индуцировать возникновение опухолей, такие вирусы были названы онкогенными.
Введение клеток в культуру, их происхождение В соответствии с целями и задачами экспериментальной работы можно выделить два направления культивирования животных клеток: — культуры клеток; — культуры органов и тканей (органные культуры). Культуры клеток лишены структурной организации, теряют характерную гистиотипическую архитектуру и связанные с ней биохимические признаки и обычно не достигают равновесного состояния при отсутствии специальных условий. Клетки в культурах размножаются, что обеспечивает получение большой массы клеток, затем их идентифицируют (по фенотипическим признакам, путем выращивания в селективной среде, генотипически), разделяют на идентичные параллели и, если это необходимо, сохраняют. Динамические свойства культивируемых клеток часто трудно контролировать, также трудно реконструировать in vitro некоторые клеточные взаимодействия, наблюдаемые in vivo. В связи с этим некоторые исследователи предпочитают использовать клеточные системы, сохраняющие структурную целостность исходной ткани. Список типов клеток, которые уже введены в культуру, достаточно велик. Это элементы соединительной ткани человека (фибробласты), скелетные ткани (кость и хрящи), скелетные, сердечные и гладкие мышцы, эпителиальные ткани (печень, легкие, почки и др.), клетки нервной системы, эндокринные клетки (надпочечники, гипофиз, клетки островков Лангерганса), меланоциты и различные опухолевые клетки. Популяция клеток не всегда гомогенна и обладает фиксированным фенотипом. Некоторые культуры, например, кератиноциты эпидермиса, содержат стволовые клетки, клетки-предшественники и кератинизированные чешуйчатые клетки. В такой культуре происходит постоянное обновление за счет стволовых клеток, пролиферация и созревание клеток-предшественников, а также необратимая дифференцировка, сопровождающаяся "слущиванием" чешуйчатых клеток в культуральную среду. Какую ткань лучше брать для введения в культуру, взрослую или эмбриональную, нормальную или опухолевую? Культуры, полученные из эмбриональных тканей, характеризуются лучшей выживаемостью и более активным ростом по сравнению с соответствующими зрелыми тканями. Причиной этого служит низкий уровень специализации и наличие реплицирующихся клеток-предшественников в эмбрионах. Пролиферативная способность взрослых тканей ниже, они содержат больше неделящихся специализированных клеток. Получение первичных культур клеток взрослых тканей и их размножение является более сложной задачей, продолжительность жизни таких культур, как правило, невелика. Нормальные ткани дают начало культурам с ограниченным временем жизни, тогда как культуры, полученные из опухолей, способны пролиферировать неограниченно долгое время. Дифференцировка нормальных клеток в культуре сопровождается обычно полным прекращением пролиферации клеток. В культурах опухолевых клеток возможна частичная дифференцировка при сохранении способности к пролиферации. Свежевыделенные культуры носят название первичных культур до начала пассирования или субкультивирования. Клетки первичной культуры обычно гетерогенны и характеризуются низкой пролиферацией. В них наиболее полно представлены типы клеток той ткани, откуда они были получены. Пассирование обеспечивает возможность продления существования культуры, возможность клонирования, исследования и сохранения свойств клеток. При этом получаются более однородные популяции, а также теряются специализированные клетки. После нескольких пересевов линия клеток либо гибнет, либо трансформируется и становится постоянной клеточной линией. Свойством "бессмертности" обладают в основном клетки, полученные из опухолей. Появление постоянной линии клеток констатируется по морфологическим изменениям (уменьшение размера клеток, снижение их адгезивности, округление, увеличение ядерно/цитоплазматического отношения, по увеличению скорости роста (время удвоения клеток в культуре снижается с 36–48 до 12–36 часов), по снижению зависимости от сыворотки, по увеличению эффективности клонирования, по снижению зависимости от субстрата, по увеличению гетероплоидности (хромосомные различия между клетками) и анеуплоидности и по увеличению опухолеродности. Нормальные клетки могут трансформироваться в постоянную линию, не становясь при этом злокачественными.
Характеристика клеток, культивируемых in vitro Клетки одного и того же типа в ткани взаимодействуют друг с другом и согласовывают скорость деления, чтобы поддерживать надлежащую плотность популяции. «Социальный» контроль такого рода четко проявляется при реакциях на повреждение. Например, когда поврежден эпителий, клетки по краям раны стимулируются к делению и наползанию на обнаженную поверхность до тех пор, пока она вновь не будет закрыта; в этот момент быстрая пролиферация и движение клеток прекращаются. Сходное явление можно наблюдать на диссоциированных клетках в культуре. Эпителиальные клетки или фибробласты, помещенные в чашку, в присутствии сыворотки будут «приклеиваться» к поверхности, распластываться и делиться до тех пор, пока не образуется сплошной монослой, в котором соседние клетки соприкасаются. Адгезивные контакты обеспечиваются образованием комплексов из поверхностных рецепторов мембраны клетки. В результате поперечного движения гликопротеидов в мембране образуются электронноплотные бляшки гликопротеиновых комплексов. Такие бляшки формируются в ответ на воздействие антител, агглютинирующих агентов (лектины) или соседних клеток. При адгезии субстрат действует как многовалентное антитело, а образующиеся бляшки называют «адгезивными пятнами». Эти пятна богаты «адгезивными белками» и всегда выделяют элементы цитоскелета, которые удерживают гликопротеиды. Благодаря такому действию уменьшается «разжижение» мембраны, и клетка предохраняется от округления. Образовавшиеся на клетке адгезивные пятна формируют выступы, с помощью которых и происходит перемещение. Выступы цитоплазмы (ложноножки, псевдоподии) при контакте с соседней мембраной ингибируют движение. Клетки в этом случае направляют свои псевдоподии в другом направлении (феномен контактного ингибирования). Когда культура станет монослойной, активность ложноножек и движение клеток прекращается. Нормальные клетки перестают делиться, это явление, известно как торможение пролиферации, зависимое от плотности. Если такой монослой «поранить» иглой таким образом, чтобы на чашке образовалась свободная от клеток полоска, клетки с краев этой полоски начинают продвигаться на свободное место и делиться. Вначале такие явления объясняли только контактным торможением клеточного деления, но это, видимо, не отражает сути дела. Плотность клеточной популяции, при которой клетки в сплошном монослое перестают делиться, увеличивается с повышением концентрации факторов роста в среде. Кроме того, оказалось, что если культуральная жидкость будет протекать по поверхности чашки с островками клеток, то клетки, омываемые средой, только что прошедшей над другими клетками, будут делиться медленнее, чем те, которые омываются средой, прошедшей над свободными от клеток участками. В среде, протекавшей над клетками, недостает каких-то важных питательных веществ или факторов роста. Фактор роста обычно присутствует в среде в концентрации около 10-10 М (примерно одна молекула в объеме сферы диаметром 3 мкм). Один фибробласт имеет около 105 рецепторов фактора роста, каждый из которых обладает очень высоким сродством к нему. Таким образом, у каждой клетки достаточно рецепторов, чтобы связать все молекулы ростовых факторов в объеме сферы диаметром около 150 Кроме того, полагают, что значительная часть фактора роста, связанного рецепторами клеточной поверхности, быстро поглощается путем эндоцитоза и разрушается. Из этого ясно, что соседние клетки конкурируют между собой за малейшие количества факторов роста. Такого рода конкуренция важна как для клеток в ткани, так и для культивируемых клеток, она предотвращает рост популяции выше некоторого уровня ее плотности. Конкуренция за факторы роста и питательные вещества не единственный фактор, влияющий на скорость деления в клеточной культуре. Форма клеток во время их распластывания и движения по поверхности субстрата на свободные места тоже сильно влияет на их способность делиться. При культивировании нормальных клеток в суспензии, когда они не прикреплены к твердой поверхности и поэтому имеют округлую форму, они почти никогда не делятся (зависимость деления от прикрепления). Влияние распластывания клеток на пролиферацию можно продемонстрировать при выращивании клеток на субстратах с различной адгезивностью поверхности или на таких субстратах, где имеются лишь крошечные адгезивные участки, на которых клетка может прикрепиться, но не может распластаться. Частота деления клеток возрастает с увеличением степени их распластывания. Возможно, что сильно распластанные клетки могут улавливать больше молекул фактора роста и поглощать больше питательных веществ благодаря своей большей поверхности. Однако некоторые типы клеток, почти не способные к пролиферации в суспензии, охотно делятся, как только им удается вступить в контакт с участком субстрата, даже если этот участок настолько мал, что клетка не может на нем распластаться. Такие «фокальные» контакты являются местами соединения (хотя и непрямого) внутриклеточных актиновых филаментов с молекулами внеклеточного матрикса. Эти и другие наблюдения определенно наводят на мысль, что контроль клеточного деления каким-то образом связан с организацией цитоскелета. Хотя механизм и функции этой связи не ясны, можно думать, что зависимость деления клеток от их прикрепления, вероятно, позволяет ткани сохранять целостность и предотвращает пролиферацию клеток, обособившихся от нормального окружения. Цикл прикрепления и открепления, вероятно, позволяет перегруппировать адгезивные контакты как между клетками, так и между клетками и матриксом, чтобы встроить вновь возникшие дочерние клетки в ткань, перед тем как они смогут начать следующий цикл деления. Ослабление контактов, видимо, составляет важную особенность пролиферативного поведения большинства типов клеток. Например, в ранней стадии реакции фибробластов на ростовой фактор отмечается разрушение их фокальных контактов. Потеря управляемости роста у раковых клеток почти всегда связана с необратимым уменьшением клеточной адгезивности, которое проявляется также в потере фокальных контактов при выращивании таких клеток в культуре. Изменение ростовых свойств культивируемых клеток называется трансформацией. Трансформация — процесс необратимый и, очевидно, включает генетические изменения в той специфической части наследственной информации, которая контролирует неопластический фенотип, добавляемый к трансформированному геному хозяина. Изменение ростовых свойств является одной из адаптивных особенностей, позволяющей клеткам пролиферировать в условиях, неблагоприятных для нетрансформированных клеток. Таким образом, в условиях, которые ограничивают рост нормальных клеток, трансформированные клетки будут расти до более высокой плотности популяции, что, по-видимому, будет связано с их пониженной потребностью в факторах роста. Существуют доказательства, что основное изменение при трансформации связано с изменением транспорта питательных веществ через клеточную мембрану, а это, в свою очередь, может сделать клетки менее зависимыми от «геометрических» факторов роста. Трансформированные клетки способны расти в условиях, в которых геометрические характеристики, а именно отношение площади поверхности к объему менее благоприятны. Следовательно, трансформированные клетки будут расти в суспензионных культурах, образуя сферические клоны. Так, трансформированные клетки, будучи введенными иммунологически толерантным животным в относительно небольших количествах, могут образовывать опухоли. По этой причине трансформация иногда приравнивается к злокачественным изменениям. Трансформация может быть или вирусной, или «спонтанной». Большинство исследований выполнены с клетками, подвергнутыми вирусной трансформации. Когда при этом использовали хорошо известные трансформирующие вирусы, такие, как вирусы SV40 и полиомы, то свойства трансформированных клеток проявлялись очень наглядно и были описаны подробно. Аналогичные изменения, наблюдавшиеся после спонтанной трансформации, привели к предположению, что такая трансформация является результатом активации последовательностей генов (онкогенов), уже присутствовавших в геноме трансформированной до этого клетки, сходной с таковой в вирусном геноме. Многие исследования подтвердили, что спонтанно трансформированные клетки имеют такую же последовательность оснований в ДНК, как и в клетках, трансформированных вирусами. Но в одной из работ было показано, что аналогичные (может быть, неразличимые) изменения вызываются также точечными мутациями в нормальных генах. Старение характерно для клеток, имеющих ограниченный потенциал пролиферации, то есть низкую плотность насыщения при идеальных условияхкультивирования. Примером могут служить линии диплоидных клеток человека. Повышенная способность к росту трансформированных клеток означает, что трансформация преобладает над процессом старения. Старение, безусловно, зависит от генетических факторов, так как каждый вид имеет характерную продолжительность жизни, но вариабельность внутри популяций по этому показателю свидетельствует также о влиянии фенотипа. При адаптировании диплоидных клеток человека (линия WI38) уже с самого начала было показано, что культивируемые клетки могут проявлять феномен старения и имеют ограниченное время жизни (50±10 удваиваний популяции). Зависящие от возраста изменения, которые при этом наблюдались, включали удлинение межмитотических интервалов (19±25 % — 31±41 %/час), изменение метаболизма, уровней ферментов и экспрессии продукта. Следует отметить, что не установлено четкой связи между продолжительностью жизни в зависимости от происхождения клетки (мышь, человек) и потенциалом удваивания их клеток в культуре. Ограниченные по продолжительности жизни клеточные линии, например линии диплоидных клеток фибробластов, не являются идеальными объектами для целей производства, поскольку их должны использовать до того, как в клетках произойдут серьезные изменения старения. В практических отношениях это означает, что период жизни этих клеток, когда их можно применять в целях производства, заключается между 12 и 30 пассажами (в первом случае — для приготовления посевного материала). Трансформированные же клетки не имеют ограниченной продолжительности жизни, и это обусловливает такое их преимущество в биотехнологии, как использование в качестве субстрата для генерации различных продуктов в сочетании с более высокими плотностями популяции клеток, более высокими скоростями роста и способностью расти в суспензиях.
Питательные среды и условия культивирования После извлечения клеток из ткани или организма и помещения их в культуру культуральная среда должна обеспечивать все внешние условия, которые клетки имели in vivo. Это обеспечивает выживание клеток, их пролиферацию и дифференцировку. Внеклеточная среда должна обеспечивать клетки питательными и гормональными факторами, т. е. обладать всем необходимым для роста и выживания клеток. Культуры клеток животных и человека предъявляют определенные требования к жидкой (питательная среда), газообразной (концентрация газов) и твердой (поверхность субстрата) фазе. Питательная среда представляет собой раствор определенного состава, к которому добавляются компоненты невыясненного биологического происхождения (добавки плазмы, сыворотки крови, тканевые экстракты и т. д.). Основу питательных сред составляют солевые растворы. Минеральные компоненты в этих растворах подобраны так, что раствор выполняет буферные функции, поддерживая постоянный кислотно-щелочной баланс среды в процессе культивирования. Постоянство pH среды является одним из главных требований условий культивирования. Для приготовления питательных сред обычно используются солевые растворы Эрла и Хенкса. Эти растворы, как и фосфатно-солевой буфер Дульбекко и Фогта используются также для орошения и промывки клеток при пассировании культур, выделении клеточных линий и других манипуляциях с культурами клеток. Другим важным условием культивирования является осмотическое давление. Оно определяется числом молей осмотически активных частиц (ионов и неионизированных молекул) растворенных веществ на 1 кг растворителя (осмоляльность) или на 1 литр раствора (осмолярность). В разбавленных водных растворах эти величины близки. Осмоляльность раствора[73] (осмоль/кг) = Smi*xi, где mi концентрация i-гo растворенного вещества (моль/кг), хi количество частиц, на которые диссоциировала его молекула. Например, для раствора Эрла расчетная величина осмоляльности равна 310.6 мосмоль/кг, реальная 283. Диапазоны pH и осмоляльности, при которых происходит размножение клеток, узки и варьируют в зависимости от типа клеток. Например, для клонального роста диплоидных фибробластов человека WI38 оптимальны рН = 7.30 + 0.15 и осмоляльность 285 + 40 мосмоль/кг, а для фибробластов из эмбриона цыпленка 7.12 + 0.18 и 300 + 20 соответственно. Для поддержания pH в большинстве сред используется бикарбонатный буфер: НСО3 = СО2 + ОН, если выделяется углекислый газ, увеличивается концентрация ОН. Растворы могут содержать малое количество бикарбонатного буфера (раствор Хенкса), они предназначены для поддержания pH в плотно закрытых сосудах. В других (растворе Эрла) бикарбоната больше, они используются в системах с повышенным парциальным давлением СО2. Если культуры ведутся вне СО2-инкубатора, где pH поддерживать труднее, необходимы альтернативные буферные системы. Хорошим буфером является HEPES 4-(2-оксиэтил)1-пиперазинэтансульфоновая кислота. HEPES легко растворим в воде, не связывает двухвалентные катионы, не цитотоксичен до концентрации 0.05 Моль. Применяется в концентрациях 0.01-0.03 М. Стандартные среды для ведения культур животных клеток. Среды Игла МЕМ (minimal essential medium) и ВМЕ (basal medium, Eagle). Чаще используется MEM. Она содержит минеральные вещества, аминокислоты (13 незаменимых), 6 водорастворимых витаминов, холин и инозит, выполняющие роль углеводородного субстрата. MEM используется только с сывороткой, так как в ней отсутствуют биотин, витамин В12, ионы железа и микроэлементы. Основа раствор Эрла. Среда Дульбекко DME или DMEM (двойная модификация среды Игла). Используется при культивировании клеток различных типов, в том числе нетрансформированных клеток и гибридом. Является основой для бессывороточных сред. Содержит двойную концентрацию аминокислот, глицин, серин, пируват, железо. При использовании этой среды необходим инкубатор с 10 % концентрацией СО2. Среда Искова IMDM — модификация среда Дульбекко. Добавлены незаменимые аминокислоты, биотин, витамин B12, селенит натрия. В среду введен HEPES и уменьшены концентрации NaCl и NаНСО3. Среда бессывороточная, обычно используется для культивирования лимфоцитов и кроветворных клеток. Среда МакКоя 5А и серия сред RPMI. Среда МакКоя 5А разработана в 1958 году для поддержания клонального роста клеток карциносаркомы Уолкера 256 в присутствии сыворотки, а затем уже других первичных культур и различных клеточных линий. Обычно производится в модификации Ивката и Грейса (RPMI) и предназначена для культивирования лейкоцитов в присутствии сыворотки, часто применяется и для культивирования гибридом. Концентрация СО2 в атмосфере при культивировании 5 %. Среда 199 разработана в 1950 году для культивирования фрагментов сердца из эмбриона цыпленка. Для среды характерны широкий спектр питательных веществ и невысокая их концентрация. Используется без добавок, как поддерживающая для первичных клеток, а с сывороткой как ростовая среда для быстро размножающихся клеток. Нормальные, сохраняющие специфические функции клетки на стандартных средах не размножаются (если не трансформированы). Для оптимального роста клеток обычно добавляют 5-20 % фетальной (эмбриональной) сыворотки. Сыворотка представляет собой чрезвычайно сложную смесь мелких и крупных молекул, способных как вызывать, так и тормозить рост клеток. К главным функциям сыворотки относятся: обеспечение гормональными факторами, стимулирующими рост клеток и их функции; обеспечение факторами прикрепления и распластывания клеток; обеспечение транспортными белками, переносящими гормоны, минеральные вещества, липиды и т. д. Белки сыворотки, прямо и специфически участвующие в стимуляции клеточного деления, называются факторы роста. Большинство ростовых факторов присутствуют в сыворотке в концентрации нескольких нанограммов на миллилитр и ниже. Некоторые из этих факторов специфичны для клеток на определенной стадии дифференцировки, действие других не ограничено каким-либо одним типом клеток. Один и тот же тип клеток может быть стимулирован различными ростовыми факторами. Например, фибробласты размножаются в ответ на фактор роста фибробластов, фактор роста эпидермиса, фактор роста, синтезируемый тромбоцитами и соматомедины. Все эти вещества являются митогенами (стимулируют митоз). Другим важным фактором роста практически для всех типов клеток является гормон инсулин. Из других гормонов наиболее часто применяются глюкокортикоиды (гидрокортизон, дексаметазон), стероиды (эстрадиол, тестостерон, прогестерон) и гормоны щитовидной железы (трииодтиронин). Гормоны стимулируют или подавляют рост в зависимости от типа клеток и их плотности. Глюкокортикоиды, например, влияют на пролиферацию клеток, изменяя их чувствительность к факторам роста. Для переноса низкомолекулярных факторов (витаминов, аминокислот, липидов и других) необходимы транспортные белки. В этой роли выступает альбумин. Транспорт железа обеспечивает трансферрин, а поверхность большинства культивируемых клеток содержит рецепторы для этого белка. К факторам прикрепления и распластывания клеток относятся коллаген и фибронектин, более специализированы хондронектин (адгезия хондроцитов) и ламинин (адгезия эпителиальных клеток). В последние годы разработаны бессывороточные среды для размножения клеток. Чаще всего эти среды узко специализированы, т. е. предназначены для определенного типа клеток. К базовой среде добавляется инсулин, трансферрин, гидрокортизон или его аналог дексаметазон и т. д. Бессывороточные среды имеют определенные преимущества: улучшение воспроизводимости результатов опыта вследствие большей стабильности состава среды; снижение риска заражения культуры вирусами, грибами, микоплазмой; облегчение очистки продуктов клеточного метаболизма; снижение влияния дополнительных белков на результаты биологических исследований; отсутствие цитотоксичности сыворотки. Культивирование клеток в присутствии сыворотки обнаруживает и ряд недостатков: для большинства тканей сыворотка не является физиологической жидкостью, с которой они контактировали в исходной ткани, поэтому, например, сыворотка вызывает рост фибробластов, но тормозит рост эпидермальных кератиноцитов; сыворотка может быть цитотоксичной, так как содержит полиаминоксидазу, действующую на полиамины (спермин, спермидин), являющиеся продуктами секреции быстро пролиферирующих клеток (эмбриональная сыворотка содержит относительно много такого фермента); значительная вариабельность состава сывороток разных партий; сыворотки могут содержать недостаточное количество специфических ростовых факторов, что вызывает необходимость добавления их к культурам клеток. Многие клетки млекопитающих, прежде чем приступить к пролиферации и образовать клеточный монослой, должны прикрепиться к субстрату и распластаться на нем. В связи с этим встает вопрос о подходящем материале. В качестве субстрата в настоящее время используют несколько материалов. Стекло лучше всего пирекс (алюмоборосиликатное стекло), так как натрийсиликатное стекло может подщелачивать среду и его необходимо кипятить в слабой кислоте перед употреблением. С каждым использованием пригодность такого стекла падает. Пластик — чаще всего используют полистирол, поликарбонат, поливинилхлорид, тефлон и другие. Металлы — подходит как нержавеющая сталь, так и титан, так как эти вещества химически инертны и обладают высоким отрицательным поверхностным зарядом. Клетки прикрепляются за счет электростатических взаимодействий, поэтому поверхность культуральных сосудов должна быть смачиваемой и отрицательно заряженной. Этого можно достичь химической обработкой окисляющими агентами или физическими воздействиями (высоковольтным разрядом, ультрафиолетовым светом, бомбардировкой высокоэнергетическими электронами). Фирмы, производящие пластиковую посуду, используют эти методы. Некоторые исследователи, несмотря на это, предпочитают даже новую посуду перед посадкой клеток обрабатывать смесью концентрированной серной кислоты и бихромата калия (хромовая смесь), после чего следует тщательная промывка. Иногда поверхность сосуда покрывают веществом, облегчающим прикрепление клеток. Наиболее часто для этого используют коллаген и полиаминокислоты.
Системы культивирования клеток Существует 2 основных системы культивирования клеток. 1. Непроточные культуры — тип культур, в котором клетки вводят в фиксированный объем среды. По мере роста клеток происходит использование питательных веществ и накопление метаболитов, поэтому среда должна периодически меняться, что приводит к изменению клеточного метаболизма, называемого еще и физиологической дифференцировкой. Со временем, в результате истощения среды происходит прекращение пролиферации клеток. Увеличить продолжительность жизни непроточных культур можно несколькими способами: — прерывистый (часть культуры заменяется равным объемом свежей среды); — постоянный (объем культуры увеличивается с постоянной низкой скоростью, а небольшие порции клеток периодически удаляются); — перфузионный (осуществляется постоянное поступление свежей среды в культуру и одновременное удаление равного объема использованной (бесклеточной) среды. Перфузия может быть открытой, когда из системы удаляется вся среда, и закрытой, когда удаляемая среда проходит через дополнительный сосуд, где восстанавливается ее pH и осуществляется аэрирование, и возвращается в культуральный сосуд. Все системы непроточных культур характеризуются накоплением отходов в той или иной форме и непостоянством внешних условий. 2. Проточные культуры обеспечивают истинные гомеостатические условия без изменения концентрации питательных веществ и метаболитов, а также числа клеток. Гомеостаз обусловлен постоянным вхождением среды в культуру и одновременным удалением равного объема среды с клетками. Такие системы пригодны для суспензионных культур и монослойных культур на микроносителях. Существует 2 крупных направления в культивировании животных клеток: монослойные культуры и суспензионные культуры. Суспензионные культуры предпочтительнее с точки зрения увеличения выхода клеток. Монослойные культуры также обладают рядом преимуществ: 1. Легко провести полную замену среды и промыть клетки перед добавлением свежей питательной среды. Это важно в тех случаях, когда рост клеток идет в одних условиях, а наработка продукта в других условиях, например при переносе клеток из среды с сывороткой в бессывороточную среду. Можно также полностью удалять нежелательные компоненты. 2. Позволяют обеспечить высокую плотность клеток. 3. У многих клеток экспрессия требуемого продукта идет эффективнее, если клетки прикреплены к субстрату. 4. Монослойные культуры могут быть использованы для любого типа клеток, что обеспечивает наибольшую гибкость исследований. 5. В некоторых случаях, например для распространения вирусов, требуются тесные межклеточные контакты. Недостатками монослойных культур являются: — требования большого пространства; — возрастание стоимости и трудоемкости при увеличении масштаба; — недостаточно эффективный контроль, обусловленный трудностями отбора пробы; — сложности в определении и контролировании pH, концентрации кислорода. Необходимо отметить, что применение микроносителей устраняет эти недостатки. Существует много различных разновидностей этого способа культивирования. Рассмотрим три основных направления (рис. 27):
1. Чашка Колле, плоский флакон с клетками на дне 2. Вращающаяся бутыль (круглый сосуд) с клетками на дне и стенках Колонка с клетками на микроносителях pH, давление, СO2, температура система контроля и регенерации среда компоненты питательной среды стеклянные бусы перистальтический насос 1. Культивирование в плоских флаконах (матрацах). 2. Культивирование во вращающихся бутылях, когда в каждый момент времени 1520 % поверхности бутыли покрыто питательной средой, а клетки находятся попеременно то в среде, то в воздухе. 3. Культивирование в колонках на микроносителях, в качестве которых выступают плотно упакованные, не смещающиеся стеклянные бусы диаметром 35 мм, стопка пластин и др., а питательная среда омывает их, протекая сверху вниз.
Использование культуры клеток человека Практически любые клетки человека могут быть введены в культуру и служить средством и объектом во многих медико-биологических исследованиях. Благодаря культивированию клеток возможности исследования и диагностики расширяются почти беспредельно, так как имеется возможность оценки не только морфологических и биохимических изменений, но и изменений в поведении клеток, их реакции на различные агенты, в том числе и на лекарственные воздействия. Кроме того, если в ряду поколений воспроизводится дефект, свойственный клеткам in vivo, значит это дефект наследственный. Наибольшее распространение получили культуры фибробластов. Широкое использование фибробластов для изучения патогенеза и диагностики наследственных болезней обусловлено не только легкостью их культивирования, но и тем, что соединительная ткань, главным клеточным элементом которой являются фибробласты, составляет значительную часть массы тела. Кроме того, фибробласты составляют строму многих органов, являются важными участниками их морфогенеза и создают условия микроокружения, необходимого для дифференцировки и функционирования специализированных клеток. В фибробластах имеется фермент моноаминоксидаза, изменения активности которого характерны для некоторых нервных и психических заболеваний. Фибробласты содержат рецепторы к глюкокортикоидным гормонам, инсулину, некоторым нейромедиаторам. Гринбергом в 1978 году была доказана возможность экстраполяции данных, полученных на культивируемых фибробластах, на условия in vivo. Во-первых, фибробласты in vitro сохраняют важнейшие черты, свойственные клеткам в организме, а также онтогенетические и индивидуально-генотипические свойства организмадонора. Во-вторых, не существует другого такого типа клеток, который в полной мере мог бы представлять свойства клеток организма. В-третьих, изменения, которые возникают при введении фибробластов в культуру, можно легко контролировать и свести к минимуму при создании соответствующих условий. Все вышеперечисленное также способствует использованию фибробластов для изучения клеточных, биохимических, молекулярных аспектов патогенеза ряда болезней, в том числе и связанных с наследственными дефектами нервной системы.
Культивирование клеток и тканей беспозвоночных Интерес к клеточным культурам беспозвоночных связан с разнообразием и оригинальностью роста и метаморфоза, которые могут быть объектом для изучения основных процессов клеточной дифференцировки и регуляции активности генов. С другой стороны, при рассмотрении способов получения энтомопатогенных препаратов отмечалось, что вирусы могут размножаться только при использовании живых клеток насекомых, в связи с чем, для получения вирусных препаратов необходимым условием являлось предварительное разведение насекомых-хозяев. Использование клеточных культур беспозвоночных позволяет решить эту проблему. Среды для культивирования клеток и тканей насекомых сильно варьируют по составу. При составлении сред используются данные по составу гемолимфы. Среды отличаются от сред для клеток и тканей млекопитающих наличием органических кислот, повышенным содержанием аминокислот и более высоким осмотическим давлением. Для получения культуры клеток и тканей беспозвоночных используют эмбрионы, имагинальные диски и органы насекомых: — имагинальные диски (зачатки взрослых органов насекомых) используют для изучения процессов дифференцировки in vitro; — эмбрионы с удаленной оболочкой используют для изучения начальных стадий развития насекомых; — отдельные органы для различных целей. Например, слюнные железы Diptera для изучения процессов пуффирования в политенных хромосомах (пуф вздутие хромосом при "включении" ДНК на транскрипцию, когда определенные участки ее раскручиваются и РНК-синтезирующие ферменты начинают синтез РНК; при линьке насекомых пуфы появляются в определенной последовательности).
КУЛЬТИВИРОВАНИЕ ОРГАНОВ
Органная культура — культивирование in vitro органа или части органа, в которых сохраняются анатомическая связь и функционирование тканей, максимально приближенные к таковым в условиях in vivo, то есть в организме. Миграция изолированных клеток на периферии экспланта подавляется специальными условиями культивирования, в результате чего могут даже образовываться дифференцированные структуры. Например, на периферии эксплантов легочной ткани развиваются новые мелкие бронхи, состоящие из альвеол, окаймленных бронхиальным эпителием. Органная культура сохраняет межклеточные взаимодействия, в течение долгого периода поддерживает гистологическую и гистохимическую дифференцировку, как правило, остается в не растущем состоянии в течение нескольких дней и даже недель. Эти культуры не способны к размножению. Ткани, зависимые от гормонов, сохраняют чувствительность к ним и характерные ответы, эндокринные органы продолжают секрецию специфических гормонов и т. д. Наибольшее сходство процессов морфогенеза in vivo и in vitro отмечено для эмбриональных тканей. Первые исследования в области культивирования органов и тканей относятся к концу прошлого века. Уже в 1897 году немецкий ученый Лёб (B.Loeb) опубликовал данные о культивировании фрагментов печени, почек, щитовидной железы и яичников кролика на небольших кровяных сгустках в культуральных пробирках. Дальнейшие исследования показали, что для предотвращения центральных некрозов в эксплантах пробирки должны быть заполнены кислородом. В результате многочисленных экспериментов было также установлено, что большинство органов или их фрагментов, за исключением кожи, растут на твердом субстрате лучше, чем в жидкой среде. Что же можно использовать в качестве субстрата? Существует несколько видов техники культивирования органов. В качестве субстрата можно использовать сгусток плазмы. Этот способ был предложен Феллом и Робинсоном и получил название "техника часового стекла", став классической техникой морфогенетического анализа эмбриональных органов (рис. 28).
Рис. 28. Метод часовых стекол (по Феллу и Робинсону, 1929)
Культивирование проводят во впадине часового стекла на поверхности сгустка, состоящего из плазмы цыпленка и эмбрионального экстракта кур. Часовое стекло помещают в чашку Петри и закрывают сверху влажной ватой или фильтровальной бумагой для предотвращения высыхания. Культивируют в термостате при 37,5 °C. Существуют модификации этого метода, при которых часовое стекло покрывается крышкой, приклеенной воском и другие. Недостатком метода, ограничивающим применение его в биологических исследованиях, является разжижение сгустка в окрестностях экспланта, который в результате оказывается в жидкости. Кроме того, сложный состав среды затрудняет проведение биохимических исследований. Эти недостатки устраняются при использовании сгустка агара. Такая техника была предложена Спраттом (рис. 29). Метод основан на получении агарового геля 1–4 % концентрации, основу которого составляют забуференные солевые растворы или питательные среды типа 199 с добавлением эмбриональной сыворотки.
Рис. 29. Метод часовых стекол с агаровым сгустком (по Вольффу и Хафену, 1952)
В середине 20-го века Чен обнаружил, что культуры можно выращивать на бумажных плотиках, плавающих на поверхности жидкости в часовом стекле. С целью улучшения техники позднее бумагу обрабатывали силиконом, комбинировали с миллипоровыми фильтрами, а затем перешли на плотики из ацетата вискозы. Этот материал хорошо растворяется в ацетоне, что облегчает подготовку ткани для гистологического анализа. Метод культивирования на плотиках не лишен недостатков, основной из них — погружение ткани в среду при затоплении плотика. Решение этой проблемы было предложено Троувеллом, который предложил культивировать органы на поверхности металлической сетки (рис. 30). Сетка представляет собой квадрат размерами 25*25 мм с отогнутыми краями, образующими четыре ножки высотой около 4 мм. Скелетные ткани культивируют непосредственно на сетке, тогда как мягкие вначале эксплантируются на бумагу, а затем помещаются на сетку.
Рис. 30. Модифицированный метод Троувелла (по И. Ласнитски, 1989)
В 1976 году, для длительного культивирования взрослых тканей человека, таких как эпителий бронхов и молочной железы, пищевод и др. был предложен метод поочередного культивирования в жидкой среде и газовой фазе. Для этого экспланты прикрепляются ко дну пластикового сосуда и покрываются средой. Сосуды помещают в камеру с определенным газовым составом, а камера помещается на качающуюся платформу.
ГИБРИДИЗАЦИЯ ЖИВОТНЫХ КЛЕТОК
История метода Предположение о том, что соматические клетки могут сливаться друг с другом, было высказано еще в начале XIX века в связи с открытие многоядерных клеток. В историческом аспекте представляет интерес то обстоятельство, что открытие поликарионов как бы подтверждало ошибочное представление Шлейдена, который считал, что новые клетки зарождаются в виде пузырьков внутри цитоплазматической мембраны родительских клеток. Разделявший эту точку зрения Рудольф Вир хов представил в 1851 г. рисунок многоядерной опухолевой клетки в полной уверенности, что ядра являются эндогенными зачатками новых клеток. Кроме того, открытие поликарионов подлило масла в огонь борьбы с клеточной теорией. Противники ее выдвинули гипотезу, согласно которой организм представляет собой единую тканевую массу с непрерывной цитоплазмой, а существование поликарионов рассматривали как подтверждение этой гипотезы. Со временем восторжествовала клеточная теория, а существование поликарионов отнесли к разряду интересных исключений. Гибриды соматических клеток были открыты лишь в 60-х годах нашего столетия. В 1960 г. Барский с сотрудниками сообщили о выделении линии гибридных клеток. Гибридные клетки были получены путем смешения двух линий, выделенных ранее из 1 клетки мышиной саркомы. Исходные линии отличались по числу и морфологии хромосом, а также по способности к образованию опухоли при введении их мышам. Гибридные клетки содержали число хромосом, отличное от исходных клеточных линий, а также содержали поверхностные антигены клеток обеих родительских линий. Далее было установлено, что клеточные гибриды можно получить, используя клетки различных видов животных. В качестве агента, индуцирующего слияние, выступал инактивированный вирус HVJ, называемый также вирусом Сендай. С этих пор вирус Сендай стал широко использоваться в экспериментах по слиянию клеток. При изучении межвидовых гибридных клеток, способных к пролиферации были сделаны два очень важных наблюдения: — в гибридах могут проявиться оба генома; — в долгоживущих межвидовых гибридах элиминируются хромосомы одного вида. Слияние клеток не обязательно должно быть чем-то стимулировано. Как in vivo, так и in vitro оно может проходить и без добавления специальных агентов. Несмотря на то, что все слияния такого рода можно считать спонтанными, некоторые из них постоянно происходят в процессе онтогенеза, а следовательно, эволюционно запрограммированы. До сих пор нерешенной остается одна из труднейших загадок биологии, состоящая в том, что мембраны, находящиеся внутри клетки сливаются часто, тогда как мембраны, разграничивающие клетки, сливаются редко. Например, пузырьки аппарата Гольджи сливаются друг с другом, образуя клеточные пластинки при цитокинезе у растений, мембраны ЭР — с элементами АГ при переносе вновь синтезированных белков и т. д. В то же время нормальные клетки в естественных условиях крайне редко сливаются друг с другом. Исключение составляет процесс оплодотворения. Кроме того, в качестве подобного рода исключения выступает процесс плазмогамии у высших грибов, когда одноядерные гаплоидные клетки сливаются, образуя двуядерные (дикарионы). Такие клетки размножаются митотически, оставаясь двуядерными, и в результате образуют всем хорошо известные плодовые тела. В естественных условиях слияние клеток происходит и у млекопитающих. Например, клетки могут сливаться при формировании мышечных трубочек. Еще в XIX веке было показано, что миофибриллы поперечно-полосатых мышц образуются в поликарионах — крупных удлиненных многоядерных клетках. Поликарионы — продукт слияния одноядерных миобластов. Слияние опухолевых клеток — довольно обычное явление, при этом опухолевые клетки in vivo иногда сливаются и с нормальными. Эксперименты по спонтанному слиянию клеток проводились и in vitro. При проведении подобных экспериментов получают так называемых "химерных" или аллофеных мышей — животных, в тканях которых содержатся клетки различных генотипов (рис. 31).
Рис. 31. Получение аллофенных мышей
Методы создания химер 1. Агрегационный — был предложен практически одновременно и независимо друг от друга Тарковским в Варшаве и Минц в Филадельфии (1961–1962 гг.). Из матки беременных самок-докторов извлекают зародыши, достигшие стадии 8 бластомеров. Бластомеры, полученные от двух животных с различными генотипами (например, от мышей с белой черной окраской шерсти) помещают в условия, способствующие их агрегации и образованию 16-ти клеточного зародыша. Такие составные зародыши развиваются in vitro до стадии бластоцисты, после чего их вводят в матку приемной матери, у которой предварительно вызывают ложную беременность путем введения соответствующих гормонов. В результате получаются аллофенные мышата. Когда у мышонка появляется шерсть, окраска у него оказывается не белой или черной, как у родителей, а смешанной, с чередующимися черными и белыми пятнами или полосами. Это доказывает, что ткани животных-химер мозаичны, т. е. состоят из "белых" и "черных" клеток. Внутренние ткани таких животных, естественно, также мозаичны, хотя это проявляется не так очевидно, как в случае окраски шерсти. Различия могут касаться белков, выполняющих ферментативную функцию: они могут катализировать одни и те же реакции у мышей-родителей, нуждаться в одних и тех же кофакторах, но при этом быть не идентичными, хотя и сходными. Такие белки называются изоферментами, и их можно разделить с помощью электрофореза. Агрегационные химеры можно получать не только между двумя эмбрионами, но и между различным числом изолированных бластомеров или отдельными частями эмбрионов. Масса химерных эмбрионов не больше обычной и подвержена действию механизмов эмбриональной регуляции. Преимущество метода — не требует вмешательства микрохирургической техники, поэтому широко используется в эмбриогенетике. 2. Инъекционный — был разработан Р. Гарднером в 1968 г. Используются эмбрионы на стадии бластоцисты. Бластоцисту фиксируют и, используя микроманипуляторы, вводят путем инъекции клетки внутриклеточной массы бластоцисты доноров в бластоцель эмбриона — рецепиента. Этим методом можно инъецировать не только внутриклеточную массу ранних эмбрионов, но и более дифференцированные клетки. Инъекционный метод нашел применение при получении межвидовых химер. Первые межвидовые химеры были получены между двумя ближайшими видами мышей, которые обычно не скрещиваются: М. muskulus и М. caroli. Причем было отмечено, что химерные эмбрионы, полученные инъекционным методом, нормально развивались только при пересадке их в матку того вида, чья бластоциста была использована в качестве рецепиента. Например, в бластоцисту М. muskulus вводили внутриклеточную массу эмбриона М. caroli. Полученные химеры имплантировались в матку М. muskulus и благополучно развивались там, а в организме М. caroli погибали спустя две недели. Межвидовые химерные зародыши между мышью и крысой путем агрегации были получены только в 70-х годах. Первые химерные животные были получены только в 1973 году Р. Гарднером и М. Джонсоном. Успех этих экспериментов позволил приступить в 80-х годах к созданию химерных сельскохозяйственных животных. Выяснилось, что агрегационный метод неприемлем для получения химер крупного рогатого скота. Химер телят Bos indicus + Bos taurus удалось получить только инъекционным методом. В 1984 году были получены межвидовые химеры между овцой и козой — овцекозы, причем практически одновременно в Англии и ФРГ. Использовались оба метода. Половым путем овцы и козы не скрещиваются, так как имеют разный набор хромосом: коза 2n = 60, овца 2n = 54. В ФРГ в 1985 году были получены химерные телята после агрегации половинок 32-клеточных эмбрионов от коров швицкой (бурой) и голштино — фризской пород. В фенотипе химер сочетались обе масти — бурая и черно — пестрая. Химерные животные не передают потомкам генетическую мозаичность. У них происходит расщепление, как у гетерозигот, поэтому ценные генетические комбинации нарушаются. Но на протяжении 1 поколения хозяйственно ценные признаки поддерживаются, поэтому можно, например, сочетать как молочную, так и мясную продуктивность. Химерность довольно часто встречается и у растений. Как правило, она существует в скрытом виде, не проявляясь фенотипически. Однако пластидные мутации позволяют увидеть ее непосредственно на растении. Чаще всего спонтанная химерность наблюдается у гетерозиготных растений. Различные клеточные типы четко разделены в пространстве, образуя отдельные слои при делении апикальных меристем. Примером видимой мутации хлоропластов и образования химерного растения является пестролистность или появление секторов ткани другого цвета.
Механизм слияния клеток Для индукции слияния клеток используются вещества различной природы. Ионы Са2+, полиэтиленгликоль, лизолецитин, моноолеат глицерина, вирус Сендай. Лизолецитин — поверхностно-активное вещество липидной природы, продукт деградации лецитина путем обработки последнего фосфолипазой А. Лизолецитин повреждает мембраны и токсичен для живых систем. Цитотоксический эффект этого вещества можно уменьшить снижая его концентрацию или добавляя во время обработки альбумин. Моноолеат глицерина также соединение липидной природы, но его повреждающее действие менее выражено, а частота слияния клеток при применении этого вещества возрастает в 4–7 раз по сравнению со спонтанным процессом. К другим агглютинирующим агентам, способность которых вызывать слияние клеток, была исследована специально, относятся лектины растений и антитела. Преимущество вируса Сендай как сливающего агента заключается в полном отсутствии цитотоксического эффекта. Вирус перед употреблением инактивируют, облучая ультрафиолетовой лампой в течение 5 минут, при этом он теряет способность к размножению, но сохраняет способность сливать клетки. Вирус Сендей имеет два недостатка: — необходимость наращивать, титровать, концентрировать и инактивировать вирус; — клетки растений и грибов не имеют рецепторов к этому вирусу, поэтому он неприменим для их гибридизации. Первый этап слияния (рис. 32) — сближение мембран соседних клеток и уста новление между ними тесного контакта. Мембраны должны быть приближены друг к другу на расстояние в несколько ангстрем так, чтобы между ними стали возможны взаимодействия, подобные гидрофобным связям. Вызывают подобное сближение агенты, индуцирующие агглютинацию клеток. Миксовирусы, например, вирус Сендай, наряду с другими вирусами, которые не обусловливают слияния, прежде всего, вызывают агглютинацию клеток, т. е. достаточно тесное их сближение, необходимое для успешного последующего слияния.
Рис. 32. Этапы слияния клеток (по Н. Рингертцу, Р. Сэвиджу, 1979): А — 1-й этап, сближение цитоплазматических мембран: 1 — гликопротеиды, 2 — липиды, 3 — плазматические мембраны, 4 — митохондрии, 5 — микрофиламенты, 6 — частицы вируса Сендай; Б — 2-й этап, выход гликопротеидов и обнажение липидных слоев мембраны; В — 3-й этап, образование мицелл; Г — начало 4-го этапа, слияние мембран, образование цитоплазматических мостиков.
Полиэтиленгликоль также вызывает агрегацию клеток, хотя механизм действия его неизвестен. Возможно, благодаря тому, что в водном растворе ПЭГ несет небольшой отрицательный заряд, молекулы этого размера достаточно велики, чтобы между клетками возникали электростатические связи. Подтверждением этой гипотезы является усиление агглютинации клеток, вызываемой ПЭГ: двухвалентные ионы, по-видимому, образуют мостики между ПЭГ и отрицательно заряженными углеводами, находящимися на клеточной поверхности. Согласно другой гипотезе, протопласты сливаются в результате дегидратации. Поглощение воды индуцирует образование пор на поверхности мембраны и происходит перетекание внутриклеточного материала. После слияния участки с порами сохраняются некоторое время. Существует два предположения, объясняющие возникновение пор: 1. При высокой концентрации ПЭГ (20–30 %) вся свободная вода поглощается им, вызывая разрывы в мембране; 2. ПЭГ уменьшает полярность воды, что вызывает перераспределение полярных и гидрофобных компонентов мембраны, стабилизирующих липидные слои. С особым успехом для этих целей используется ПЭГ с молекулярным весом от 1500 до 7500. Лектины и антитела — двух- или поливалентные соединения, и их агглютинирующая функция связана со способностью одной молекулы какого-либо из этих соединений взаимодействовать с рецепторами, находящимися на поверхности двух клеток, что и приводит к образованию связи между клетками. Достаточное количество таких молекулярных связей может удерживать клетки вместе, препятствуя их расхождению либо в результате броуновского движения, либо в результате электростатического отталкивания, либо в результате активной миграции клеток. На втором этапе гликопротеиды, расположенные на поверхности, начинают высвобождаться из участков мембраны, лежащих между вирионами, и притягиваются к местам прикрепления вирусных частиц. Углеводные компоненты — наименее изученная часть клеточной поверхности. В мембранах они встречаются в виде нейтральных сахаров, а также ковалентно связываются с липидами или белками. Именно в таком виде они активно участвуют во многих биологических процессах. Установлено, что гликопротеиды определяют антигенную специфичность клеток, несут отрицательный заряд, характерный для клеток при физиологически нейтральных pH, принимают участие в распознавании и адгезии клеток, определяют рецепторные участки для вирусов, бактерий, агглютинирующих агентов, принимают участие в процессе регуляции проницаемости мембран для ионов. Эти свойства и определяют их участие в слиянии клеток. Углеводы препятствуют слиянию клеток, так как разделяют липидные слои мембран, не давая им соприкасаться, так как на поверхности нормальной клетки рецепторные участки расположены редко или поодиночке. В результате трансформации вирусом происходит их объединение в группы. Гликопротеиды мигрируют в мембране к месту адсорбции вируса, оставляя свободными соседние участки, где и происходит слияние клеток. В некоторых случаях в удалении углеводных групп, препятствующих слиянию клеток, принимают участие лизосомные ферменты. При этом происходит соединение лизосом или пузырьков АГ с плазматической мембраной, и локальное высвобождение гликозидаз. Такая лизосомная активность может быть запрограммирована (в гаметах, миобластах, макрофагах, т. е. клетках, сливающихся в естественных условиях) или вызвана экспериментально при индуцированном слиянии клеток. Третий этап — мицелизация обнажившихся липидов двух противолежащих мембран. Мицеллы — липидные капли, где молекулы липидов гидрофильными головками обращены к воде, а гидрофобные "хвосты" жирных кислот спрятаны внутрь. Этот процесс усиливается при высоких значениях pH и высокой концентрации ионов кальция. В обеих клетках начинается эндоцитоз вирусных частиц. Четвертый этап — слияние мембран. Благодаря мостикам, образуемым Са2+, размер мицелл уменьшается. Под действием АТФ и Са2+, активизируются микрофиламенты. Возникший цитоплазматический мостик стабилизируется и расширяется благодаря функционированию микрофиламентов. На последних этапах слияния клеток важную роль играет наличие АТФ. АТФ образуется благодаря деятельности митохондрий. Поликарионы, образующиеся при слиянии двух или трех клеток, представляют собой не бесформенные раздутые или двулопастные шары цитоплазмы, а принимают форму, характерную для одной из родительских клеток или промежуточную между ними. Форму клетки определяют микротрубочки и микрофиламенты. В процессе слияния клеток микрофиламенты актиномиозинового типа, расположенные под цитоплазматической мембраной, принимают непосредственное участие, так как обеспечивают объединение цитоплазм слившихся клеток и стабилизацию вновь образованной системы. Однако, формирование цитоскелета — энергозависимый процесс, требующий большого количества АТФ. Видимо, этим обусловлено присутствие митохондрий в местах слияния клеточных мембран.
МОНОКЛОНАЛЬНЫЕ АНТИТЕЛА
Функциональная структура антител Слияние клеток лежит в основе получения гибридных клеток, продуцирующих антитела. Антитела — белки сыворотки крови, которые синтезируются в организме как проявление защитной реакции при попадании в него чужеродного вещества (антигена). Иммунная система вырабатывает специфические антитела на огромное множество антигенов. В основе такой способности лежит наличие большого многообразия клонов лимфоцитов,каждый из которых вырабатывает антитела с узкой специфичностью. В совокупности называемые иммуноглобулинами (Ig), антитела составляют один из главных белковых компонентов крови — по весу около 20 % суммарного белка плазмы. В качестве антигенов выступают различные вещества: клетки микроорганизмов, вирусы, белки, нуклеиновые кислоты, в некоторых случаях низкомолекулярные вещества типа антибиотиков или пестицидов. Антитела образуются не против всей молекулы белка или бактериальной клетки, а только к небольшим участкам на их поверхности, получивших название антигенных детерминант. В случае белковой молекулы антигенном детерминантом являются участки поверхности, содержащие около 5 аминокислотных остатков. Простейшие молекулы антител имеют форму буквы Y с двумя идентичными антиген-связывающими участками — по одному на конце каждой из двух "ветвей". Связывание антигенных детерминант приводит к потере определенных функций молекулы или клетки, на чем и основан защитный механизм действия антител. Поскольку участков два, они могут сшивать антигены:
Если молекула антигена имеет три или большее число антигенных детерминант, то антитела могут сшивать их в обширную сеть. Достигнув определенных размеров, такая сеть может выпасть из раствора в осадок:
Тенденция больших иммунных комплексов к осаждению (преципитации) удобна для выявления антител и антигенов. Образование таких комплексов может приводить к агглютинации (склеиванию) молекул. Это явление лежит в основе определения групп крови, когда эритроциты склеиваются антителами той или иной специфичности — реакция гемагглютинации. Молекула антитела образована четырьмя полипептидными цепями (рис. 33).
Рис. 33. Строение антитела
Две из них — идентичные легкие (L-цепь, из 220 аминокислот), а две — тяжелые (Н-цепь, из 440 аминокислот). Все четыре цепи соединены между собой нековалентными и ковалентными (дисульфидами) связями. Антиген-связывающие участки образуются за счет одной Н и одной L-цепи. Эффективность реакций связывания антигена возрастает благодаря гибкому шарнирному участку антитела, который позволяет изменять расстояние между двумя антиген-связывающими участками. Шарнирный участок находится на Н-цепи. Н-цепь образует также "хвостовой" участок молекулы, который содержит также одну или несколько олигосахаридных цепочек, функция которых неясна. Как L, так и Н-цепь построены из повторяющихся сегментов, или доменов, каждый из которых сворачивается независимо, образуя компактную функциональную единицу (эпитоп). Эти участки также могут выступать в качестве антигенных детерминант и, соответственно, связываться другими антителами.
Получение моноклональных антител Как образуются антитела? Иммунный ответ — сложный процесс межклеточных взаимодействий различных типов лимфоидных клеток с участием специальных гормонов, в результате чего В-лимфоциты начинают активно синтезировать и выделять в кровь специфические антитела против данного антигена. На поверхности В-лимфоцитов существуют рецепторы, аналогичные антителам, взаимодействие которых с антигеном в сложном межклеточном комплексе служит стимулом для начала биосинтеза антител. Получение антител для нужд человека начинается с иммунизации животных. После нескольких инъекций антигена в присутствии стимуляторов иммунного ответа в сыворотке крови накапливаются специфические антитела. Антитела выделяют из сыворотки в виде g-глобулиновой фракции, осаждая сыворотку крови сульфатом аммония, спиртом, ПЭГ и другими веществами. Полученные антитела содержат много примесных белков. Высокоочищенные антитела выделяют с помощью ионообменной хроматографии. Стандартные препараты получить довольно сложно, так как состав их зависит от вида животного, его индивидуальных особенностей, цикла иммунизации, других малоконтролируемых факторов. В то же время, для современного биохимического анализа очень важна специфичность, то есть способность выделить данное вещество в сложных многокомпонентных средах, таких, как сыворотки крови, сок растений, ферментная среда. Такое возможно при использовании иммунохимического метода, использующего антитела, взаимодействующие узко специфично по принципу "антиген — антитело". Для проведения такого анализа необходимы абсолютно идентичные антитела, синтез которых обычными способами неприемлим. Решение проблемы было предложено в 1975 году английскими учеными Георгом Кёлером и Цезарем Мильштейном. Они разработали методику получения клеточных гибридов — гибридом. Гибридомы образуются в результате слияния лимфоцитов, взятых от иммунизированных животных, с клетками миеломы костного мозга, культивируемыми in vitro. Животное иммунизируют, в ответ на введение антигена в организме мыши активизируются продуцирующие антитела В-лимфоциты. Эти клетки могут жить только в организме хозяина, при переводе на искусственную питательную среду они гибнут. Если слить иммунную клетку с опухолевой, образуются гибридные клетки, способные неограниченно долго жить в искусственных средах. Одновременно они сохраняют способность синтезировать антитела. Гибридомы, синтезирующие определенные виды антител, отбирают на селективных ростовых средах. Затем их помещают в культуральную жидкость, в которой они размножаются и образуют много родственных клеток (клон). Такие клоны могут синтезировать антитела, получившие название моноклональных (МКА). МКА — антитела, однородные по структуре и специфичности, которые можно производить в неограниченных количествах. Другой метод получения антител основан на инъекции полученной гибридомы в брюшную полость мышки. Там гибридома реплицируется и вызывает образование асцитной опухоли (скопления клеток, плавающих в жидкости, заполняющей брюшную полость). Асцитная жидкость, выделенная из этой мыши, представляет суспензию, содержащую антитела. Клетки и белки, не относящиеся к МКА, удаляются. Оставшийся материал, представленный преимущественно антителами, используют. Этот метод позволяет получать высококонцентрированные препараты антител. Но массовое производство требует одновременного использования нескольких тысяч мышей. Кроме того, получаемый материал требует доочистки. Это дорого и трудоемко, поэтому в настоящее время предпочтение отдается первому способу, с использованием культуры клеток.
Методы анализа на основе моноклональных антител Высокая специфичность антител в отношении антигена превращает их в мощный инструмент для идентификации различных веществ, будь то макромолекулы, клеточные фрагменты или целые клетки. Начало широкому использованию антител в диагностических целях положил в 1955 году американский иммунолог А. Кунс. Он присоединил к антителам светящийся краситель. Флюоресцирующие антитела сделали видимыми места расположения интересующих его молекул в клетке. Этот метод получил название иммунофлюоресцентного. Чувствительность метода можно повысить несколькими путями. В первом случае иммунный ответ усиливается за счет применения антител нескольких порядков:
Антиген иммобилизуется на подложке, к нему добавляются антитела 1-го порядка, связывающиеся непосредственно с антигеном. В исследуемый образец добавляются антитела 2-го порядка, связывающиеся с антигенными детерминантами антител 1-го порядка. Антитела 2-го порядка имеют флюоресцирующую (или другую) метку. Поскольку участков связывания может быть несколько, то реакция проявляется более отчетливо. Другая система усиления сигнала основана на высоком сродстве биотина (низкомолекулярного растворимого витамина) к стрептавидину (бактериальному белку). Здесь возможны два варианта: А. Если возможно ковалентно связать биотин непосредственно с антителами, то стрептавидин метят маркером и используют аналогично антителам второго порядка:
Б. Система "биотин-антитело + стрептавидин + меченый биотин":
В этом случае образуется целая сеть из молекул стрептавидина, связанного с меченым биотином. Следовательно, происходит многократное усиление сигнала. Применение антител второго и третьего порядков позволяет также упрощать процедуру определения микроорганизмов в мазке. При этом не обязательно иметь меченые антитела против всех бактерий. Достаточно иметь обычные антитела кролика или мыши против интересующего микроорганизма и меченые МКА против этих иммуноглобулинов. Если микроорганизм в мазке присутствует, то к нему "приклеятся" специфические антитела, а к ним уже — меченые. В результате мазок будет светится при люминесцентной микроскопии. Фотометрические или флуоресцентные методы могут быть использованы не во всех случаях, например, если измерение проводят очень мутной среде. Кроме красителя в качестве метки можно использовать фермент (иммуноферментный анализ) или радиоактивный изотоп (иммунорадиологический). От чувствительности детекции маркера зависит чувствительность метода анализа.
Радиоактивные метки. Выбор маркера и способа его «привязки» к антигену является одним из важных этапов в проведении анализа. Первоначально широко применялись радиоизотопные метки (радиоиммунный анализ — РИА), предложенные американскими исследователями (С. А. Берсон, Р. С. Ялоу, 1959). Однако в последние годы все более широкое использование в качестве маркеров находят ферменты. Это обусловлено рядом принципиальных трудностей, связанных с применением изотопных маркеров. Так, изотоп 125I имеет время полураспада 60 суток, чем ограничивается срок его использования. Изотоп 3Н имеет длительное время жизни (12.5 лет), однако под антитело + биотин действием бэта-излучения происходит распад молекул антигена, в результате чего время жизни меченых 3Н-соединений тоже ограничено. Кроме того, эффективность счета трития существенно ниже, чем 125I. Ограничивающими факторами РИА являются сложность и высокая стоимость оборудования, необходимость централизованной системы распределения иммунохимических наборов, меченных радиоактивными изотопами, определенная опасность изотопов для окружающей среды. Учитывая трудности использования радиоизотопных меток, были предложены в качестве маркеров ферменты. При иммуноферментном анализе антиген связывается с поверхностью лунки полистирольного планшета. В лунку добавляют антитела, несущие фермент в качестве метки, инкубируют и отмывают. Далее приливают субстрат, который меняет окраску при взаимодействии с этим ферментом. Изменение окраски можно измерить с помощью спектрофотометрии. Таким способом проводится индикация и количественная оценка биоорганических соединений с чувствительностью до 10-12 г/литр. В настоящее время известно более 2000 разных ферментов, однако, только некоторые находят применение в иммуноферментном анализе. Это объясняется высокими требованиями, предъявляемыми к свойствам ферментов. Фермент должен быть высоко активен, а продукты его реакции детектироваться с высокой чувствительностью, он должен быть стабилен, так чтобы его активность сохранялась не менее одного года. Содержание фермента-маркера в определяемом образце должно быть минимальным. Именно из-за этого для разных объектов используют разные ферменты. Во многих случаях, когда необходим качественный результат, оценка иммунохимической реакции может быть проведена визуально. Для введения ферментативной метки разработано много разных химических, биохимических и иммунологических способов. Первым реагентом, использованным для синтеза иммуноферментных конъюгатов, был глутаровый альдегид, реагирующий с аминогруппами лизина белковых молекул. С помощью глутарового альдегида получены конъюгаты антител и антигенов с пероксидазой, щелочной фосфатазой, глюкоамилазой. В настоящее время широко используются иммунопероксидазные конъюгаты и конъюгаты с бэта-галактозидазой. Ковалентные методы получения иммуноферментных конъюгатов нашли весьма широкое распространение, однако, к некоторых случаях действие сшивающего реагента отрицательно сказывается на ферментативной и иммунологической активности компонентов гибридной макромолекулы. В связи с этим определенный интерес представляют иммунологические методы введения ферментной метки. Один из подходов получил название метода «гибридных антител». Ферментативным гидролизом получают Fab-фрагменты молекул антител против определяемого антигена и используемого фермента. Затем смесь продуктов гидролиза подвергают восстановлению меркаптоэтанодом; при этом Fab-фрагменты обратимо диссоциируют на симметричные части. После удаления восстанавливающего агента молекулы снова ассоциируют, образуя гибридные молекулы антител, специфичные к определяемому антигену и ферменту. При добавлении фермента образуется комплекс антитело — фермент (рис. 19,а). Гибридомная технология открывает принципиально новый путь получения гибридных антител, который заключается в том, что сливаются моноклональные клетки, специфичные против данного антигена и фермента-маркера, в результате чего образуются гибридомы второго поколения, синтезирующие антитела, с двумя специфичностями. Другой путь заключается в том, что получают антитела одного и того же вида животного (например, кролика) против определяемого антигена и фермента, которые соединяют между собой через антитела другого вида животных (антитела барана против кролика). Добавление фермента к такой тройной молекуле также приводит к образованию комплекса антитело — фермент. В настоящее время разрабатываются подходы получения гибридных антител методами клеточной и генной инженерии, что позволит существенно упростить способ их получения. Стабильность иммуноферментных конъюгатов при хранении — важнейший параметр, обусловливающий возможность их практического использования. Методы направленной стабилизации конъюгатов пока еще не разработаны. Не существует также корреляции между стабильностью конъюгатов и методом их получения. Однако высокая стабильность гибридных молекул обеспечивает их применение на практике и значительно превосходит стабильность антител и антигенов, меченных радиоактивными изотопами. В лиофилизованном состоянии ферментные конъюгаты сохраняют свои свойства до двух лет. Кроме ферментов в качестве маркеров могут быть использованы субстраты. В частности, в иммунокофакторном анализе применяются в качестве меток АТФ и НАД, которые могут быть «пришиты» к молекуле антигена через адениновый остаток таким образом, что сохраняется их способность взаимодействовать с ферментом. Аналогично были использованы субстраты пероксидазы (люминол, изолюминол), которые могут быть окислены пероксидом водорода в реакции хемилюминесценции, катализируемой пероксидазой.
Применение моноклональных антител Наиболее широко используются моноклональные антитела в медицинской диагностике. Разработаны сумки-укладки для постановки диагнозов. Если к антителами присоединить радиоактивные или магнитоактивные материалы и ввести их в живой организм, то можно выявить в нем патологические зоны. Такие МКА присоединяются к пораженным болезнью клеткам организма, а соответствующие индикаторные материалы позволяют выяснить их местонахождение. МКА используются и в процессах очистки веществ. Современные технологии основаны на присоединении антител к твердой матрице носителя. К ним добавляют смесь молекул, содержащую искомый антиген. Затем комплексы антиген — антитело отмываются от примесей, не связанных с матрицей. После разрушения ковалентных связей антиген — антитело в растворе остаются свободные антигены. Если получить антитела определенного типа и иммунизировать ими животное, то образуются анти-антитела (антиидиотипные антитела). Они действуют на иммунную систему как псевдоантиген и поэтому могут быть использованы для ее стимуляции. На этом принципе основано получение вакцин нового типа. Наборы МКА могут быть также предназначены для борьбы с аллергенами. Моноклональные антитела и "мишенная" лекарственная терапия. Предполагается, что большое разнообразие раковых заболеваний обусловлено активацией эндогенных генов, вызванной химическими агентами, внутренними хромосомными перестройками. Эти гены кодируют определенные белки, и поэтому раковые клетки могут содержать уникальные белки на поверхности клетки. Возможно, именно эти белки участвуют в супрессии роста здоровых клеток. Инактивируя эти белки, можно тормозить рост раковых клеток. Благодаря высокий специфичности МКА широко используются в качестве зондов для точного определения природы молекул поверхности клеток и клеточных органелл. С их помощью также можно проводить детекцию активности ферментов. Методы иммуноферментного анализа применяют в диагностике вирусных заболеваний растений. Это позволяет сократить время получения безвирусного посадочного материала, отбирать новые вирусоустойчивые сорта. При генноинженерных экспериментах можно быстро отбирать клоны — продуценты.
КЛОНИРОВАНИЕ ЖИВОТНЫХ
История метода Все клетки организма животных несут одинаковую генетическую информацию. Однако в процессе морфогенеза соматические клетки дифференцируются, в результате чего часть генома репрессируется. Чем выше уровень специализации клеток, тем меньше их тотипотентность. Эта закономерность была установлена в экспериментах по пересадке ядер. Впервые трансплантацию ядер соматических клеток зародышей в энуклеированные клетки лягушки осуществили американские исследователи Р. Бриггс и Т. Кинг в 1952 году. Ученые, пользуясь микропипеткой, удаляли ядра из яйцеклеток шпорцевой лягушки, а вместо них пересаживали ядра клеток эмбрионов, находящихся на разных стадиях развития. Проведенные исследования показали, что ядра ранних эмбрионов в стадии поздней бластулы и даже ранней гаструлы обладают тотипотентностью и обеспечивают нормальное развитие эмбрионов. Если брать ядра из клеток зародыша на ранней стадии его развития — бластуле, то примерно в 80 % случаев зародыш благополучно развивается дальше и превращается в нормального головастика. Если же развитие зародыша, донора ядра, продвинулось на следующую стадию — гаструлу, то лишь менее чем в 20 % случаев оперированные яйцеклетки развивались нормально. При пересадке ядер из более дифференцированных клеток (мезодермы и средней кишки) поздней гаструлы у эмбрионов наблюдалось недоразвитие и даже отсутствие нервной системы. После пересадки ядра из клеток более позднего развития яйцеклетки вообще не развивались. Более широкие исследования, охватывающие не только амфибий, но и рыб, а также дрозофил, в 1962 г. были начаты английским биологом Дж. Гордоном. Он первым в опытах с южноафриканскими жабами Xenopus laevis в качестве донора ядер использовал не зародышевые клетки, а уже вполне специализировавшиеся клетки эпителия кишечника плавающего головастика. Ядра яйцеклеток реципиентов он не удалял хирургическим путем, а разрушал ультрафиолетовыми лучами. В большинстве случаев реконструированные яйцеклетки не развивались, но примерно десятая часть их них образовывала эмбрионы. 6,5 % из этих эмбрионов достигали стадии бластулы, 2,5 % — стадии головастика и только 1 % развился в половозрелых особей. Однако появление нескольких взрослых особей в таких условиях могло быть связано с тем, что среди клеток эпителия кишечника развивающегося головастика довольно длительное время присутствуют первичные половые клетки, ядра которых могли быть использованы для пересадки. В последующих работах, как сам автор, так и многие другие исследователи не смогли подтвердить данные этих первых опытов. Позже Гордон модифицировал эксперимент. Поскольку большинство реконструированных яйцеклеток (с ядром клетки кишечного эпителия) погибают до завершения стадии гаструлы, он попробовал извлечь из них ядра на стадии бластулы и снова пересадить их в новые энуклеированные яйцеклетки (такая процедура называется "серийной пересадкой" в отличие от "первичной пересадки"). Число зародышей с нормальным развитием после этого увеличивалось, и они развивались до более поздних стадий по сравнению с зародышами, полученными в результате первичной пересадки ядер. Затем Гердон вместе с Ласки (1970) стали культивировать in vitro (вне организма в питательной среде) клетки почки, легкого и кожи взрослых животных и использовать уже эти клетки в качестве доноров ядер. Примерно 25 % первично реконструированных яйцеклеток развивались до стадии бластулы. При серийных пересадках они развивались до стадии плавающего головастика. Таким образом было показано, что клетки трех разных тканей взрослого позвоночного (X. laevis) содержат ядра, которые могут обеспечить развитие по крайней мере до стадии головастика. В свою очередь Ди Берардино и Хофнер (1983) использовали для трансплантации ядра неделящихся и полностью дифференцированных клеток крови — эритроцитов лягушки Rana pipiens. После серийной пересадки таких ядер 10 % реконструированных яйцеклеток достигали стадии плавающего головастика. Эти эксперименты показали, что некоторые ядра соматических клеток способны сохранять тотипотентность. Причины, по которым ядра клеток взрослых животных и даже поздних эмбрионов остаются тотипотентными, пока точно не установлены. Решающую роль играет взаимодействие ядра и цитоплазмы. Содержащиеся в цитоплазме животных вещества принимают участие в регулировании экспрессии клеточного генов ядра. Не исключено, что ядра дифференцированных клеток не могут изменить асинхронную репликацию ДНК на синхронную, характерную для ранних стадий эмбриогенеза. В связи с этим вопрос о возможности активации генов в дифференцированных соматических клетках представляет особую важность. Работы М. ди Бернардино и Н. Хоффера показали, что цитоплазма ооцитов амфибий содержит факторы, восстанавливающие тотипотентность ядер дифференцированных соматических клеток. Эти факторы реактивируют репрессированные участки генома. В 1985 г. была описана технология клонирования костных рыб, разработанная советскими учеными Л.А. Слепцовой, Н.В. Дабагян и К.Г.Газарян. Зародыши на стадии бластулы отделяли от желтка. Ядра клеток зародышей впрыскивали в цитоплазму неоплодотворенных икринок, которые начинали дробиться и развивались в личинки. Эти эксперименты показали, что потеря ядром тотипотентности в процессе онтогенеза связана не с утерей генов, а их репрессией. При культивировании соматических клеток in vitro частота тотипотентности ядер увеличивается. Генетический механизм стабильной репрессии генома дифференцированных клеток не выяснен, способы восстановления тотипотентности не разработаны, поэтому в основном ведется клонирование путем трансплантации ядер эмбриональных клеток. Пересадки ядер у млекопитающих начались позднее, в 80-х годах. Это было связано с техническими трудностями, так как зигота млекопитающих имеет небольшие размеры. Например, диаметр зиготы мыши приблизительно 60 мкм, а диаметр оплодотворенной яйцеклетки лягушки около 1200 мкм, т. е. в 20 раз больше. Зигота коровы несколько крупнее, чем зигота мыши, диаметр ее составляет 160 мкм, но пронуклеусы скрыты яичным желтком, поэтому перед микроманипуляциями необходима специальная обработка зигот. Несмотря на перечисленные трудности, первые сообщения о получении клонов мышей, идентичных донору, появились уже в 1981 году. В качестве донора были использованы эмбриональные клетки одной из линий мышей, взятые на стадии бластоцисты. Достоверность полученных данных вначале была поставлены под сомнение, так как воспроизвести результаты проведенных экспериментов в других лабораториях не удавалось, однако пару лет спустя Дж. Мак Грат и Д. Солтер также достигли успеха. В этих экспериментах клоны мышей удавалось получить лишь в том случае, если трансплантировали ядра эмбрионов на стадии не позднее 2 бластомеров. Было показано, что ядра 8-клеточных зародышей и клеток внутренней клеточной массы бластоцисты не обеспечивают развитие in vitro реконструированных яйцеклеток даже до стадии морулы, которая предшествует стадии бластоцисты. Небольшая часть (5 %) ядер 4-клеточных зародышей дает возможность развиваться только до стадии морулы. Эти и многие другие данные показывают, что в эмбриогенезе у мышей клеточные ядра рано теряют тотипотентность, что связано очевидно, с очень ранней активацией генома зародыша — уже на стадии 2-х клеток. У других млекопитающих, в частности, у кроликов, овец и крупного рогатого скота, активация первой группы генов в эмбриогенезе происходит позднее, на 8-16-клеточной стадии. Возможно, поэтому первые значительные успехи в клонировании эмбрионов были достигнуты на других видах млекопитающих, а не на мышах. Тем не менее, работы с мышами, несмотря на их непростую судьбу, значительно расширили наши представления о методологии клонирования млекопитающих[74].
Методы трансплантации ядер В нашей стране Б.В. Конюховым и Е.С. Платоновым в 1985 г. был разработан метод менее травматического переноса ядер методом микроманипуляции. Он протекает в два этапа: сначала тонкой микропипеткой прокалывают зоны пеллюцида и плазматической мембраны и извлекают пронуклеусы, а затем другой пипеткой, большего диаметра (12 мкм) в то же отверстие вводят диплоидное ядро донора. В этом случае меньше травмируется цитоплазма зиготы и транспортируемое ядро донора. Трансплантация ядер может осуществляться и другим способом, с использованием цитохалазинов (веществ, синтезируемых грибами). Цитохалазин В разрушает структуру микрофиламентов и способствует уникальному расположению ядра. Ядро остается соединенным с клеткой тоненьким стебельком цитоплазмы. При центрифугировании этот мостик разрывается, образуются безъядерные клетки (цитопласты) и кариопласты, представляющие собой ядра, окруженные тонким слоем цитоплазмы и цитоплазматической мембраной. Цитопласты отделяют от интактных клеток в градиенте плотности. Они сохраняют способность прикрепляться к поверхности культурального сосуда и могут быть использованы для слияния с кариопластами других клеток с целью получения жизнеспособной клетки. Методы выделения кариопластов несколько сложнее и включают в себя ряд операции по центрифугированию, разделению в градиенте плотности и т. д. В некоторых случаях к смеси клеток и кариопластов добавляют частицы тантала диаметром 1–3 мкм. Они проникают в клетки и никогда в кариопласт, поэтому более тяжелые клетки осаждаются быстрее кариопластов. Цитопласты содержат все виды органелл, присущие нормальной клетке, сохраняют способность прикрепляться к субстрату, образовывать складчатую мембрану, передвигаться, осуществлять пиноцитоз. Кариопласты окружены тонким слоем цитоплазмы (около 10 % от всей клеточной цитоплазмы), содержат компактный эндоплазматический ретикулум, несколько митохондрий и рибосом. У некоторых клеточных линий 1/10 кариопластов способна восстановить весь утраченный объем цитоплазмы и восстановиться в жизнеспособные клетки. Для реконструкции клеток суспензию кариопластов в солевом буфере добавляют к монослою культуры цитопластов из пропорции 100 кариопластов на 1 цитопласт. Цитопласты должны быть уже покрыты инактивированными вирусными частицами. Инкубируют при температуре 4 °C 45 минут, а затем еще 45 минут при температуре 37 °C. Отмывают раствором Эрла для удаления не слившихся кариопластов.
Клонирование млекопитающих Американские исследователи С. Стик и Дж. Робл, используя методику МакГрата и Солтера, в 1988 г. получили 6 живых кроликов, пересадив ядра 8 клеточных эмбрионов одной породы в лишенные ядра яйцеклетки кроликов другой породы. Фенотип родившихся полностью соответствовал фенотипу донора. В этих экспериментах только 6 из 164 реконструированных яйцеклеток (3,7 %) развились в нормальных животных. Это, конечно, очень низкий выход, практически не позволяющий рассчитывать на получение таким методом клона генетически идентичных животных. Ценность этой работы тем не менее в том, что она показала возможность клонирования эмбрионов кроликов. Первые успешные эксперименты по клонированию сельскохозяйственных животных были проведены С. Уилладсином (S.Willadsen) в 1986 г. Он сливал безъядерные яйцеклетки с бластомерами, выделенными из 8 и 16-клеточного эмбриона овцы. Дж. Робл и его сотрудники в 1987 провели работы по пересадке ядер крупного рогатого скота. Они пересаживали в зиготы кариопласты — мужской и женский пронуклеусы вместе с окружающей их цитоплазмой, а также ядра 2-, 4- или 8-клеточных эмбрионов коровы. Сначала зиготы центрифугировали чтобы освободить пронуклеусы от окружающих их гранул желтка, после чего ядра были хорошо видны под микроскопом, что значительно облегчало их удаление. При помощи манипулятора и заостренной стеклянной микропипетки извлекали один из бластомеров вместе с ядром из ранних зародышей и переносили его в энуклеированную зиготу. Реконструированные зародыши были заключены в агаровый цилиндр и пересажены в перевязанный яйцевод овцы. Через пять дней культивирования их вымывали, освобождали от агара и исследовали. Реконструированные зародыши в этой работе развивались только в тех случаях, когда в зиготы пересаживали пронуклеусы: 17 % таких зародышей достигли стадии морулы или бластоцисты. Два зародыша были пересажены второму реципиенту — в матку коровы, и развитие их завершилось рождением живых телят. Если в качестве доноров использовали ядра 2-, 4- или 8-клеточных зародышей, то реконструированные яйцеклетки не развивались даже до стадии морулы. Позже были и более успешные работы. С. Уиладсин (1989), в частности, сообщил, что ему удалось получить четырех генетически идентичных бычков холстейнской породы в результате пересадки в реципиентные яйцеклетки ядер бластомеров одного 32-клеточного зародыша. Автор утверждал, что большинство ядер сохраняет тотипотентность на 32-клеточной стадии, а значительная их часть даже на 64-клеточной стадии, обеспечивая нормальное развитие реконструированных яйцеклеток до стадии ранней бластоцисты в яйцеводе овцы. После пересадки в матку коров — окончательных реципиентов, как полагает автор, они могут и дальше нормально развиваться. К. Бондиоли и соавторы (1990), используя в качестве доноров ядер 16-64-клеточные зародыши коров, трансплантировали 463 реконструированных зародыша в матку синхронизированных реципиентов, и было получено 92 живых теленка. Семь из них были генетически идентичны, представляя собой клон, полученный в результате пересадки ядер клеток одного донорского эмбриона. Таким образом, клеточные ядра зародышей крупного рогатого скота достаточно долго сохраняют тотипотентность и могут обеспечить полное развитие реконструированных яйцеклеток. Иначе говоря, методические трудности клонирования зародышей крупного рогатого скота практически решены. Экспериментов по клонированию свиней немного. Успешные исследования провели Р. Пратер с сотрудниками в 1989 г. Скудность данных, видимо, связана с определенными трудностями работы с этим объектом. В 1993–1995 годах, группа исследователей под руководством Я. Уилмута (Ian Wilmut) из Рослинского института получила клон овец — 5 идентичных животных, донорами ядер которых была культура эмбриональных клеток. Клеточную культуру получали следующим образом: выделяли микрохирургически эмбриональный диск из 9-дневного овечьего эмбриона (бластоцисты) и культивировали клетки in vitro в течение многих пассажей (по крайней мере до 25). Сначала клеточная культура напоминала культуру стволовых недифференцированных эмбриональных клеток, но вскоре, после 2-3-х пассажей, клетки становились уплотненными и морфологически сходными с эпителиальными. Эта линия клеток из 9-дневного зародыша овцы была обозначена как TNT4. Чтобы донорское ядро и реципиентная цитоплазма находились на сходных стадиях клеточного цикла, останавливали деление культивируемых клеток TNT4 на определенной стадии (GO) и ядра этих клеток пересаживали в энуклеированные яйцеклетки (соответственно на стадии метафазы II). Реконструированные эмбрионы заключали в агар и трансплантировали в перевязанные яйцеводы овец. Через 6 дней эмбрионы вымывали из яйцевода первого реципиента и исследовали под микроскопом. Отбирали те, которые достигли стадии морулы или бластоцисты и пересаживали их в матку овцы — окончательного реципиента, где развитие продолжалось до рождения. Родилось 5 ягнят (самок) из них 2 погибли вскоре после рождения, 3-й в возрасте 10 дней, а 2 оставшихся нормально развивались и достигли 8-9-месячного возраста. Фенотипически все ягнята были сходны с породой овец, от которой получали исходную линию клеток TNT4. Это подтвердил и генетический анализ. Эта работа, особенно в части культуры эмбриональных клеток, — значительное достижение в клонировании млекопитающих, хотя она и не вызвала столь шумного интереса, как статья того же Уилмута с соавторами, опубликованная в начале 1997 года, где сообщалось, что в результате использования донорского ядра клетки молочной железы овцы было получено клональное животное — овца по кличке Долли. Последняя работа методически во многом повторяет предыдущее исследование 1996 года, но в ней ученые использовали эмбриональные и фибробластоподобные клетки плода и клетки молочной железы взрослой овцы. Клетки молочной железы получали от шестилетней овцы породы финн дорсет, находящейся на последнем триместре беременности. Все три типа клеточных культур имели одинаковое число хромосом — 54, как обычно у овец. Эмбриональные клетки использовали в качестве доноров ядер на 7-9-м пассажах культивирования, фибробластоподобные клетки плода — на 4-6-м пассажах и клетки молочной железы — на 3-6-м пассажах. Деление клеток всех трех типов останавливали на стадии GO и ядра клеток пересаживали в энуклеированные ооциты (яйцеклетки) на стадии метафазы II. Был использован метод электрослияния. Большинство реконструированных эмбрионов сначала культивировали в перевязанном яйцеводе овцы, но некоторые и in vitro в химически определенной среде. Коэффициент выхода морул или бластоцист при культивировании in vitro в одной серии опытов был даже вдвое выше, чем при культивировании в яйцеводе. Выход морул или бластоцист в серии опытов с культурой клеток молочной железы был примерно втрое меньше, чем в двух других сериях, когда в качестве доноров ядер использовали культуру фибробластов плода или эмбриональных клеток. Число живых ягнят в сравнении с числом пересаженных в матку окончательного реципиента морул или бластоцист было также в два раза ниже. В серии опытов с клетками молочной железы из 277 реконструированных яйцеклеток был получен только один живой ягненок, что говорит об очень низкой результативности такого рода экспериментов (0,36 %). Анализ генетических маркеров всех семи родившихся в трех сериях экспериментов живых детенышей показал, что клетки молочной железы были донорами ядер для одного, фибробласты плода — для двух и эмбриональные клетки — четырех ягнят. Овца по кличке Долли развилась из реконструированной яйцеклетки, донором ядра которой была культивируемая клетка молочной железы овцы породы финн дорсет и фенотипически не отличается от овец этой породы, но сильно отличается от овцы-реципиента. Анализ генетических маркеров подтвердил этот результат. Успех авторов этой работы, прежде всего, связан с использованием длительных клеточных культур, так как после многих пассажей в культуре клеток могли быть отобраны малодифференцированные стволовые клетки, которые, вероятно, и были использованы как доноры ядер. Большое значение также имел тот факт, что авторы, учитывая результаты своих предыдущих работ, синхронизировали стадии клеточного цикла яйцеклеток реципиентов и клеток доноров. Аналогичные эксперименты проводили позднее Tanja Dominko и сотрудники лаборатории Висконсинского университета, которые обеспечили клонирование эмбрионов из клеток кожи ушей взрослого рогатого скота. Эмбрионы, генетически идентичные корове, пожертвовавшей клетки уха, были внедрены в матки коров — рецепиентов. Наблюдалась постепенная гибель эмбрионов, поэтому жизнеспособных телят не получили. Причины пока не установлены. В августе 1997 года появилось сообщение о том, что Алан Троунсон (Австралия) разработал технологию, которая позволяет сформировать эмбрион из 16, 32 или 64 клеток и затем каждая из них может использоваться для формирования 16, 32 или 64 идентичных эмбрионов. Коллектив исследователей во главе с Аланом Троунсоном создал 470 генетически идентичных эмбрионов рогатого скота от единственной бластоцисты. Такая технология обеспечивает безграничный источник генетического материала для клонирования. Несмотря на отсутствие немедленных практических результатов сделанного открытия, теоретическую значимость его трудно переоценить. Впервые было доказано, что гены запрограммированы обратимо. Дальнейшие исследования могут позволить понять, как регулируется работа генов, дифференциация клеток, почему клетки в одних случаях растут и размножаются управляемо, а в других (при раке) — неконтролируемо. Уже сейчас корпорация Genzyme Transgenics планирует исследования с целью создания трансгенного крупного рогатого скота, содержащего в молоке человеческий альбумин. Был куплен патент на получение эмбрионов, содержащих геном клеток соединительной ткани (фибробластов), включающий ген, ответственный за синтез человеческого белка. Несколько коров в настоящее время беременны трансгенными телятами. Подобная технология позволяет увеличить эффективность создания трансгенных молочных животных, так как при обычном впрыскивании генов в оплодотворенную яйцеклетку рождается только 5-10 % трансформированных животных, из них — несколько самцов, не дающих молока. Использование новой технологии клонирования позволяет получать животных только женского пола, дающих трансгенный протеин.
РЕГУЛИРОВАНИЕ ВОСПРОИЗВОДСТВА СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
Воспроизводство животных — это основной фактор, лимитирующий эффективность производства животноводческих продуктов на промышленной основе. Причины, препятствующие достижению оптимальных результатов в воспроизводстве домашнего скота различны. Новые методы расширяют возможности регулирования воспроизводства. Они связаны с манипулированием на уровне клеток или эмбрионов, с использованием физиологически активных соединений, поэтому названы биотехнологическими. К числу этих методов относят: стимуляцию и синхронизацию окоты, суперовуляцию, искусственное осеменение, трансплантацию эмбрионов, хранение гамет и эмбрионов, целенаправленное получение двоен, регулирование пола, раннюю диагностику беременности, управление процессом родов, создание химер и Стимуляция и синхронизация окоты осуществляется с помощью прогестерона — женского полового гормона стероидной природы, регулирующего ход эстрального цикла, простагландинов, а также их комбинации. Этот прием позволяет вызывать появление окоты у групп племенных животных в один и тот же период времени. В США для синхронизации окоты у телок в молочном и мясном скотоводстве выпускается новый препарат под названием "Синхромейт-В". Он представляет совокупность двух гормонов, один из которых имплантируется под кожу, а другой инъецируется внутримышечно. Имплант помещается под кожу уха телки и сразу же после этого следует инъекция другого гормона. Под действием этих двух гормонов эстральный цикл телки прерывается и временно останавливается. Через некоторое время имплант удаляют из уха животного, начинается новый эстральный цикл. Так как имплант удаляется одновременно у всех телок, то и цикл начинается в одно и то же время. Применение гормональных препаратов снимает необходимость ежедневного контроля за состоянием половой активности животных. Преимущество синхронизированной охоты состоит в реальной возможности формирования однородных групп животных в период осеменения, одновременности рождения приплода, точном учете кормов в группах.
Суперовуляция Потенциальные возможности воспроизводства самок млекопитающих огромны. В их яичниках содержатся десятки и сотни тысяч овоцитов. Однако в процессе онтогенеза лишь небольшая часть из них реализуется в виде потомков. Остальные овоциты подвергаются атрезии (обратному развитию) и воспроизводстве не участвуют. Суперовуляция — состояние, вызванное гормонами, когда в яичниках животных развивается и овулирует в несколько раз больше яйцеклеток. В зависимости от вида число овулирующих яйцеклеток может быть увеличено в 3–8 и даже в 50 раз. С помощью этого приема становится возможным получение большего количества эмбрионов от лучших по продуктивности коров[75].
Искусственное осеменение Искусственное осеменение животных является самым старым и хорошо отработанным биотехнологическим методом разведения сельскохозяйственных животных. Применение этого метода позволяет ограничить распространение половых инфекций, которые нередко служат причиной бесплодия животных. Оно также позволяет эффективно использовать генетический потенциал лучших производителей. Экономический эффект от искусственного осеменения обусловлен снижением затрат на содержание большого поголовья производителей, возможностью быстрого размножения генотипа с хозяйственно — полезными признаками, улучшением генетического потенциала ремонтного стада.
Трансплантация эмбрионов Трансплантация эмбрионов в настоящее время является одной из наиболее актуальных проблем в области животноводства. С помощью пересадки эмбрионов можно резко увеличить выход числа потомков от высокопродуктивных коров. Трансплантация эмбрионов, или эмбриотехнология, заключается в получении одного или нескольких эмбрионов из матки племенных животных (доноров) и пересадке в матку коров (рецепиентов), где эмбрионы развиваются до отела. Этот методв сочетании с суперовуляцией у доноров позволяет получить большое потомство от высокопродуктивных животных. Этим способом эмбрионы можно внедрить в ту или иную породу в другие регионы, используя в качестве рецепиентов коров мясных пород. Применение этого метода также упрощает обмен генофондом сельскохозяйственных животных между странами и континентами. Пересадка эмбрионов может быть использована для получения потомства от ценных, но бесплодных коров, утративших способность к размножению в результате несчастного случая, болезни или по возрасту. Когда было установлено, что кролик обладает иммунитетом по отношению к ящуру, была выдвинута идея использования метода трансплантации для оздоровления потомства зараженных ящуром животных. Половые пути кролика, куда трансплантируются эмбрионы, способны разрушать вирус ящура в эмбрионах. Трансплантация может быть использована и для временного хранения эмбрионов. В яйцеводах крольчих удается осуществлять трансконтинентальную перевозку эмбрионов овец. Извлечение эмбрионов до 70-х годов производили в основном хирургическим путем, впоследствии он был заменен менее травматичным и трудоемким нехирургическим, основанным на введении в матку особого зонда по естественному каналу. Зонд имеет три канала. Один из каналов предназначен для надувания баллончика, который закупоривает рог матки, препятствуя вытеканию жидкости. По другому каналу вводится физиологический раствор с температурой 25–30 °C, который вымывает эмбрионы и возвращается вместе с ними через третий канал зонда в пробирку, помещенную в водяную баню с температурой 35 °C. Из этой жидкости извлекаются эмбрионы. В среднем при суперовуляции от донора можно получить от 5 до 7 эмбрионов. Трансплантацию производят с помощью специального зонда или пистолета для осеменения. Эмбрионы помещаются в рога матки. Стельность у самок — рецепиентов проверяется по уровню прогестерона в плазме крови на 21-й день.
Регулирование пола В практике разведения животных очень важно научиться управлять образованием в потомстве мужских и женских особей. Метод разделения эмбрионов по полу основан на определении белков, специфичных для самцов. Этом метод широко применяется в животноводческой практике многих стран. В Канаде уже с 1975 года рождаются телята, разделенные по полу на стадии эмбрионов. В перспективе для целенаправленного получения особей мужского или женского пола может быть применен метод микрохирургической замены X и У хромосом. Такие манипуляции уже проводились на растительных клетках и яйцеклетках земноводных.
ЛИТЕРАТУРА К РАЗДЕЛУ «КУЛЬТУРЫ ЖИВОТНЫХ КЛЕТОК»
1. Алберте Б., Брей Д., Льюис Дж. и др. Молекулярная биология клетки. Т. 1. М.: Мир, 1994. 2. Артамонов В. И. Биотехнология — агропромышленному комплексу. М.: Наука, 1989. 160 с. 3. Артамонов В.И. Сельские профессии биотехнологии. М.: Изд-во МСХА, 1992. 127 с. 4. Бекер М.Е., Лиепиньш Г.К., Райпулис Е.П. Биотехнология. М.: Агропромиздат, 1990. 334 с. 5. Березин И.В., Клесов А. А., Швядас В.К., Угарова Н.Н., Варфоломеев С.Д., Ярополов А.И., Казанская Н.Ф., Егоров А.М. Инженерная энзимология. // Биотехнология. Кн. 8. М.: Высшая школа, 1987. 143 с. 6. Герасименко В.Г. Биотехнология. Киев: Выща школа, 1989. 343 с. 7. Гринберг К.Н., Кухаренко В.И., Ляшко В.Н., Терехов С.М., Пичугина Е. М., Фрейдин М.И., Чериков В.Г. Культивирование фибробластов человека для диагностики наследственных болезней. // Методы культивирования клеток. Л.: Наука, 1988. С. 251–260. 8. 3авертляев Б.П. Биотехнология в воспроизводстве и селекции крупного рогатого скота. Л.: Агропромиздат, 1989. 255 с. 9. Какпаков В. Т. Культивирование клеток и тканей беспозвоночных. // Методы культивирования клеток. Л.: Наука, 1988. С. 241–250. 10. Конюхов Б. В. Долли — случайность или закономерность? // Человек. 1998. № 3 11. Крыжанов М.А., Залесских А.Ф., Троицкая М.В., Соловьев В.В., Тихомиров А.И. Культивирование клеточной линии НЕР-2 на микроносителях с целью получения вируса полиомиелита. // Биотехнология и генетика. Н.Новгород, 1991. С. 87–92. 12. Прудовский И.А., Сухарев С.И. Гибридизация клеток животных. // Методы культивирования клеток. Л.: Наука, 1988. С. 176–193 13. Рингертц Н., Сэвидж Р. Гибридные клетки. М.: Мир, 1979. 412 с. 14. Спиер Р.Е., Адамс Г.Д., Гриффитс Дж. Б. и др. Биотехнология клеток животных. М.: Агропромиздат, 1989. Т. 1, 2. 15. Тартаковский А.Д. Питательные среды для культивирования клеток млекопитающих. // Методы культивирования клеток. Л.: Наука, 1988. С. 44–63. 16. Фрешни Р. Культура животных клеток. Методы. М.: Мир, 1989. 318 с. 17. Фридлянская И.И. Получение моноклональных антител (гибридомная технология). // Методы культивирования клеток. Л.: Наука, 1988. С. 194–205. 18. Цуцаева А.А., Петренко Т.Ф. Криоконсервация культивируемых клеток животных. // Методы культивирования клеток. Л.: Наука, 1988. С. 63–69. 19. Wilmut I. et al. Viable offspring derived from fetal and adult mammalian cell line // Nature. 1997. V.385. P. 810–813.
Генная инженерия
ВВЕДЕНИЕ В ГЕНЕТИЧЕСКУЮ ИНЖЕНЕРИЮ
Возможности генной инженерии Важной составной частью биотехнологии является генетическая инженерия. Родившись в начале 70-х годов, она добилась сегодня больших успехов. Методы генной инженерии преобразуют клетки бактерий, дрожжей и млекопитающих в "фабрики" для масштабного производства любого белка. Это дает возможность детально анализировать структуру и функции белков и использовать их в качестве лекарственных средств. В настоящее время кишечная палочка (Е. coli) стала поставщиком таких важных гормонов как инсулин и соматотропин. Ранее инсулин получали из клеток поджелудочной железы животных, поэтому стоимость его была очень высока. Для получения 100 г кристаллического инсулина требуется 800-1000 кг поджелудочной железы, а одна железа коровы весит 200–250 грамм. Это делало инсулин дорогим и труднодоступным для широкого круга диабетиков. В 1978 году исследователи из компании "Генентек" впервые получили инсулин в специально сконструированном штамме кишечной палочки. Инсулин состоит из двух полипептидных цепей А и В длиной 20 и 30 аминокислот. При соединении их дисульфидными связями образуется нативный двухцепочечный инсулин. Было показано, что он не содержит белков Е. coli, эндотоксинов и других примесей, не дает побочных эффектов, как инсулин животных, а по биологической активности от него не отличается. Впоследствии в клетках Е. coli был осуществлен синтез проинсулина, для чего на матрице РНК с помощью обратной транскриптазы синтезировали ее ДНК-копию. После очистки полученного проинсулина его расщепили и получили нативный инсулин, при этом этапы экстракции и выделения гормона были сведены к минимуму. Из 1000 литров культуральной жидкости можно получать до 200 граммов гормона, что эквивалентно количеству инсулина, выделяемого из 1600 кг поджелудочной железы свиньи или коровы. Соматотропин — гормон роста человека, секретируемый гипофизом. Недостаток этого гормона приводит к гипофизарной карликовости. Если вводить соматотропин в дозах 10 мг на кг веса три раза в неделю, то за год ребенок, страдающий от его недостатка, может подрасти на 6 см. Ранее его получали из трупного материала, из одного трупа: 4–6 мг соматотропина в пересчете на конечный фармацевтический препарат. Таким образом, доступные количества гормона были ограничены, кроме того, гормон, получаемый этим способом, был неоднороден и мог содержать медленно развивающиеся вирусы. Компания "Genentec" в 1980 году разработала технологию производства соматотропина с помощью бактерий, который был лишен перечисленных недостатков. В 1982 году гормон роста человека был получен в культуре Е. coli и животных клеток в институте Пастера во Франции, а с 1984 года начато промышленное производство инсулина и в СССР. При производстве интерферона используют как Е. coli, S. cerevisae (дрожжи), так и культуру фибробластов или трансформированных лейкоцитов. Аналогичными методами получают также безопасные и дешевые вакцины. На технологии рекомбинантных ДНК основано получение высокоспецифичных ДНК-зондов, с помощью которых изучают экспрессию генов в тканях, локализацию генов в хромосомах, выявляют гены, обладающие родственными функциями (например, у человека и курицы). ДНК-зонды также используются в диагностике различных заболеваний. Технология рекомбинантных ДНК сделала возможным нетрадиционный подход "белок-ген", получивший название "обратная генетика". При таком подходе из клетки выделяют белок, клонируют ген этого белка, модифицируют его, создавая мутантный ген, кодирующий измененную форму белка. Полученный ген вводят в клетку. Если он экспрессируется, несущая его клетка и ее потомки будут синтезировать измененный белок. Таким образом, можно исправлять дефектные гены и лечить наследственные заболевания. Если гибридную ДНК ввести в оплодотворенное яйцеклетку, могут быть получены трансгенные организмы, экспрессирующие мутантный ген и передающие его потомками. Генетическая трансформация животных позволяет установить роль отдельных генов и их белковых продуктов, как в регуляции активности других генов, так и при различных патологических процессах. С помощью генетической инженерии созданы линии животных, устойчивых к вирусным заболеваниям, а также породы животных с полезными для человека признаками. Например, микроинъекция рекомбинантной ДНК, содержавшей ген соматотропина быка в зиготу кролика позволила получить трансгенное животное с гиперпродукцией этого гормона. Полученные животные обладали ярко выраженной акромегалией. Сейчас даже трудно предсказать все возможности, которые будут реализованы в ближайшие несколько десятков лет.
Генная инженерия как наука, методы Генетическая инженерия — конструирование in vitro функционально активных генетических структур (рекомбинантных ДНК), или иначе — создание искусственных генетических программ (Баев А. А.). По Э. С. Пирузян генетическая инженерия — система экспериментальных приемов, позволяющих конструировать лабораторным путем (в пробирке) искусственные генетические структуры в виде так называемых рекомбинантных или гибридных молекул ДНК. Речь идет о направленном, по заранее заданной программе конструировании молекулярных генетических систем вне организма с последующим введением их в живой организм. При этом рекомбинантные ДНК становятся составной частью генетического аппарата рецепиентного организма и сообщают ему новые уникальные генетические, биохимические, а затем и физиологические свойства. Цель прикладной генетической инженерии заключается в конструировании таких рекомбинантных молекул ДНК, которые при внедрении в генетический аппарат придавали бы организму свойства, полезные для человека. Технология рекомбинантных ДНК использует следующие методы: — специфическое расщепление ДНК рестрицирующими нуклеазами, ускоряющее выделение и манипуляции с отдельными генами; — быстрое секвенирование всех нуклеотидов очищенном фрагменте ДНК, что позволяет определить границы гена и аминокислотную последовательность, кодируемую им; — конструирование рекомбинантной ДНК; — гибридизация нуклеиновых кислот, позволяющая выявлять специфические последовательности РНК или ДНК с большей точностью и чувствительностью, основанную на их способности связывать комплементарные последовательности нуклеиновых кислот; — клонирование ДНК: амплификация in vitro с помощью цепной полимеразной реакции или введение фрагмента ДНК в бактериальную клетку, которая после такой трансформации воспроизводит этот фрагмент в миллионах копий; — введение рекомбинантной ДНК в клетки или организмы.
История генетической инженерии Генная инженерия появилась благодаря работам многих исследователей в разных отраслях биохимии и молекулярной генетики. На протяжении многих лет главным классом макромолекул считали белки. Существовало даже предположение, что гены имеют белковую природу. Лишь в 1944 году Эйвери, Мак Леод и Мак Карти показали, что носителем наследственной информации является ДНК. С этого времени начинается интенсивное изучение нуклеиновых кислот. Спустя десятилетие, в 1953 году Дж. Уотсон и Ф. Крик создали двуспиральную модель ДНК. Именно этот год принято считать годом рождения молекулярной биологии. На рубеже 50—60-х годов были выяснены свойства генетического кода, а к концу 60-х годов его универсальность была подтверждена экспериментально. Шло интенсивное развитие молекулярной генетики, объектами которой стали Е. coli, ее вирусы и плазмиды. Были разработаны методы выделения высокоочищенных препаратов неповрежденных молекул ДНК, плазмид и вирусов. ДНК вирусов и плазмид вводили в клетки в биологически активной форме, обеспечивая ее репликацию и экспрессию соответствующих генов. В 70-х годах был открыт ряд ферментов, катализирующих реакции превращения ДНК. Особая роль в развитии методов генной инженерии принадлежит рестриктазам и ДНК-лигазам. Историю развития генетической инженерии можно условно разделить на три этапа. Первый этап связан с доказательством принципиальной возможности получения рекомбинантных молекул ДНК in vitro. Эти работы касаются получения гибридов между различными плазмидами. Была доказана возможность создания рекомбинантных молекул с использованием исходных молекул ДНК из различных видов и штаммов бактерий, их жизнеспособность, стабильность и функционирование. Второй этап связан с началом работ по получению рекомбинантных молекул ДНК между хромосомными генами прокариот и различными плазмидами, доказательством их стабильности и жизнеспособности. Третий этап — начало работ по включению в векторные молекулы ДНК (ДНК, используемые для переноса генов и способные встраиваться в генетический аппарат клетки-рецепиента) генов эукариот, главным образом, животных. Формально датой рождения генетической инженерии следует считать 1972 год, когда в Стенфордском университете П. Берг, С. Коэн, X. Бойер с сотрудниками создали первую рекомбинантную ДНК, содержавшую фрагменты ДНК вируса SV40, бактериофага и Е. coli.
ФЕРМЕНТЫ ГЕНЕТИЧЕСКОЙ ИНЖЕНЕРИИ
Основные группы ферментов Генетическая инженерия — потомок молекулярной генетики, но своим рождением обязана успехам генетической энзимологии и химии нуклеиновых кислот, так как инструментами молекулярного манипулирования являются ферменты. Если с клетками и клеточными органеллами мы подчас можем работать микроманипуляторами, то никакие, даже самые мелкие микрохирургические инструменты не помогут при работе с макромолекулами ДНК и РНК. Что же делать? В роли "скальпеля", "ножниц" и "ниток для сшивания" выступают ферменты. Только они могут найти определенные последовательности нуклеотидов, "разрезать" там молекулу или, наоборот, "заштопать" дырку в цепи ДНК. Эти ферменты издавна работают в клетке, выполняя работы по репликации (удвоению) ДНК при делении клетки, репарации повреждений (восстановлению целостности молекулы), в процессах считывания и переноса генетической информации из клетки в клетку или в пределах клетки. Задача генного инженера — подобрать фермент, который выполнил бы поставленные задачи, то есть смог бы работать с определенным участком нуклеиновой кислоты. Следует отметить, что ферменты, применяемые в генной инженерии, лишены видовой специфичности, поэтому экспериментатор может сочетать в единое целое фрагменты ДНК любого происхождения в избранной им последовательности. Это позволяет генной инженерии преодолевать установленные природой видовые барьеры и осуществлять межвидовое скрещивание. Ферменты, применяемые при конструировании рекомбинантных ДНК, можно разделить на несколько групп: — ферменты, с помощью которых получают фрагменты ДНК (рестриктазы); — ферменты, синтезирующие ДНК на матрице ДНК (полимеразы) или РНК (обратные транскриптазы); — ферменты, соединяющие фрагменты ДНК (лигазы); — ферменты, позволяющие осуществить изменение структуры концов фрагментов ДНК.
Рестриктазы Рестриктазы (рестрицирующие эндонуклеазы, эндонуклеазы рестрикции) — это ферменты, узнающие и атакующие определенные последовательности нуклеотидов в молекуле ДНК (сайты рестрикции). Еще в 1953 году было обнаружено, что ДНК определенного штамма Е. coli, введенная в клетки другого штамма (например, ДНК штамма В — в клетки штамма С) не проявляет, как правило, генетической активности, так как быстро расщепляется на мелкие фрагменты. В 1966 году было показано, что это явление связано со специфической модификацией хозяйской ДНК — она содержит несколько метилированных оснований, отсутствующих в немодифицированной ДНК, причем метилирование (добавление к основанию метильной группы) происходит уже после завершения репликации. Бактерия способна отличить свою собственную ДНК от любой вторгающейся «чужеродной» именно по типу ее модификации. За «метку» отвечают метилирующие ферменты модификации, так называемые ДНК-метилазы. Различие в модификации делает чужеродную ДНК чувствительной к действию рестрицирующих ферментов, которые узнают отсутствие метильных групп в соответствующих сайтах. Системы рестрикции и модификации широко распространены у бактерий; их существование играет важную роль в защите резидентной ДНК от загрязнения последовательностями чужеродного происхождения. Рестриктаза, которая расщепляла неметилированную ДНК была выделена в 1968 г. Мезельсоном и Юанем. Этот фермент был высокоспецифичен по отношению к определенной последовательности ДНК, но расщеплял молекулы неспецифически, в другом месте, на некотором удалении от участка узнавания. Вскоре, в 1970 г. Смит и Вилькокс выделили из Haemophilus influenzae первую рестриктазу, которая расщепляла строго определенную последовательность ДНК (Hind III). Поскольку разные бактерии по-разному метят свою ДНК, то и рестриктазы должны узнавать разные последовательности. И действительно, с тех пор выделены рестриктазы, узнающие более 150 сайтов рестрикции (мест расщепления ДНК).
Полимеразы Впервые ДНК-полимераза была выделена Корнбергом с сотрудниками в 1958 году из Е. coli. ДНК-полимераза I Е. coli (Pol I) не связывается с молекулами двухцепочечной кольцевой ДНК. Однако если такие молекулы денатурировать и получить одноцепочечные формы, то с последними полимераза связывается в количествах, пропорциональных длине этих участков — примерно одна молекула на 300 нуклеотидных остатков. Pol 1 связывается с одноцепочечными участками двойной спирали ДНК, в местах одноцепочечных разрывов с 3'-гидроксилом и 5'-фосфатом, а также с концами двухцепочечных молекул ДНК. Фермент состоит из мономерной полипептидной цепи с молекулярной массой 103 кДа и имеет 3-х доменную структуру. Каждый домен обладает своей ферментативной активностью: 5'-3' полимеразной, 3'-5' экзонуклезной, 5'-3' экзонуклеазной. 1. 5'-3' полимеразная активность. Для реакции необходимо наличие одноцепочечной ДНК-матрицы и комплементарного участку этой цепи фрагмента — праймера (затравки) с 3'-ОН концом. 2. 3'-5' экзонуклеазная активность. Гидролизует одноцепочечную или двухцепочечную ДНК с 3'-ОН конца. 3'—5' нуклеаза расщепляет диэфирную связь только в неспаренных участках ДНК. Известно, что при полимеразной реакции с определенной частотой возможно включение в растущую цепь некомплементарного нуклеотида. Однако полимераза не может присоединять нуклеотид к неправильно спаренному концу, образовавшемуся при ее участии. На помощь приходит 3'-5' экзонуклеаза, убирающая ошибочный нуклеотид, на место которого затем присоединяется правильный нуклеотид-предшественник. 3'-5' экзонуклеолитическая активность проявляется в направлении, обратном синтезу ДНК (см. рис. 34). Таким образом, 3'-5' экзонуклеазная активность ДНК-полимеразы играет важную роль в точности полимеризации, направляемой матрицей. Эффективность, или число оборотов, данной экзонуклеазы в оптимальных условиях составляет 2 % от числа оборотов субъединицы с полимеразной активностью.
Рис. 34. ДНК-полимераза I Е. coli: а) структура, б) модель взаимодействия с молекулой ДНК (Щелкунов С. Н., 1994)
3. 5'-3' экзонуклеазная активность. Деградирует одну цепь двухцепочечной ДНК, начиная со свободного 5'-конца. В отличие от 3'-5' экзонуклеазы 5'-3' экзонуклеаза расщепляет диэфирную связь только в спаренных участках двухцепочечной молекулы ДНК. Более того, в то время как 3'-5' нуклеаза отщепляет одномоментно только один нуклеотид, 5'-3' нуклеаза может вырезать с 5'-конца олигонуклеотиды длиной до десяти остатков (около 20 % продуктов гидролиза): Скорость нуклеазного отщепления увеличивается на порядок при одновременно протекающей реакции полимеризации. При этом увеличивается относительное количество олигонуклеотидов в продуктах гидролиза ДНК. Такое сочетание ферментативных активностей позволяет ДНК-полимеразе I Е. coli играть активную роль в репарации повреждений ДНК in vivo. N — концевой домен соединен с соседним петлей из аминокислотных остатков и легко отделяется с помощью протеолитических ферментов. Оставшаяся часть бифункциональна, так как состоит из полимеразы и 3'-5' экзонуклезы. Она названа фрагментом Кленова (по фамилии одного из авторов, описавших ее). Фрагмент Кленова (Pol IK) обычно используют для достройки одноцепочечных 5'-концов на двухцепочечной ДНК, часто генерируемых рестриктазами, до тупых; для синтеза второй цепи на одноцепочечной ДНК, а также для гидролиза одноцепочечных 3'-концов на двухцепочечных молекулах ДНК.
Обратная транскриптаза Обратная транскриптаза используется для транскрипции м-РНК в комплементарную цепь ДНК. При изучении ретровирусов, геном которых представлен молекулами одноцепочечной РНК, было обнаружено, что в процессе внутриклеточного развития ретровирус проходит стадию интеграции своего генома в виде двухцепочечной ДНК в хромосомы клетки-хозяина. В 1964 г. Темин выдвинул гипотезу о существовании вирусспецифичного фермента, способного синтезировать на РНК-матрице комплементарную ДНК. Усилия, направленные на выделение такого фермента, увенчались успехом, и в 1970 г. Темин с Мизутани, а также независимо от них Балтимор открыли искомый фермент в препарате внеклеточных вирионов вируса саркомы Рауса. Данная РНК-зависимая ДНК-полимераза получила название обратная транскриптаза, или ревертаза. Наиболее детально изучена ревертаза ретровирусов птиц. Каждый вирион содержит около 50 молекул этого фермента. Обратная транскриптаза состоит из двух субъединиц — а (65 кДа) и Ь (95 кДа), присутствующих в эквимолярном количестве. Обратная транскриптаза обладает, по крайней мере, тремя ферментативными активностями: 1) ДНК-полимеразной, использующей в качестве матрицы как РНК, так и ДНК; 2) активностью РНКазы Н, гидролизующей РНК в составе гибрида РНК — ДНК, но не одно- или двухцепочечную РНК; 3) ДНК-эндонуклеазной активностью. Первые две активности необходимы для синтеза вирусной ДНК, а эндонуклеаза, по-видимому, важна для интеграции вирусной ДНК в геном клетки-хозяина. Очищенная обратная транскриптаза синтезирует ДНК как на РНК-, так и на ДНК-матрицах. Чтобы начать синтез, ревертазе, как и другим полимеразам, необходим короткий двухцепочечный участок (праймер). Праймером может служить одноцепочечный сегмент как РНК, так и ДНК, которые в процессе реакции оказываются ковалентно связанными с новосинтезированной цепью ДНК.
Рис. 35. Схема синтеза двухцепочечных ДНК-копий молекул РНК
Обратную транскриптазу преимущественно используют для транскрипции матричной РНК в комплементарную ДНК (кДНК). Реакцию обратной транскрипции проводят в специально подобранных условиях с использованием сильных ингибиторов РНКазной активности. При этом удается получать полноразмерные ДНК-копии целевых молекул РНК. В качестве праймера при обратной транскрипции поли (А) — содержащих мРНК используют олиго (dT), а для молекул РНК, не имеющих 3'-поли (А) концов, — химически синтезированные олигонуклеотиды, комплементарные 3'-концу изучаемой РНК. После синтеза на мРНК комплементарной цепи ДНК и разрушения РНК (обычно применяют обработку щелочью) осуществляют синтез второй цепи ДНК. При этом используют способность ревертазы образовывать на 3'-концах одноцепочечных кДНК самокомплементарные шпильки, которые могут выполнять функции праймера. Матрицей служит первая цепь кДНК. Данная реакция может катализироваться как ревертазой, так и ДНК-полимеразой I Е. coli. Показано, что сочетание этих двух ферментов позволяет повысить выход полноценных двухцепочечных молекул кДНК. По окончании синтеза первая и вторая цепи кДНК остаются ковалентно связанными петлей шпильки, служившей праймером при синтезе второй цепи. Эту петлю расщепляют эндонуклеазой S1, специфически разрушающей одноцепочечные участки нуклеиновых кислот. Образующиеся при этом концы не всегда оказываются тупыми, и для повышения эффективности последующего клонирования их репарируют до тупых с помощью фрагмента Кленова ДНК-полимераЗы I Е. coli. Полученную двухцепочечную кДНК можно затем встраивать в клонирующие векторы, размножать в составе гибридных молекул ДНК и использовать для дальнейших исследований.
Лигазы В 1961 г. Мезельсон и Вейгл на примере фага 1 показали, что рекомбинация включает разрыв и последующее воссоединение молекул ДНК. Это положило начало поискам фермента, участвующего в сшивании фрагментов ДНК. В 1967 году такой фермент был найден и получил название ДНК-лигаза. Он катализирует синтез фосфодиэфирной связи в 2-х цепочечной молекуле нуклеиновой кислоты. Иными словами, ДНК-лигазы сшивают рядом расположенные нуклеотиды, образуя связь между остатками сахаров. ДНК-лигазы абсолютно необходимы в процессах репарации ДНК, в процессах репликации — при удвоении цепи ДНК. Существует 2 типа ДНК-лигаз, отличающихся по потребностям в кофакторах и способу действия. ДНК-лигаза Е. coli в качестве кофактора использует дифосфопиридиннуклеотид, а лигаза фага Т4 — АТФ в присутствии Мg2+. Лигаза фага Т4 более универсальна, так как помимо лигирования липких концов способна катализировать реакцию воссоединения двухцепочечных фрагментов ДНК с тупыми концами. Она используется чаще.
Ферменты, изменяющие структуру концов фрагментов ДНК Терминальная трансфераза (концевая дезоксинуклеотидилтрансфераза) была обнаружена Боллумом в 1962 году в тимусе теленка. Субстратом терминальной трансферазы при использовании в качестве кофактора ионов Мg2+ является одноцепочечная ДНК с 3'-ОН концом или двухцепочечная ДНК с выступающим одноцепочечным 3'-ОН концом. Если в качестве кофактора используются Со2+, этот фермент может катализировать присоединение дезоксинуклеотидов к 3'-ОН концу двухцепочечной ДНК с тупыми концами. При введении в реакцию, направляемую терминальной трансферазой, лишь одного типа дезоксинуклеотидов образуются молекулы ДНК, имеющие гомополимерные 1-цепочечные 3'-концы. Таким же образом можно достроить другим молекулам ДНК гомополимерные 3'-концы, комплементарные первым. Смешение полученных препаратов ДНК при определенных условиях может приводить к формированию гибридных молекул ДНК. Именно с помощью концевой дезоксинуклеотидилтрансферазы в 1972 г. был выполнен первый эксперимент по рекомбинации молекул ДНК in vitro. Поли (А) — полимераза Е. coli была открыта Сиппелом в 1973 году. Она катализирует присоединение к 3'-ОН концу одноцепочечных молекул РНК поли (А) последовательностей. Применяется при подготовке молекул РНК к копированию с них комплементарной ДНК для введения радиоактивной метки в 3'-конец РНК.
КЛАССИФИКАЦИЯ, НОМЕНКЛАТУРА И ХАРАКТЕРИСТИКА РЕСТРИКТАЗ
Классификация рестриктаз Общепринято термины "рестриктаза", "эндонуклеаза рестрикции" и "сайт специфическая эндодезоксирибонуклеаза" считать синонимами. Все рестрикционные эндонуклеазы бактерий узнают специфические, довольно короткие последовательности ДНК и связываются с ними. Этот процесс сопровождается разрезанием молекулы ДНК либо в самом сайте узнавания, либо в каком-то другом, что определяется типом фермента. Наряду с рестрикционной активностью бактериальный штамм обладает способностью метилировать ДНК; для этого процесса характерна такая же специфичность в отношении последовательностей ДНК, как и для рестрикции. Метилаза добавляет метильные группы к адениновым или цитозиновым остаткам в том же сайте, в котором связывается рестрикционный фермент. В результате метилирования сайт становится устойчивым к рестрикции. Следовательно, метилирование защищает ДНК от разрезания. Различают 3 основных класса рестриктаз: 1, 2 и 3. Все рестриктазы узнают на двуспиральной ДНК строго определенные последовательности, но рестриктазы 1-го класса осуществляют разрывы в произвольных точках молекулы ДНК, а рестриктазы 2-го и 3-го классов узнают и расщепляют ДНК в строго определенных точках внутри сайтов узнавания или на фиксированном от них расстоянии. Ферменты типов 1 и 3 имеют сложную субъединичную структуру и обладают двумя типами активностей — модифицирующей (метилирующей) и АТФ-зависимой эндонуклеазной. Ферменты второго класса состоят из 2 отдельных белков: рестрицирующей эндонуклеазы и модифицирующей метилазы, поэтому в генной инженерии используются исключительно ферменты 2-го класса. Они нуждаются в ионах магния в качестве кофакторов. В настоящее время выделено более 500 рестриктаз класса 2, однако среди ферментов, выделенных из различных микроорганизмов, встречаются такие, которые узнают на ДНК одни и те же последовательности. Такие пары или группы называют изошизомерами. Различают истинную изошизомерию, когда ферменты узнают одну и ту же последовательность нуклеотидов и разрывают ДНК в одних и тех же точках, и ложную, когда ферменты, узнавая один и тот же сайт на ДНК, производят разрывы в разных точках в пределах того же сайта. Большинство рестриктаз класса 2 узнают последовательности, содержащие от 4 до 6 нуклеотидных пар, поэтому рестриктазы делят на мелко- и крупнощепящие. Мелкощепящие рестриктазы узнают тетрануклеотид и вносят в молекулы гораздо больше разрывов, чем крупнощепящие, узнающие последовательность из шести нуклеотидных пар. Это связано с тем, что вероятность встречаемости определенной последовательности из четырех нуклеотидов гораздо выше, чем последовательности из шести нуклеотидов. Например, в ДНК бактериофага Т7, состоящей из 40000 пар оснований, отсутствует последовательность, узнаваемая рестриктазой R1 из Е. coli. К мелкощепящим относятся рестриктазы Нра II и Alu (из Arthrobacter luteus), к крупнощепящим — Eco R I (из Escherichia coli) и Hind III. Если предположить, что участки узнавания рестриктаз распределены вдоль цепи ДНК случайно, то мишень для ферментов, узнающих последовательность (сайт) из четырех нуклеотидов, должна встречаться в среднем 1 раз через каждые 256 пар оснований, а для ферментов, узнающих шесть нуклеотидов, — через 4096 пар оснований. Если сайт рестрикции окажется внутри гена, то обработка ДНК-рестриктазой приведет к его инактивации. Вероятность такого события очень велика при обработке мелкощепящими рестриктазами и незначительна при применении крупнощепящих эндонуклеаз. Поэтому с целью получения неповрежденного гена расщепление проводят поочередно несколькими крупнощепящими рестриктазами, либо применяют прием "недорестрикции", т. е. рестрикцию проводят в таких условиях, когда происходит расщепление лишь в одном сайте.
Номенклатура и характеристика рестриктаз В 1973 году Смит и Натане предложили номенклатуру рестриктаз, включающую следующие пункты: 1. Аббревиатура названия каждого фермента является производной от бинарного названия микроорганизма, содержащего данную метилазно-рестриктазную систему. Составляют по правилу: к первой прописной букве названия рода добавляют две первые строчные буквы вида. Streptomyces albus — Sal, Escherichia coli — Eco 2. В случае необходимости добавляют обозначение серотипа или штамма, например, Есо В. 3. Различные системы рестрикции — модификации, кодируемые одной бактериальной клеткой, обозначают римскими цифрами: Hind II, Hind I, Hind III (Haemophilus influenzae). 4. Рестриктазы обозначают буквой R (R Hind III), метилазы — M (M Hind III). Открытие новых рестриктаз заставило Робертса в 1978 году внести дополнения в систему рациональных обозначений ферментов: если сокращенное название совпадает для нескольких ферментов, то 2 первые буквы аббревиатуры остаются неизменными, а третья берется из последующих букв видового названия: Haemophilus parainfluenzae — Нра I Haemophilus parahaemolyticus — Hph I. Рестриктазы по-разному расщепляют ДНК (рис. 36). Одни вносят разрывы по оси симметрии узнаваемой последовательности (Нра I, Ssp I). Другие — со сдвигом, со "ступенькой" (Pst I). В первом случае образуются так называемые "тупые" концы, а во втором — "липкие", то есть фрагменты имеют на своих концах однонитевые взаимно комплементарные участки длиной в четыре нуклеотида. Такие фрагменты особенно удобны для создания рекомбинантных ДНК.
Рис. 36.
Механизм действия рестриктаз В качестве мишеней (мест узнавания) часто выступают палиндромы из 4–6 пар оснований — сайты рестрикции. Точки узнавания рестриктазами симметричны относительно поворота на 180 °C, то есть последовательность нуклеотидов слева направо в одной нити такая же, как справа налево в другой. Симметрия подразумевает, что те из них, которые должны быть метилированы, встречаются на обеих цепях ДНК. В результате сайт-мишень может быть полностью метилирован (обе цепи модифицированы), полуметилирован (только одна цепь метилирована) или не метилирован. Полностью метилированный сайт не подвержен ни рестрикции, ни модификации. Полуметилированный сайт не узнается ферментом рестрикции, но может быть превращен с помощью метилазы в полностью метилированный. У бактерий метилирование, как правило, связано с сохранением имеющегося состояния модификации. Репликация полностью метилированной ДНК ведет к образованию полуметилированной ДНК. Вероятно, узнавание полуметилированных сайтов представляет собой обычный этап функционирования метилазы in vivo. Неметилированный сайт-мишень представляет собой субстрат либо для рестрикции, либо для модификации in vitro. В клетке немодифицированная ДНК с большей вероятностью рестрицируется. Реакция разрезания осуществляется в две ступени. Сначала разрезается одна цепь ДНК, а затем рядом разрезается другая. В областях, прилегающих с каждой стороны к сайту разрезания, может иметь место экзонуклеотическая деградация. Происходит эффективный гидролиз АТФ, роль которого еще не выяснена. Каким образом фермент узнает один сайт, а разрезает другой, достаточно удаленный? Важно отметить, что белок никогда не отделяется от молекулы ДНК, с которой он первоначально связался. Если фермент инкубировать со смесью модифицированной и немодифицированной ДНК, он предпочтительно разрезает немодифицированную ДНК. Следовательно, узнавая сайт связывания, белок не отделяется от неметилированной ДНК для того, чтобы найти сайт разрезания. Существуют две альтернативные модели, объясняющие взаимосвязь между сайтами узнавания и разрезания: в соответствии с одной из них движется фермент, согласно другой модели, перемещается ДНК. Если движется фермент, то его перемещение вдоль ДНК будет продолжаться до тех пор, пока он не сделает выбор сайта разрезания. Если же движется ДНК, то фермент остается прикрепленным в сайте узнавания, а ДНК протаскивается через второй сайт связывания на ферменте, и это продолжается до тех пор, пока фермент не достигает области разрезания (пока не охарактеризованной). Получены электронно-микроскопические данные, свидетельствующие, что фермент вызывает образование петли в ДНК и остается, по-видимому, связанным с сайтом узнавания после разрезания; эти данные подтверждают вторую модель.
ПОСТРОЕНИЕ РЕСТРИКЦИОННЫХ КАРТ
Ферменты рестрикции стали эффективным инструментом исследования. Они позволяют превращать молекулы ДНК очень большого размера в набор фрагментов длиной от нескольких сотен до нескольких тысяч оснований. С помощью метода электрофореза в агарозном геле (см. раздел 1) фрагменты ДНК, различающиеся по размеру, можно легко разделить, а затем исследовать каждый фрагмент отдельно. Короткие фрагменты мигрируют намного быстрее, чем длинные. При сравнительно высокой концентрации агарозы большие фрагменты вообще не могут проникнуть в гель. В процессе миграции рестрикционные фрагменты не деградируют, их можно элюировать (вымывать) в виде биологически активных двуцепочечных молекул. При окрашивании гелей красителями, связывающимися с ДНК, выявляется набор полос, каждая из которых отвечает рестрикционному фрагменту, молекулярную массу которого можно определить, проведя калибровку с помощью ДНК с известными молекулярными массами. Сравнение размеров фрагментов ДНК, полученных после обработки определенного участка генома набором рестрицирующих нуклеаз позволяет построить рестрикционную карту, на которой указано положение каждого сайта рестрикции относительно других участков. Молекулу ДНК длиной 5000 пар нуклеотидов (п. н.). обрабатывают отдельно рестриктазами А и В. Фрагменты разделяют электрофорезом. Фермент А разрезал ДНК на 4 фрагмента размером 2100, 1400, 1000 и 500 п. н. Обработка рестриктазой В дала 3 фрагмента: 2500, 1300 и 1200 п. н. (рис. 37). Для определения расположения сайтов рестрикции этих ферментов на следующем этапе применяют процедуру двойного расщепления — обрабатывают ДНК двумя эндонуклеазами. Обработка изучаемого фрагмента одновременно двумя рестриктазами дала 6 фрагментов: 1900,
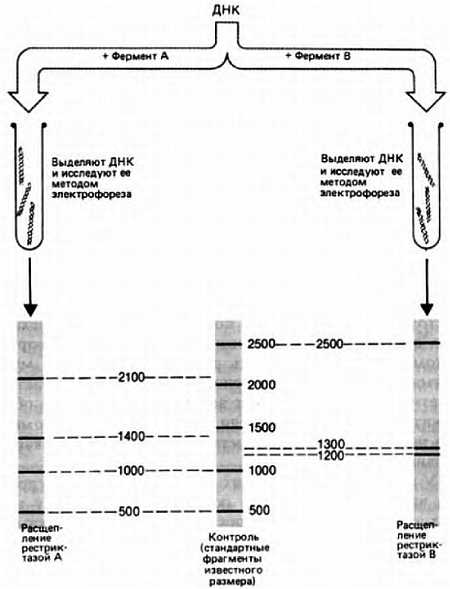
Рис. 37. Результаты электрофореза после обработки фрагмента ДНК разными рестриктазами.
Наиболее полный вариант — элюировать каждый фрагмент, образующийся в результате расщепления одной рестриктазой, а затем обработать его второй. Смесь фрагментов, полученных после такой обработки, также анализируют с помощью электрофореза. В нашем примере были получены следующие результаты: Обработка каждого из 4-х A-фрагментов рестриктазой В
2100–1900 и 200, 1400 — 800 и 600, 1000–1000 (изменений нет) 500 — 500 (изменений нет)Обработка каждого из 3-х В-фрагментов рестриктазой А
2500–1900 и 600 1300 — 800 и 500 1200–1000 и 200Анализ полученных результатов показывает, что каждый из ферментов, полученный при расщеплении A-фрагментов рестриктазой В можно обнаружить в образцах, полученных при расщеплении В-фрагментов рестриктазой А. Ключом к рестрикционному картированию являются перекрывающиеся фрагменты. Такими в рассматриваемом примере являются В-фрагмент 2100 и А-фрагмент 2500. При обработке другой рестриктазой они дают фрагмент 1900. Из данных о расщеплении этих фрагментов мы предполагаем, что с одной стороны на расстоянии 200 п.н. от фрагмента 1900 находится следующий A-сайт, а с другого конца, на расстоянии 600 п.н. — следующий В-сайт (рис. 38).
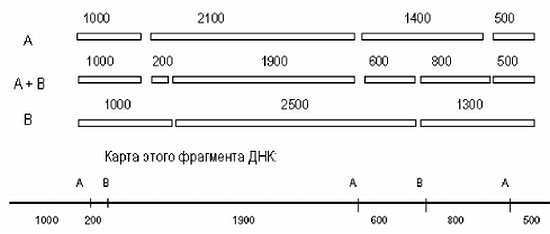
Рис. 38. Анализ фрагментов рестрикции и карта фрагмента ДНК
При обработке двумя эндонуклеазами фрагмент 200 п. н. образуется 1 раз, при обработке рестриктазой А из В-фрагмента 1200, т. е. фрагмент 1200 лежит слева. Остается определить, как продолжается карта вправо. Очевидно, это А-фрагмент 1400, так как он рассечен рестриктазой В на фрагменты 600 и 800. Вправо от фрагмента 2500 следует отложить, очевидно, фрагмент 1300. Тогда логично наличие А-фрагмента 500 и деления В-фрагмента 1300 рестриктазой А на 800 и 500. При построении рестрикционных карт обычно используют несколько рестриктаз, поэтому приходится анализировать сложные соотношения между фрагментами, полученными при действии разных ферментов. Для упрощения процедуры картирования можно применять неполное расщепление. В определенных условиях рестриктаза узнает и расщепляет не все сайты в молекуле ДНК. Например, при частичном расщеплении ДНК ферментом А могут образовываться фрагменты 3100 п.н., 1400 п.н. и 500 п.н. Сопоставив их с данными полного расщепления (2100, 1400, 1000 и 500), можно сразу поставить рядом 2100 и 1000 (фрагмент 3100). А получив фрагмент 3500 — расположить рядом 2100 п.н. и 1400 п.н. Другой прием — введение радиоактивной концевой метки. Концевые фрагменты определяются в этом случае по включению метки. Можно также сопоставить фрагменты путем гибридизации нуклеиновых кислот. Перекрывающиеся фрагменты (в данном случае 2100 и 2500) будут гибридизоваться. Первая карта была получена для вируса SV40 (обезьяний вирус, вызывающий злокачественную трансформацию), содержащего 5423 пары оснований. Использовали рестриктазу Hind-II, расщепляющую кольцевую ДНК вируса на 11 фрагментов. Порядок их расположения в ДНК был установлен путем исследования наборов фрагментов, образующихся по мере того, как расщепление доходит до конца. Первый разрыв превращал кольцевую молекулу в линейную, которая затем расщеплялась на все меньшие и меньшие фрагменты. Исследовали вначале наборы перекрывающихся фрагментов, а затем продукты полного расщепления. Таким образом, была получена рестрикционная карта кольцевой вирусной ДНК, на которую были нанесены сайты расщепления рестриктазой. Повторив подобные эксперименты с другой рестриктазой можно получить более подробную карту, где отмечено много сайтов рестрикции. Располагая такой информацией, можно идентифицировать на ДНК биологически важные участки. Поскольку рестрикционная карта отражает расположение определенной последовательности нуклеотидов в данном участке, сравнение таких карт для двух или более родственных генов позволяет оценить гомологию между ними. Анализируя рестрикционные карты, можно сравнивать определенные участки ДНК разных видов животных без определения их нуклеотидной последовательности. Таким образом, например, было установлено, что хромосомные участки,кодирующие цепи гемоглобина у человека, орангутанга и шимпанзе сохранились в практически неизменном виде в течение последних 5-10 млн. лет (с тех пор как виды дивергировали). Метод рестрикционного картирования позволяет увидеть крупные генетические изменения, такие как делеции или инсерции. При этом происходит уменьшение или увеличение рестрикционных фрагментов, а также исчезновение или возникновение сайтов рестрикции. Один из приемов картирования — фингерпринт («метод отпечатков пальцев» или DNA-fingerprint). Он подразумевает использование неупорядоченных и неполных наборов фрагментов, которые являются характеристикой генома, хотя описывает его не полностью.
КОНСТРУИРОВАНИЕ РЕКОМБИНАНТНЫХ ДНК
Под рекомбинантными понимают ДНК, образованные объединением in vitro (в пробирке) двух или более фрагментов ДНК, выделенных из различных биологических источников. Ключевыми в этом определении являются слова "фрагмент ДНК" и "объединение in vitro", что указывает на сущность генетической инженерии и ее отличие от всех остальных методов получения гибридных (или химерных) организмов, таких как генетическая селекция, эмбриональная инженерия и т. д. Фрагменты ДНК, в том числе и фрагменты, содержащие гены, получают с использованием ферментов рестриктаз. Рестриктазы могут образовывать фрагменты, как с тупыми, так и с липкими концами. Сшивка фрагментов ДНК производится тремя основными методами, зависящими от того, какие концы имеют фрагменты сшиваемых ДНК.
Сшивка по одноименным "липким" концам (рестриктазно лигазный метод) Этот метод является самым распространенным и популярным. Впервые этим способом гибридная ДНК была получена С. Коэном с сотрудниками в 1973 году. Не которые рестриктазы, например Pst I, внося в цепи ДНК симметричные, расположенные наискось друг от друга разрывы на равных расстояниях от центра сайта узнавания и образующие "ступеньку" (рис. 36). Эти комплементарные друг другу участки имеют тенденцию к ассоциации за счет спаривания оснований, и поэтому их называют комплементарными или липкими концами. Спаривание оснований происходит только между комплементарными последовательностями, поэтому ААТТ-концы, образуемые Eco RI, не будут спариваться, например, с АГЦТ-концами, образуемыми Hind III. Но любые два фрагмента (независимо от их происхождения), образовавшиеся под действием одной и той же рестриктазы, могут слипаться за счет образования водородных связей между однонитиевыми участками комплементарных нуклеотидов (рис. 39).

Рис. 39. Схема рестриктазно-лигазного метода
Однако после такого спаривания полной целостности двойной спирали не восстановится, поскольку останется два разрыва в фосфодиэфирном остове. Для его восстановления, то есть сшивания, или лигирования нитей используют фермент ДНК-лигазу. Этот фермент в живой клетке выполняет ту же функцию — сшивание фрагментов ДНК, синтезирующихся при репликации.
Сшивка по "тупым" концам (коннекторный метод) Липкие концы не абсолютно необходимы для связывания фрагментов ДНК. Тупые концы также могут быть соединены за счет действия ДНК-лигазы, если и лигаза, и тупые концы присутствуют в реакционной смеси в высоких концентрациях. В этом случае реакция лигирования имеет свои особенности и ее эффективность ниже, чем при сшивке по липким концам. Впервые такие эксперименты были выполнены в 1972 году Полем Бергом в Стенфордском университете, США. Липкие концы также можно ферментативным путем присоединить к молекулам ДНК с тупыми концами. Для этого используют фермент — концевую трансферазу из тимуса теленка, которая присоединяет нуклеотиды к 3-концам цепей ДНК. Если к 3'-концам одного из рекомбинируемых in vitro фрагментов ДНК с помощью концевой дезоксинуклеотидилтрансферазы достроить одноцепочечные олиго (dA) — сегменты определенной длины, а к концам другого фрагмента — олиго (dT) — сегменты примерно такой же длины, то при смешении полученных таким образом фрагментов происходит спаривание за счет образования водородных связей между олиго (dA) — и олиго (dT) — последовательностями (рис. 40). Для ковалентного соединения двух фрагментов используется ДНК-лигаза. Эти процедуры составляют основу для второго общего метода получения рекомбинантных молекул ДНК.

Рис. 40. Пришивание «липких» концов и сшивка фрагментов ДНК
Поскольку можно формировать достаточно длинные взаимокомплементарные одноцепочечные концы, гибридные молекулы образуются с высокой эффективностью. В частности, поэтому при клонировании ДНК-копий матричных РНК, которые доступны в ограниченных количествах, обычно используют коннекторный метод. При таком способе соединения между фрагментами встраиваются участки ААААА. Такие дополнительные последовательности ТТТТТ могут влиять на функции соединяемых молекул и поэтому всегда, когда только возможно, для получения рекомбинантных молекул ДНК пользуются липкими концами, образовавшимися в результате действия рестриктаз.
Сшивка фрагментов с разноименными липкими концами В ситуации, когда необходимо сшить фрагменты, образованные разными эндонуклеазами рестрикции, и имеющие разные, то есть некомплементарные друг другу липкие концы, применяют так называемые линкеры (или "переходники"). Линкеры — это химически синтезированные олигонуклеотиды, представляющие собой сайты рестрикции или их комбинацию. Впервые эту идею предложил Шеллер с сотрудниками в 1977 году. Существуют большие наборы таких генных "переходников". Естественно, что при использовании линкеров должна учитываться необходимость соблюдения правил экспрессии генетической информации. Часто в середину линкера помещают какой-либо регуляторный генетический элемент, например, промотор или участок, связанный с рибосомой. В этом случае линкеры обеспечивают не только объединение генов, но и обуславливают их экспрессию. Существуют линкеры "тупой конец — липкий конец". При необходимости липкие концы можно превратить в тупые. Это достигается либо отщеплением липких концов с помощью фермента — эндонуклеазы S1, которая разрушает только одноцепочечную ДНК, либо липкие концы "застраивают", то есть с помощью ДНК-полимеразы I на однонитевых липких концах синтезируют вторую нить.
ОПРЕДЕЛЕНИЕ НУКЛЕОТИДНОЙ ПОСЛЕДОВАТЕЛЬНОСТИ (СЕКВЕНИРОВАНИЕ) ДНК
Описанные методы, позволившие идентифицировать генетически важные участки ДНК, имели большое значение сами по себе. Но они также проложили путь к разработке исключительно эффективных методов секвенирования ДНК и создания рекомбинантных молекул. Секвенирование позволяет довольно быстро определить полную нуклеотидную последовательность сегмента длиной 100–500 нуклеотидных пар, образующегося при расщеплении ДНК рестрикционными эндонуклеазами.
Метод Маскама и Гилберта (химический) Один из методов основан на химической деградации ДНК. Он был предложен в 1976 году Максамом и Гилбертом и назван их именем. Суть метода сводится к следующему: один из концов фрагмента ДНК метят с помощью изотопа фосфора 32Р. В последнее время вместо радиоактивной вводят флюоресцирующую метку. Ее можно «цеплять» и к нуклеотидам, причем для каждого типа нуклеотидов подбирать различную окраску. Препарат меченой ДНК делят на четыре порции и каждую из них обрабатывают реагентом, специфически разрушающим одно или два из четырех оснований, причем условия реакции подбирают таким образом, чтобы на каждую молекулу ДНК приходилось лишь несколько повреждений. Разрушение идет в 2 этапа. На первом этапе происходит модификация азотистого основания и последующее выщепление его. На втором этапе производят гидролиз ДНК в местах выщепления оснований. Пуриновые основания модифицируются диметилсульфатом. Адениновые остатки метилируются по третьему атому азота, гуаниновые — по положению N7. Если такую модификацию обработать 0,1 М НСl при 0 °C, то выщепляется метиладенин. При последующей инкубации в щелочной среде (0,1 М NaOH) при температуре +90 °C происходит разрушение сахаро-фосфатной связи в местах выщепления оснований. Обработка поврежденных молекул пиперидином приводит к гидролизу ДНК по остаткам метилгуанина. Пиримидиновые основания модифицируются гидразином. В бессолевой среде модифицируется и цитозин, и тимин, в присутствии 2 М NaCl модифицируется только цитозин. При дальнейшей обработке пиперидином происходит расщепление ДНК по точкам модификации. Можно использовать и другие реакции химической модификации оснований и расщепления по ним молекул ДНК. В результате получается набор меченых фрагментов, длины которых определяются расстоянием от разрушенного основания до конца молекулы. Фрагменты, образовавшиеся во всех четырех реакциях, подвергают электрофорезу в четырех соседних дорожках; затем проводят радиоавтографию, и те фрагменты, которые содержат радиоактивную метку, оставляют "отпечатки" на рентгеновской пленке. По положению отпечатков можно определить, на каком расстоянии от меченого конца находилось разрушенное основание, а зная это основание — его положение. Так набор полос на рентгеновской пленке определяет нуклеотидную последовательность ДНК. Аналогично наблюдают флюоресцентное окрашивание. Если для каждого из четырех нуклеотидов был подобран свой цвет флюоресцентной метки, то при электрофорезе их наносят на 1 дорожку. Тогда расположение нуклеотидов отмечено штрихами разного цвета, а процедуру считывания легко автоматизировать.
Метод Сэнгера (ферментативный) Другой метод, разработанный Сэнгером и носящий его имя, основан не на химическом, а на ферментативном подходе. Сэнгер использовал ДНК-полимеразу I. В клетке этот фермент участвует в процессе репликации, заполняя пробелы между вновь синтезированными фрагментами ДНК (фрагментами Оказаки). Для работы фермента в пробирке требуются предшественники ДНК — дезоксирибонуклеотидтрифосфаты (dNTP), а также одноцепочечная матрица, на которой должен быть небольшой двухцепочечный участок — затравка, с которого начинается синтез (рис. 41). Были также синтезированы модифицированные дидезоксирибонуклеотиды, в которых дезоксирибоза 3'-ОН отсутствует, для каждого из четырех оснований ДНК. ДНК-полимераза включает эти предшественники в ДНК. Однако, включившись в ДНК, модифицированное основание не может образовать фосфодиэфирную связь со следующим дезоксирибонуклеотидом. В результате рост (элонгация) данной цепи останавливается (терминируется) в том месте, где в ДНК включился дидезоксирибонуклеотид (ddNTP). Поэтому их называют терминаторами элонгации.
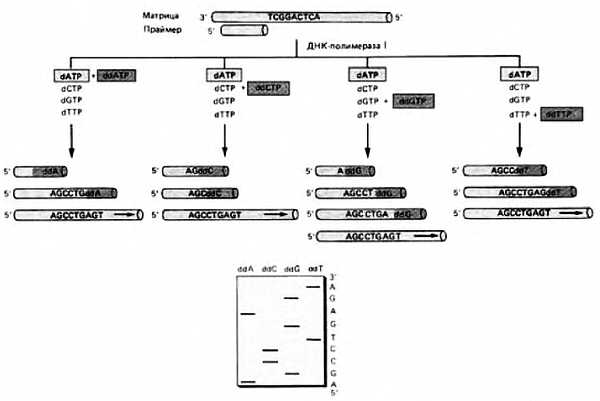
Рис. 41. Ферментативный метод секвенирования ДНК
Реакционная смесь по Сэнгеру состоит из цепи ДНК, нуклеотидную последовательность которой надо определить, короткого фрагмента "меченой" ДНК, комплементарной концевому отрезку этой цепи (затравка), одного из четырех ddNTP и соответствующего dNTP в строго определенном соотношении (чтобы они конкурировали), а также остальных трех dNTP. Готовят четыре смеси, каждая из которых содержит один из четырех ddNTP. В каждой из пробирок образуется набор меченых фрагментов разной длины. Длина их зависит от того, в каком месте в цепь включен дефектный нуклеотид. Полученные меченые фрагменты ДНК разделяют в полиакриламидном геле (с точностью до одного нуклеотида), проводят радиоавтографию и по картине распределения фрагментов в четырех пробах устанавливают нуклеотидную последовательность ДНК (рис. 41). В настоящее время определение точной нуклеотидной последовательности любого сегмента ДНК умеренной длины — вполне разрешимая задача. Уже определена последовательность нескольких сотен генов про- и эукариот. Зная последовательность гена и генетический код, легко определить аминокислотную последовательность кодируемого им белка. Раньше для определения структуры белка приходилось делать тщательный и весьма трудоемкий анализ выделенного и очищенного белка. Сейчас часто бывает проще определить структуру белка через нуклеотидную последовательность, чем с помощью прямого секвенирования. Если секвенирование белка занимает месяцы и даже годы, то ДНК удается секвенировать за несколько недель. Определение последовательности ДНК привело также к тому, что были обнаружены области, которые не кодируют белки, но принимают участие в регуляции экспрессии генов и репликации ДНК. В 1996 году был секвенирован геном дрожжей, в 1998 г. — геном арабидопсиса, в 2000 году — геном человека, однако в данном случае речь идет только об установлении последовательности нуклеотидов, так как генетическая структура и функции отдельных участков генома еще не идентифицированы, это более сложная задача. Сразу вслед за разработкой быстрых методов секвенирования появились столь же быстрые и простые методы синтеза сравнительно длинных олигонуклеотидов с определенной, заранее заданной последовательностью. Теперь за три-четыре дня можно синтезировать последовательность из 12–20 нуклеотидов. Автоматизация этой процедуры еще более облегчает и ускоряет синтез. Появились приборы — ДНК-синтезаторы, которые выполняют эту работу за несколько часов.
ГИБРИДИЗАЦИЯ КАК ВЫСОКОЧУВСТВИТЕЛЬНЫЙ МЕТОД ВЫЯВЛЕНИЯ СПЕЦИФИЧЕСКИХ ПОСЛЕДОВАТЕЛЬНОСТЕЙ НУКЛЕОТИДОВ
Если водный раствор ДНК нагреть до 100 °C и повысить pH до 13, то ДНК диссоциирует на 2 цепи (денатурирует), так как комплементарные связи между основаниями разрушаются. В 1961 году было обнаружено, что этот процесс обратим: выдерживание ДНК при температуре 65 °C вело к восстановлению структуры двойной спирали. Этот процесс называется ренатурация или гибридизация. Процессы гибридизации происходят между любыми одинарными цепями, если они комплементарны: ДНК-ДНК, РНК-РНК, ДНК-РНК. Для теста необходимо иметь чистый одноцепочечный фрагмент ДНК, комплементарный той последовательности, которую хотим обнаружить. Этот фрагмент получают либо клонированием, либо путем химического синтеза. Одноцепочечная ДНК, используемая в качестве индикатора, называется ДНК-зонд. Она может содержать от 15 до 1000 нуклеотидов. ДНК-зонды применяются в различных целях. Гибридизация ДНК-зонда с РНК, выделенной из анализируемой клетки, может выявить наличие или отсутствие экспрессии гена. Если гибридизации не происходит, значит, ген молчит, не работает. ДНК-зонды также позволяют проводить диагностику наследственных болезней. В большинстве случаев мутации, ведущие к наследственным болезням, рецессивны, то есть болезнь развивается, если человек получает дефектные гены от обоих родителей. Аномальные эмбрионы лучше выявлять до рождения. Например, для серповидноклеточной анемии в мутантном гене, кодирующем бэта-цепь гемоглобина, последовательность ГАГ заменена на ГТГ. В этом случае синтезируют олигонуклеотид длиной около 20 оснований, метят радиоактивной меткой. Из эмбриональных клеток, содержащихся в амниотической жидкости, выделяют ДНК и используют ее для гибридизации. Если эмбрион дефектен, то тест будет положительным. Анализ проводят по следующей схеме (рис. 42): исследуемую ДНК гидролизуют рестриктазами, фракционируют электрофорезом, переносят разделенные фрагменты на нитроцеллюлозный фильтр и проводят реакцию гибридизации с мечеными олигонуклеотидами. Этот метод был разработан Саузерном в 1975 году. В отечествен ной литературе его принято называть «южный блоттинг». «Блоттинг» — в переводе с английского означает «промокашка», «саузерн» — «южный», в данном случае игра слов: фамилия ученого переводится как географическое направление.

Рис. 42. Блоттинг ДНК по Саузерну
Молекулы ДНК разгоняют в агарозном геле электрофорезом. ДНК в геле денатурируют щелочью. Щелочь нейтрализуют и пластину геля покрывают листом нитроцеллюлозы. Сверху на нитроцеллюлозу помещают стопку листов фильтровальной бумаги, обеспечивая медленный ток буферного раствора через гель в направлении, перпендикулярном направлению электрофореза. ДНК диффундирует из геля и связывается с нитроцеллюлозным фильтром. После прогревания фильтра при 80 °C в вакууме ДНК необратимо связывается с нитроцеллюлозой. Расположение полос иммобилизованной ДНК точно соответствует их расположению в геле. ДНК, связанную с нитроцеллюлозным фильтром, можно гибридизовать с радиоактивно меченой ДНК. Блоттинг по Саузерну является исключительно полезным также и для локализации изучаемых генов в определенных фрагментах, полученных в результате гидролиза различными рестриктазами гибридных молекул ДНК, хромосомной ДНК и т. д. Аналогичным методом на нитроцеллюлозу переносят молекулы РНК (Северный блоттинг) и белка (Западный блоттинг). Из частей света осталась свободной только западная, хотя вакантными остались 2 класса макромолекул — углеводы и липиды. Одним из наиболее точных и современных методов анализа является использование чипов. Они представляют собой пластинки с иммобилизованными меченными ДНК-зондами. Каждая такая пластинка может содержать несколько десятков тысяч зондов, расположенных в определенной последовательности. Метка проявляется только в спаренных двухцепочечных фрагментах. Если в исследуемом образце есть последовательности, комплементарные последовательностям зонда, то гибридизацию можно определить визуально или с помощью специальных приборов. Как правило, детекторы соединены с компьютером, то есть процедура считывания и обработки информации автоматизирована. Такие ДНК-чипы можно применять для комплексной диагностики инфекционных заболеваний, наследственных дефектов, установления экспрессии тех или иных генов (в этом случае идет гибридизация с мРНК), то есть отслеживания нарушений обмена веществ. Они дешевы, очень надежны, просты в обращении и могут многократно использоваться. Недостаток — дорогая аппаратура для детекции.
МЕТОДЫ КЛОНИРОВАНИЯ ДНК
После того, как ДНК сшита в пробирке, ее необходимо размножить. Существует два подхода к клонированию ДНК. Первый подход предполагает использование бактериальных или дрожжевых клеток для размножения введенной в них чужеродной ДНК. Второй способ представляет собой амплификацию ДНК in vi tro.
Клонирование ДНК in vivo Используя микроорганизмы, можно создавать два типа библиотек ДНК: геномную и клоновую (кДНК).
Геномная библиотека Если геном какого-либо организма разрезать, вставить в плазмидные или вирусные вектора и ввести в клетку, то в таком виде его можно сохранить. При разрезании плазмидной или фаговой ДНК вероятность выпадения целых и неизмененных кусков генома довольно высока. Такой способ получения геномной библиотеки получил название «метод дробовика», так как геном в данном случае представлен отдельными фрагментами. Принципы создания плазмидных и вирусных векторов общие, поэтому рассмотрим их на примере плазмидных. Следует отметить, что из вирусных ДНК лучше использовать ДНК фагов, так как они имеют большую емкость и позволяют вставлять более крупные куски генома. Очищенные кольцевые молекулы ДНК обрабатывают рестриктазой, получая линейную ДНК. Клеточную ДНК обрабатывают той же рестриктазой, добавляют к плазмидной, добавляют лигазы. Таким образом, получают рекомбинантную плазмидную ДНК, которую вводят в бактериальные или дрожжевые клетки. Плазмида реплицируется с образованием многих копий. Многие плазмиды несут ген устойчивости к антибиотикам, и если в рекомбинантной плазмиде есть такой ген, то клетки легко выявлять, выращивая на среде с антибиотиком. Каждая такая колония представляет собой клон или потомство одной клетки. Плазмиды одной колонии содержат клон геномной ДНК, а совокупность плазмид можно назвать библиотекой геномной ДНК. Недостаток такого метода в том, что фрагменты ДНК образуются в огромном количестве. Разрезание геномной ДНК определяется случаем, поэтому лишь часть фрагментов содержат полноценные гены. Некоторые фрагменты могут содержать только часть гена или же интронные последовательности.
Библиотека кДНК Создание кДНК начинается с синтеза на матрице РНК с помощью обратной транскриптазы комплементарной нити ДНК. Затем создают щелочные условия, разрушают цепь РНК на нуклеотиды, после чего с помощью ДНК-полимеразы синтезируют комплементарную цепь ДНК. При этом образуется фрагмент ДНК с тупыми концами. Такую ДНК встраивают в плазмиды и вводят в клетки бактерий. При амплификации плазмиды образуется клон комплементарной копии ДНК (кДНК). Преимущества клоновой ДНК перед клонами геномной ДНК в том, что кодирующая белок нуклеотидная последовательность гена ничем не прерывается. Гены эукариот содержат интроны, которые должны удаляться из транскриптной РНК перед превращением ее в матричную, после чего следует сплайсинг (сращивание). Бактериальные клетки не могут осуществлять такую модификацию РНК, образовавшуюся путем транскрипции гена эукариотической клетки. Поэтому если преследуют получение белка путем экспрессии клонированного гена, то лучше использовать банк кДНК, полученной на основе матричной РНК.
Полимеразная цепная реакция В 1985 году К. Мюллис с сотрудниками разработали метод клонирования последовательностей ДНК in vitro, который получил название полимеразной цепной реакции (ПЦР). К анализируемому образцу ДНК добавляют в избытке 2 синтетических олигонуклеотида — праймера размером около 20 нуклеотидов. Каждый из них комплементарен одному из 3'-концов фрагмента ДНК. ДНК нагревают для разделения цепей двойной спирали, а при охлаждении происходит гибридизация праймеров с комплементарными участками фрагментов ДНК. В результате в растворе будут находиться однонитевые ДНК с короткими двухцепочечными участками — затравками (праймерами). При добавлении нуклеотидов и ДНК-полимеразы синтезируются комплементарные цепи и образуются идентичные фрагменты ДНК (первый цикл, рис. 43). Реакция останавливается и ДНК снова денатурируется прогреванием.

Рис. 43. Первый цикл полимеразной цепной реакции
В процессе охлаждения праймеры, находящиеся в избытке, вновь эффективно гибридизуются, но уже не только с цепями исходной ДНК, но и с вновь синтезированными. Внесение в систему ДНК-полимеразы инициирует второй цикл полимеразной реакции. Многократное повторение описанной процедуры позволяет провести 30 и более циклов ферментативного удлинения праймеров. При этом число сегментов ДНК, ограниченных с обоих концов используемыми праймерами, с каждым циклом ПЦР увеличивается экспоненциально (приближается к зависимости 2n, где n — число циклов). Выход всех других продуктов реакции увеличивается по линейной зависимости (рис. 44). Таким образом, в процессе рассматриваемой реакции эффективно амплифицируется только та последовательность ДНК, которая ограничена праймерами.
Рис. 44. Схема полимеразной цепной реакции
Первоначально для ПЦР использовали фрагмент Кленова ДНК-полимеразы I Е. coli. Однако недостатком данного подхода являлось то, что после каждого цикла реакции необходимо было вносить в реакционную смесь новую порцию фермента. Кроме того, в оптимальных температурных условиях такой полимеразной реакции (37 °C) появлялись вторичные участки связывания праймеров и наблюдалась амплификация незапланированных сегментов генома, т. е. специфичность амплификации не была полной. Существенное улучшение метода полимеразной цепной реакции было достигнуто после замены фрагмента Кленова на ДНК-полимеразу термофильной бактерии Thermus aquaticus (Taq-полимераза). Температурный оптимум реакции, направляемой Taq-полимеразой, находится в районе 70 °C. Другим важным свойством является то, что данная полимераза не инактивируется после длительной инкубации при 95 °C. Используя Taq-полимеразу, удалось решить сразу две проблемы. Во-первых, термостабильная полимераза не инактивируется на этапе денатурации ДНК, и поэтому нет необходимости после каждого цикла реакции добавлять новую порцию фермента. Такое упрощение процедуры позволило автоматизировать проведение ПЦР, так как теперь требовалось лишь перенесение образца с определенным интервалом времени в разные температурные условия: 90–95 °C (температура денатурации) и 60–70 °C (температура ренатурации ДНК и ферментативной реакции). Во-вторых, высокий температурный оптимум реакции, катализируемой Taq-полимеразой, позволяет подбирать жесткие температурные условия отжига, обеспечивающие гибридизацию праймеров только в заданном районе изучаемого генома, что существенно повышает специфичность и чувствительность метода. Используя метод ПЦР, можно in vitro селективно обогащать препарат ДНК фрагментом с определенной последовательностью в миллион и более раз. Это позволяет надежно выявлять однокопийные гены и их варианты в таких больших и сложных геномах, каким является геном человека. Чувствительность метода такова, что амплифицировать в ПЦР и выявить целевую последовательность можно даже в том случае, если она встречается однажды в образце из 105 клеток. Получаемый сегмент ДНК надежно выявляется в виде дискретной полосы после электрофоретического разделения молекул ДНК и окраски их этидиум бромидом. Если к праймеру пришить фермент, то ферментная метка будет накапливаться при амплифицировании. Продукт амплификации проверяется по принципу ИФА, то есть добавляется субстрат и отмечается изменение окраски. В качестве метки можно использовать стрептавидин. Его можно пришивать как к праймеру, так и к нуклеотидам. В последнем случае нуклеотиды, меченные стрептавидином, добавляются к обычным, идущим на синтез комплементарной цепи ДНК. Этим достигается еще большее усиление сигнала. Размноженный in vitro фрагмент получают в количествах, достаточных для его прямого секвенирования. Поскольку при этом не требуется промежуточный этап клонирования фрагмента ДНК в молекулярных векторах, ПЦР иногда называют бесклеточным молекулярным клонированием (cell-free molecular cloning). Автоматизированная процедура Taq-полимеразной цепной реакции, состоящая из 30 и более циклов, занимает 3–4 часа, что существенно быстрее и проще процедуры клонирования определенного фрагмента ДНК в составе векторных молекул.
Применение метода полимеразной цепной реакции Полимеразную цепную реакцию используют для анализа индивидуальных вариаций последовательности нуклеотидов определенных локусов, для повышения эффективности клонирования целевых последовательностей изучаемых геномов и их прямого секвенирования, для детекции в организме патогенных микроорганизмов и т. п. Используя 32Р-меченные синтетические олигонуклеотиды, можно выявлять единичные замены нуклеотидов в выбранных локусах геномной ДНК человека (или других организмов). Для этого в обычном варианте метода исследуемую ДНК гидролизуют рестриктазами, фракционируют электрофорезом, переносят разделенные фрагменты по Саузерну на нитроцеллюлозный фильтр, который гибридизуют с данным меченым олигонуклеотидом в условиях, при которых даже точечная замена нуклеотидов в анализируемой последовательности приводит к разрушению комплекса ДНК-олигонуклеотид. Использование полимеразной цепной реакции для амплификации анализируемого локуса позволяет существенно упростить рассмотренный подход и повысить его чувствительность и специфичность. При этом для анализа аллельных вариантов генов достаточно всего 1 нг геномной ДНК человека, а гибридизацию можно проводить с негидролизованной рестриктазами ДНК, иммобилизованной на нитроцеллюлозном фильтре в виде небольшого пятна. Такой вариант метода позволил разработать новые диагностические тесты на генетические и инфекционные заболевания. В частности, этот подход используют для ранней диагностики наличия в организме вируса иммунодефицита человека (ВИЧ), что не удается осуществить другими методами. При этом не требуется работать с радиоактивными изотопами, так как амплифицированный сегмент вирусной ДНК выявляется напрямую после электрофоретического разделения ДНК и окраски их бромистым этидием. Метод ПЦР позволил проанализировать наличие последовательностей вирусов папилломы человека в срезах биопсий новообразований шейки матки человека, залитых парафином за 40 лет до данного исследования. Более того, с помощью ПЦР удалось амплифицировать и клонировать фрагменты митохондриальной ДНК из ископаемых останков мозга человека возраста 7 тысяч лет! На лизатах индивидуальных сперматозоидов человека продемонстрирована возможность одновременно анализировать два локуса, расположенных на разных негомологичных хромосомах. Такой подход обеспечивает уникальную возможность тонкого генетического анализа и изучения хромосомной рекомбинации, ДНК-полиморфизма и др. Метод анализа индивидуальных сперматозоидов сразу нашел практическое применение в судебной медицине, так как HLA-типирование гаплоидных клеток позволяет определять отцовство или выявлять преступника (комплекс HLA представляет собой набор генов главного комплекса гистосовместимости человека; локусы комплекса HLA — наиболее полиморфные из всех известных у высших позвоночных: в пределах вида в каждом локусе существует необычайно большое число разных аллелей — альтернативных форм одного и того же гена). Используя ПЦР, можно выявлять правильность интеграции чужеродных генетических структур в заранее определенный район генома изучаемых клеток. Суммарная клеточная ДНК отжигается с двумя олигонуклеотидными затравками, одна из которых комплементарна участку хозяйской ДНК вблизи точки встраивания, а другая — последовательности интегрированного фрагмента в антипараллельной цепи ДНК. Полимеразная цепная реакция в случае неизмененной структуры хромосомной ДНК в предполагаемом месте встройки приводит к образованию фрагментов одноцепочечной ДНК неопределенного размера, а в случае запланированной встройки — двухцепочечных фрагментов ДНК известного размера, определяемого расстоянием между местами отжига двух праймеров. Причем степень амплификации анализируемого района генома в первом случае будет находиться в линейной зависимости от количества циклов, а во втором — в экспоненциальной. Экспоненциальное накопление в процессе ПЦР амплифицируемого фрагмента заранее известного размера позволяет визуально наблюдать его после электрофоретического фракционирования препарата ДНК и делать однозначное заключение о встройке чужеродной последовательности в заданный район хромосомной ДНК.
ВВЕДЕНИЕ НОВОГО ГЕНА В КЛЕТКУ
Ввести рекомбинантный ген в клетку можно 2 способами: используя вектора или путем прямого введения.
Требования к векторной ДНК, ее состав Вектор — молекула ДНК или РНК, состоящая из двух компонентов: векторной части (носителя) и клонируемого чужеродного гена. Задача вектора — донести выбранную ДНК в клетку-рецепиент, встроить ее в геном, позволить идентификацию трансформированных клеток, обеспечить стабильную экспрессию введенного гена. Таким образом, вектор должен быть небольшим, способным поддерживаться в клетке-хозяине (реплицироваться), многократно копироваться (ампфлицироваться), экспрессировать соответствующий ген (содержать соответствующие регуляторные последовательности), должен иметь маркерный ген, позволяющий различать гибридные клетки для эффективной селекции их; должен быть способен передаваться в клетку соответствующего организма. Регуляторные последовательности, отвечающие за стабильную экспрессию гена, будут рассмотрены позднее. Можно выделить 2 группы маркерных генов, позволяющие отличить трансформированные клетки: 1. Селективные гены, отвечающие за устойчивость к антибиотикам (канамицину, тетрациклину, неомицину и др.), гербицидам (у растений). Это могут быть гены ауксотрофности по какому-либо субстрату и т. д. Основной принцип работы такого маркера — способность трансформированных клеток расти на селективной питательной среде, с добавкой определенных веществ, ингибирующих рост и деление нетрансформированных, нормальных клеток. 2. Репортерные гены, кодирующие нейтральные для клеток белки, наличие которых в тканях может быть легко тестировано. Чаще всего в качестве репортерных используются гены β-глюкуронидазы (GUS), зеленого флюоресцентного белка (GFP), люциферазы (LUC), хлорамфениколацетилтрансферазы (CAT). К настоящему времени из этого арсенала наиболее часто используют гены GUS и GFP и, в меньшей степени, LUC и CAT. Используемый в настоящее время как репортерный ген GUS является модифицированным геном из Escherichia coli, кодирующим β-глюкуронидазу с молекулярной массой 68 кД. GUS активен в широком диапазоне условий среды с оптимумом при pH 5–8 и 37 °C. Он может гидролизовать обширный спектр природных и синтетических глюкуронидов, что позволяет подбирать соответствующие субстраты для спектрофотометрического или флюориметрического определения активности фермента, а также для гистохимического окрашивания тканей in situ (например, в синий цвет). Фермент достаточно стабилен: он устойчив к нагреванию (время полужизни при 55 °C составляет около 2 ч) и к действию детергентов. В процессе замораживания-оттаивания потери активности GUS не происходит. В составе химерных белков, созданных генно-инженерными методами, GUS обычно сохраняет свою функциональную активность. В живых клетках белок GUS также весьма стабилен и активен от нескольких часов до нескольких суток. GFP (green fluorescent protein — зеленый флюоресцентный белок, или белок зеленой флюоресценции) был обнаружен Shimomura с соавт. в 1962 г. у люминесцирующей медузы Aequorea victoria. Ген GFP был клонирован в 1992 г. Prasher и соавт., и уже через несколько лет началось активное использование этого гена как репортерного в работах с самыми разными про- и эукариотическими организмами. В настоящее время ген GFP применяется в сотнях работ во всем мире, и число их стремительно нарастает. Столь быстрый рост вызван особыми свойствами белка GFP, а именно его способностью флюоресцировать в видимой (зеленой) области спектра при облучении длинноволновым УФ. Эта флюоресценция обусловлена непосредственно белком, для ее проявления не требуется субстратов или кофакторов. Благодаря этому свойству ген GFP является очень перспективным репортерным геном, позволяющим проводить разнообразные прижизненные (недеструктивные) исследования с трансгенными организмами. Многочисленные производные GFP получили общее название AFP (autofluorescent proteins — автофлюоресцентные белки). Из морской анемоны Discosoma sp. недавно выделен еще один белок DsRed, флуоресцирующий в красном свете. Еще несколько аналогичных флюоресцирующих белков было выделено в самое последнее время учеными Российской академии наук из различных коралловых полипов порядка Anthozoa. Он может быть денатурирован очень высокой температурой, крайними значениями pH или сильными восстановителями типа Na2SO4. При возвращении к физиологическим условиям GFP в значительной степени восстанавливает способ ность к флюоресценции. В составе химерных белков, созданных генноинженерными методами, GFP обычно сохраняет свою функциональную активность. В живых клетках белок GFP также очень стабилен. CAT — гены отвечают за синтез хлорамфениколацетилтрансферазы (выделены из Escherihia coli). Этот фермент катализирует реакцию переноса ацетильной группы от ацетил-КоА к хлорамфениколу. Определяется гистохимически, по изменению окраски ткани при добавлении соответствующего субстрата. LUC — ген кодирует фермент люциферазу (клонирована из бактерий и светлячка). Она вызывает свечение трансформированных клеток. Бактериальный фермент состоит из двух субъединиц. Для определения активности ферментов необходимо специальное оборудование — флуориметр и цифровая видеокамера с амплификатором светового сигнала. Фермент теряет активность при действии детергентов и повышенной температуры. Замена селективных генов на репортерные при отборе трансгенных растений часто весьма желательна, так как возможность потенциального риска для окружающей среды и здоровья человека при использовании репортерных генов практически исключена. Однако область применения репортерных генов шире, чем просто контроль трансгеноза. Другое, и, очевидно, более важное назначение репортерных генов состоит в том, чтобы выявлять (по возможности количественно) временные и пространственные особенности экспрессии данного конкретного гена, будь то собственного или чужеродного. Присоединение репортерного гена к одной лишь промоторной области позволяет исследовать в "чистом виде" ее роль в регуляции экспрессии изучаемого гена на уровне транскрипции. Замена белок-кодирующей области гена на репортерную при сохранении участка, кодирующего 5'-концевую не транслируемую последовательность мРНК, позволяет оценить роль этой последовательности в процессах транспорта мРНК из ядра в цитоплазму и инициации трансляции. Одно из самых важных свойств гена — способность к экспрессии. За это свойство отвечают различные генетические элементы, которые мы должны встроить в векторную молекулу, несущую ген.
Регуляция экспрессии гена у прокариот Многие бактериальные гены устроены таким образом, что они способны функционировать с существенно разной эффективностью. У Е. coli, например, относительное содержание различных белков варьирует в очень широких пределах (от менее чем 0.1 % до 2 %) в зависимости от их функций; при этом каждый белок в хромосоме Е. coli кодируется единственным геном. Такие вариации обусловлены действием системы контроля генной экспрессии, которая осуществляется главным образом на уровне транскрипции ДНК. Таким образом, чаще всего уровень активности гена связан с количеством синтезируемой на нем мРНК, то есть с активностью фермента РНК-полимеразы. Последовательности ДНК, расположенные перед началом структурного гена и определяющие степень активности РНК-полимеразы, называются регуляторными последовательностями. Одна из таких последовательностей представляет собой участок ДНК, с которым связывается РНК-полимераза. Этот участок называется промотором. Последовательность оснований промотора определяет частоту инициации синтеза иРНК, причем замена одного основания в этой последовательности может привести к уменьшению частоты инициации в 1000 раз. Промотор может быть сильным и слабым. Сильный промотор инициирует синтез иРНК часто, слабый — гораздо реже. С другой стороны, промотор может быть регулируемым и нерегулируемым. Например, промотор β-лактамазы нерегулируемый, но сильный. Использование таких промоторов не всегда удобно. Дело в том, что большое количество белка может блокировать рост бактерий. Кроме того, интенсивная транскрипция рекомбинантной ДНК может помешать репликации плазмиды, и она будет утрачена. Поэтому удобнее использовать регулируемые сильные промоторы (индуцибельные), включение которых, а значит и синтез чужеродного белка можно осуществить, когда получена большая бактериальная масса. Некоторые плазмидные векторы содержат промотор, работа которого регулируется температурочувствительным белковым продуктом гена-репрессора. Белок-репрессор активен при определенных температурах и блокирует действие промотора. Повысив температуру до 42 °C, можно "включить" промотор и быстро получить большое количество требуемого белка. В качестве индуцибельных промоторов используют также Тrр-промотор триптофанового оперона, который регулируется триптофановым голоданием, lас-промотор лактазного оперона, который индуцируется субстратом (лактозой) и другие. Интенсивность транскрипции определенных структурных генов может зависеть от эффективности ее терминации, в частности, от того, как часто РНК-полимераза прекращает синтез РНК, не дойдя до этих генов. Сравнительно недавно обнаружено, что во многих оперонах E.coli, контролирующих биосинтез аминокислот, между промотором и первым структурным геном имеется терминирующая последовательность. В определенных условиях происходит образование терминирующего сигнала, ослабляющего интенсивность транскрипции. Это явление получило название аттенуации, а участок ДНК — аттенуатор (ослабитель). Как и репрессия, аттенуация зависит от присутствия в среде соответствующих аминокислот. Например, избыток триптофана в клетках триптофанзависимых мутантов, дефектных по репрессору, только 1 из 10 молекул РНК-полимеразы, начавших транскрипцию, преодолевает аттенуатор и считывает структуру генов. Удаление триптофана втрое повышает эффективность транскрипции генов. В отличие от репрессии, антенуация зависит не от самой аминокислоты, а от триптофанил — тРНК (аминокилоты, присоединенной к соответствующей тРНК). На эффективность продуктивности рекомбинантной ДНК в существенной степени влияет количество копий этой ДНК в расчете на клетку. Суммарная активность экспрессируемого гена растет с ростом копийности плазмиды. Таким образом, используя многокопийные плазмиды, можно достичь сверхсинтеза нужных белковых продуктов. Обычно используемые плазмидные векторы (pBR 322 и др.) поддерживаются в клетке в количестве 20–50 копий. Сейчас исследователи имеют в своем распоряжении температурно-чувствительные мутантные плазмиды, способные накопить до одной-двух тысяч копий на клетку, не нарушая ее жизненно-важных функций. Таким образом, можно достичь сверхпродукции плазмидных генов бактериальными штаммами-сверхпродуцентами. Регуляция экспрессии у Е. coli происходит также и на уровне трансляции. Последовательность оснований длиной 6–8 нуклеотидов, расположенная непосредственно перед инициирующим кодоном АУГ, определяет эффективность трансляции. Эта последовательность представляет собой участок связывания мРНК с рибосомой. Как правило, он отстоит на 8 нуклеотидов от инициирующего кодона, и его сдвиг в ту или иную сторону может резко снижать эффективность трансляции соответствующей мРНК. Описанный участок называется последовательностью Шайна-Дальгарно, по имени исследователей, впервые его идентифицировавших. В состав вектора кроме всего прочегодолжен входить маркерный ген, позволяющий селектировать измененные клетки. Часто в качестве селективных используют широко распространенные в природе гены ферментов, модифицирующих антибиотики и инактивирующие их действие.
Особенности организации генома эукариот У эукариотических организмов механизм регуляции транскрипции гораздо более сложен. В результате клонирования и секвенирования генов эукариот обнаружены специфические последовательности, принимающие участие в транскрипции и трансляции. Для эукариотической клетки характерно: 1. Наличие интронов и экзонов в молекуле ДНК. 2. Созревание и-РНК — вырезание интронов и сшивка экзонов. 3. Наличие регуляторных элементов, регулирующих транскрипцию, таких как: а) промоторы — 3 вида, на каждый из которых садится специфическая полимераза. Pol I реплицирует рибосомные гены, Pol II — структурные гены белков, Pol III — гены, кодирующие небольшие РНК. Промотор Pol I и Pol II находятся перед участком инициации транскрипции, промотор Pol III — в рамках структурного гена. б) модуляторы — последовательности ДНК, усиливающие уровень транскрипции; в) усилители — последовательности, усиливающие уровень транскрипции и действующие независимо от своего положения относительно кодирующей части гена и состояния начальной точки синтеза РНК; г) терминаторы — специфические последовательности, прекращающие и трансляцию, и транскрипцию. Эти последовательности по своей первичной структуре и расположению относительно инициирующего кодона отличаются от прокариотических, и бактериальная РНК-полимераза их не "узнает". Таким образом, для экспрессии эукариотических генов в клетках прокариот нужно, чтобы гены находились под контролем прокариотических регуляторных элементов. Это обстоятельство необходимо учитывать при конструировании векторов для экспрессии.
Типы векторов для введения гена в клетку Существует несколько типов векторов: Бактериальные плазмиды Основная масса клеточной ДНК бактерий содержится в хромосоме (в хромосоме Е. coli, например, 4 млн. пар нуклеотидов). Однако кроме хромосом бактерии содержат большое количество очень маленьких кольцевых молекул ДНК плазмид длиной несколько тысяч пар оснований (молекулярная масса от 1,5 до 300 мегадальтон, 1 МД = 1500 п. о). Такие мини-хромосомы называют плазмидами. Как правило, плазмиды имеют в своем составе гены устойчивости к антибиотикам, ионам тяжелых металлов (R-плазмиды), а также гены, контролирующие катаболизм некоторых органических соединений (плазмиды биодеградации, или D-плазмиды). Поскольку эти гены находятся в плазмидах, они представлены гораздо большим числом копий. Высокая копийность плазмид обеспечивает клетке синтез большого количества ферментов, химически нейтрализующих антибиотики или ксенобиотики, что и обеспечивает устойчивость к последним. Плазмиды, по-видимому, вездесущи, так как их выделяют из разных штаммов и видов бактерий, но не являются обязательными компонентами генома, а в некоторых природных штаммах плазмиды не обнаружены вообще. Поскольку плазмидная ДНК значительно меньше хромосомной, ее довольно легко выделить в чистом виде. В присутствии ионов кальция плазмиды легко поглощаются бактериями-рецепиентами, даже если те их никогда не содержали, и в клетках бактериального потомства можно обнаружить много копий поглощенной плазмиды. Однако бактериальная клетка обычно может содержать в своем составе плазмиды одного типа. Это явление несовместимости плазмид. Существуют группы несовместимости — Inc-группы (от английского incompatibility — несовместимость). В такой группе может быть несколько плазмид, совместимых между собой, но не совместимых с другими плазмидами. У этих плазмид сходны многие признаки и часто значительна гомология ДНК. Число копий плазмиды в клетке может существенно варьировать. Это зависит от генетических особенностей как клетки, так и плазмиды. Плазмиды, находящиеся "под ослабленным контролем", могут размножаться до тех пор, пока их количество не достигнет 10-200 копий на клетку. Если же плазмида находится "под строгим контролем", она реплицируется с той же скоростью, что и главная хромосома. Такие плазмиды содержатся в клетке в одной или в нескольких копиях. Естественно, что для клонирования рекомбинантных ДНК стараются использовать плазмиды первого типа. Но это не обязательно, так как плазмиды в присутствии хлорамфеникола могут умножаться независимо от деления хромосомы, и количество копий плазмиды может многократно увеличиваться. Одна их наиболее часто употребляемых плазмид для клонирования pBR 322 создана на основе плазмид природного происхождения, выделенных из Е. coli. Эта плазмида содержит гены устойчивости к двум антибиотикам: ампициллину и тетрациклину, причем в генах устойчивости к этим антибиотикам имеются сайты рестрикции. Если фрагмент чужеродной ДНК встраивается в один из генов устойчивости, то последний инактивируется. Следовательно, успешное встраивание фрагмента чужеродной ДНК в один из этих генов легко детектировать по исчезновению у бактерий устойчивости к данному антибиотику. Но при этом сохраняется устойчивость к другому антибиотику. Таким образом, вектор дает возможность детектировать только те клоны бактерий, которые содержат рекомбинантную плазмиду.
Вирусы Есть вирусы, которые не ведут к гибели клетки, но встраиваются в геном клетки-хозяина и размножаются вместе с ней, либо вызывают ее неконтролируемый рост, т. е. превращают в раковую. К таким относятся ДНК-вирусы SV-40 и вирус полиомы. Внедрение некоторых опухолевых РНК-вирусов ведет к отпочковыванию вирусных частиц от клетки без ее лизиса. К таким вирусам относятся, например, ретровирусы (вирус саркомы Рауса и СПИДа). Для бактериальных клеток в качестве вектора часто используют бактериофаги. Вирусы являются одними из главных кандидатов на роль векторов для введения чужеродной ДНК. При вирусной инфекции каждая клетка может получить большое число копий чужеродного гена. ДНК можно встраивать так, чтобы она находилась под контролем сильных вирусных промоторов, что обеспечит высокий уровень экспрессии гена, и его продукты будут более доступны для исследования. В последние годы сконструированы многочисленные "челночные" векторы и их рекомбинантные производные, способные к репликации в животной и бактериальной клетке и эффективно экспрессирующие клонируемый ген в животной клетке. Наиболее распространенные векторы состоят из плазмиды pBR322 и интактного раннего района транскрипции ДНК SV40, а нужный ген встраивается под контроль промотора поздних генов или дополнительного раннего промотора. Например, в ДНК SV40 был встроен ген р-глобина кролика, который экспрессировался в линии клеток обезьяны, зараженных рекомбинантным вирусом: в клетках синтезировались и мРНК гена глобина, и сам белок. Вирус должен быть жизнеспособным после рекомбинирования его ДНК. Легче всего вирусы вводятся в бактерии. Недостатком вирусов как векторов является их небольшая емкость. Кроме того, вирусы заражают небольшой круг хозяев. Существуют гибридные вектора, содержащие ДНК фага и плазмиды. К ним относятся, например, космиды и фазмиды. Космиды — плазмидные вектора, в которые встроен участок генома фага А, обеспечивающий возможность упаковки этой молекулы ДНК в фаговую частицу. Фаговые частицы обеспечивают хорошее проникновение гибридной ДНК в клетку (путем инъекции), после чего происходит замыкание ДНК в кольцо по липким концам и репликация ее по плазмидному типу. Фазмиды также являются гибридами между фагом и плазмидой. После встройки чужеродной ДНК могут в одних условиях развиваться как фаги, в других — как плазмиды.
Вироиды Из всех известных в настоящее время инфекционных агентов имеют ранг наиболее странных. Известно, что самые мелкие вирусы, способные к независимой репликации, имеют размеры генома, соответствующие молекулярной массе 1 М, то есть около 1500 тыс. пар оснований. Это считали минимальным количеством генетической информации, необходимой для кодирования вирусоспецифических продуктов и подавления метаболизма хозяйской клетки. Однако в 1971 году были открыты инфекционные агенты, представляют собой очень короткую цепь 1 нитевой ковалентно связанной кольцевой РНК, состоящую из 270–300 нуклеотидов (на три порядка меньше самых минимальных вирусов), не заключенную в белковую оболочку. Это необычные патогены — самые простые и самые маленькие из всех известных. Каким образом вироиды продуцируют симптомы болезни в инфицированных растениях, не известно до сих пор. Установлено, что они реплицируются ферментами клетки-хозяина, не транслируются в видоспецифичные полипептиды, интегрируются в геном клетки-хозяина. Вироиды заражают персиситентно (не происходит выздоровления). Вызывают системную инфекцию, т. е. мигрируют из сайта внедрения в другие части растений, переносятся механически или через клеточный сок, через семена, пыльцу. Вироиды также связаны с ядерными фракциями растений и могут размножаться в ядрах. При работе с вироидами получают 1-нитевую ДНК-копию РНК и достраивают комплементарную нить для получения 2-нитевой ДНК вироида. Такая 2-цепочечная ДНК встраивается в плазмиду и передается в клетки Е. coli для клонирования. Считывание гена начинается с промотора, который узнается РНК-полимеразой, отвечающей за транскрипцию ДНК в матрицу РНК. Обычно это фрагмент ДНК из 41–44 пар оснований. Ген считывается слева направо, от 5' к 3' концу гена и заканчивается в терминальной области гена. За промотором начинается стартовый сайт транскрипции, за которым следует смысловая часть гена. Промоторная область гена содержит определенные короткие сочетания нуклеотидов, характерные для бактериальных генов, или для генов высших организмов. Такие сочетания служат сигналами для РНК-полимеразы, которая присоединяется к промоторной части гена и начинает его считывать. Однонитевые и двунитевые ДНК способны инициировать репликацию вироида в механически инокулированных растениях табака. Энзиматически in vitro синтезированы также РНК вироидов, высокоинфекционные для растений. Векторные системы могут быть разработаны на основе самих РНК, на основе вироидоспецифичных ДНК, а также в комбинации вироидоспецифичных ДНК с Ti-плазмидами. Вироиды инфицируют своих хозяев в течение всего их жизненного цикла, поэтому в случае использования вироидных векторных систем можно ожидать постоянной экспрессии чужеродного гена в растении.
Плазмиды агробактерий В качестве векторов могут использоваться опухолеобразующие плазмиды бактерий. Виды Agrobacterium эволюционно родственны клубеньковым бактериям, относящимся к роду Rhizobium, и имеют много общих с ними черт. Однако характер взаимодействия агробактерий с растением имеет своеобразные особенности. Взаимодействие видов Agrobacterium с растениями представляет особый интерес, так как при этом виде паразитизма один из партнеров специфически видоизменяет свойства хозяина, встраивая свои гены в его геном. Кроме того, это служит уникальным примером миграции ДНК прокариот в эукариотическую клетку. ДНК митохондрий и хлоропластов Хлоропласты и митохондрии содержат полноценную генетическую систему, то есть все компоненты, необходимые для экспрессии генетической информации: ДНК, ДНК-полимеразы, РНК-полимеразы и белоксинтезирующий аппарат (рибосомы, т-РНК, аминоацил-тРНК-синтетаЗы). Хлоропластная и митохондриальная ДНК также привлекают внимание ученых в качестве возможных векторов для переноса генов в клетку. Структурная организация этих клеточных субгеномов существенно различается. Хлоропласты и другие пластиды обладают одинаковой генетической информацией, так называемым пластомом. У высших растений он представляет собой замкнутую молекулу ДНК длиной 150 т. н.п., достаточную для кодирования примерно 100 белков. Для синтеза пластид необходимо значительно больше белков. Остальные белки кодируются ядром, синтезируются в цитоплазме и поступают в хлоропласты. Некоторые важнейшие белки хлоропластов состоят из нескольких субъединиц, часть из них синтезируется на рибосомах цитоплазмы и транспортируется в хлоропласт, где они объединяются с другими полипептидами, закодированными в самом хлоропласте и там же синтезируемыми. Таким образом, для биосинтеза функционально активного хлоропласта требуется согласованная экспрессия генома и пластома. Различные типы пластид содержат неодинаковые количества идентичных копий пластома: от 10–20 копий в пластидах корней и зрелых хлоропластах до сотен копий в молодых хлоропластах картофеля. Такой уровень амплификации позволяет надеяться на надежную экспрессию чужеродной ДНК при использовании их в качестве векторов в генно-инженерных экспериментах. Кроме того, гены рибосомальной РНК пластид и большой субъединицы РБФК кодируются геномом хлоропластов. Возможно, введение сильных промоторов в эти гены и дополнительная их модификация существенно повлияют на фотосинтетическую активность растительной ткани. Гены растений также способны к экспрессии в клетках Е. coli. Это гены большой субъединцы РБФК. Преимущество хлоропластных генов заключается в том, что их экспрессия к клетках кишечной палочки может быть достигнута путем простого объединения транскрибируемых последовательностей, т. к. в ДНК хлоропластов и бактерий до начала стартовых кодонов трансляции расположена одинаковая нуклеотидная последовательность. Это дает возможность синтезировать растительные экономически важные полипептиды с помощью клеток прокариот. В отличие от хлоропластной, ДНК митохондрий характеризуются исключительным разнообразием, и их величина колеблется от 200 до 2400 т. н.п. Однако никакой корреляции между размером митохондриального генома и числом белковых продуктов, синтезируемых изолированными митохондриями, не наблюдается. Это явление, а также большие размеры митохондриальной ДНК, по-видимому, можно объяснить присутствием ДНК, бесполезной для функционирования митохондрий. В составе митохондриальной ДНК имеются структурные гены, кодирующие полипептиды, гены рибосомных и транспортных РНК. Однако большая часть белков митохондрий, как и хлоропластов, кодируется ядерными генами. Но если геном хлоропластов представлен гомогенной популяцией крупных кольцевых молекул, то в митохондриях содержится несколько классов кольцевых молекул, не все функции которых еще ясны. Митохондриальный геном животных организмов намного меньше, 15–19 т. н.п., и более консервативен по структуре. Гены митохондрий кодируют 2 группы признаков — работу дыхательных систем и устойчивость к антибиотикам и другим ядам. В митохондриальном геноме растений есть также гены, отвечающие за признак мужской стерильности цитоплазмы.
Транспозоны Транспозоны — сегменты ДНК, которые контролируют собственную транспозицию (перемещение) из одного сайта ДНК в другой путем вырезания из исходного сайта и внедрения в новый сайт хромосомы или плазмиды. Впервые были открыты в 40-х годах американской ученой Барбарой Мак-Клинток у кукурузы. Эти гены, индентифицированные по их способности подавлять экспрессию других генов кукурузы, находящихся рядом с ними, не имели фиксированного положения в хромосоме. Они как бы передвигались по всему геному растения. Регуляторные элементы могли встраиваться и выщепляться, причем после их выщепления зачастую начинали функционировать ранее молчащие гены. Оказалось, что гены, ассоциированные с регуляторными элементами, становились нестабильными и часто мутировали из-за нестабильности самих этих элементов. В течение многих лет кукуруза оставалась единственной системой, в которой обнаруживались такие подвижные генетические элементы. Сейчас — и у бактерий, дрозофил и других организмов. Механизм перемещения фрагментов ДНК по геному до конца не выяснен. ДНК переносится ферментом транспозазой. Фермент кодируется последовательность длиной около 20 нуклеотидов в середине транспозона. Он специфически взаимодействует с концевыми инвертированными повторами мобильного элемента и может вырезать его из хромосомы. Вырезание может происходить точно — с восстановлением исходной структуры участка ДНК, и неточно, то есть с делециями и вставками от одного до нескольких нуклеотидов. Это приводит к появлению стабильных мутаций и является одним из механизмов создания новых последовательностей ДНК. Как правило, мобильные генетические элементы многократно повторены в геноме и образуют гетерогенные семейства, члены которых диспергированы по хромосомам. Большая часть членов каждого семейства являются дефектными копиями и не кодируют какой-либо функции, хотя сохраняют способность к перемещению. Поведение транспозонов можно расценить как паразитическое. Длина их от 2 до 10 тысяч нуклеотидных пар. У высших эукариот на долю транспозонов приходится примерно 10 % ДНК клетки. Большинство их перемещается изредка, но, так как их в клетке довольно много, транспозиция оказывает значительное влияние на разнообразие видов. Биологический смысл перемещения отдельных сегментов ДНК: — прерывание соответствующего гена, что ведет к эволюции; — регуляция деятельности генов, так как транспозоны могут нести сигналы для начала считывания генов. В новых областях усиливают или запрещают работу гена. Транспозоны также участвуют в горизонтальном переносе генов. У бактерий были обнаружены 2 класса подвижных генов, различающихся по длине и сложности организации. 1. Инсерционные последовательности, или 1S элементы, имеющие длину около тысячи пар нуклеотидов и содержащие только ген, отвечающий за их перемещение. 2. Транспозоны, длиной от 3 до 20 т. н. п., состоящие из ряда дополнительных генов, отвечающих за устойчивость бактерий к различным токсическим веществам. Поскольку подвижные гены могут перемещаться в пределах генома с одного места на другое, то они могут быть весьма эффективными векторами для передачи рекомбинантной ДНК. Генетическая трансформация с помощью векторов на основе транспозонов была впервые осуществлена на дрозофиле. С помощью транспозируемого элемента Р дрозофиле был передан ген, обуславливающий коричневую окраску глаз. Перенос генов при помощи транспозонов имеет большие преимущества, так как он происходит с высокой частотой и не влечет значительных перестроек интегрируемой ДНК. Кроме того, этим методом можно переносить достаточно большие фрагменты ДНК.
Способы прямого введения гена в клетку Прямое введение гена в клетку осуществляют несколькими способами: — Трансфекция — Микроинъекция — Электропорация — Метод «мини-клеток» — Упаковка в липосомы — Электронная пушка При трансфекции ДНК адсорбируется на кристаллах фосфата кальция (Грэхем Ван дер Эб, 1973). Образуются частицы кальциевого преципитата. Они поглощаются клеткой путем фагоцитоза. Для повышения эффективности трансформации к специфической ДНК, содержащей ген, по которому будет производится селекция, добавляется неспецифическая ДНК-носитель. Обычно для этой цели берут ДНК из тимуса теленка или спермы лосося. Часть ДНК связывается с мембраной и не попадает в клетки. ДНК акцептируют от 15 до 90 % клеток. Через несколько суток после введения небольшая доля клеток способны экспрессировать чужеродные гены, но затем уровень экспрессии падает и более или менее стабильную трансформацию претерпевает 10-3-10-5 клеток. Для трансфекции используется и ДЭАЭ-декстран, полимер, адсорбирующий ДНК. Эффект вхождения в клетки и время экспрессии высоки, но частота стабильной трансформации ниже, чем при использовании преципитата кальция. Частоту трансфекции увеличивает глицериновый шок (4 минуты в 15 % растворе глицерина в НЕРЕS-буфере). В клетки можно вводить любой ген, если заранее лигировать его с клонированным селективным маркером. Однако дальнейшие исследования показали, что лигирование вне клетки не обязательно. Клетки, поглощающие селективный ген, вместе с ним поглощают и другую ДНК, имеющуюся в кальциевом преципитате. Таким образом, пользуясь методом котрансформации, практически любой клонированный сегмент ДНК можно ввести в культивируемые клетки эукариот, если включить эту ДНК вместе с селективным маркером в состав смеси для образования кальциевого преципитата. Для трансфекции можно использовать хромосомы или фрагменты хромосом. Клетки-доноры блокируются на стадии митоза. Митотические хромосомы высвобождаются под воздействием осмотического шока и гомогенизации. Их очищают путем дифференциального центрифугирования. Хромосомы осаждают на поверхности клеток хлористым кальцием, а через несколько часов обрабатывают реагентом, способным перфорировать мембраны (например, глицерином). Для обработки клеток-рецепиентов используются грубо очищенные препараты хромосом, так как хромосомы при этом разрушаются меньше всего. Количество хромосом для обработки 1 клетки ограничено. Лучше использовать не более 20 хромосом на 1 клетку-рецепиент, так как при высоких концентрациях хромосом в суспензии они агглютинируют. Рецепиентная клетка содержит фрагменты донорных хромосом, которые могут встраиваться в геном, могут реплицироваться самостоятельно. Во введенных фрагментах часто наблюдаются делеции. Не все клетки способны к трансформации геномной ДНК с высокой частотой. Человеческие фибробласты эффективно включают плазмидную ДНК и почти не включают геномную. Микроинъекция ДНК в клетки млекопитающих стала возможной с появлением прибора для изготовления микропипеток диаметром 0.1–0.5 микрона и микроманипулятора (рис. 45). Так, плазмиды, содержащие фрагмент вируса герпеса с геном тимидинкиназы (ТК) и плазмиду pBR322, были инъецированы в ТК-клетки и было показано, что ТК-ген проник в ядра и нормально в них реплицировался. Метод введения ДНК с помощью микроинъекций был разработан в начале 70-х годов Андерсоном и Диакумакосом. В принципе, при наличии хорошего оборудования можно за 1 час инъецировать 500-1000 клеток, причем в лучших экспериментах в 50 % клеток наблюдается стабильная интеграция и экспрессия инъецированных генов. Преимущество описываемого метода заключается также в том, что он позволяет вводить любую ДНК в любые клетки, и для сохранения в клетках введенного гена не требуется никакого селективного давления.
Рис. 45. Введение ДНК путем микроинъекции
Электропорация основана на том, что импульсы высокого напряжения обратимо увеличивают проницаемость биомембран. В среду для электропорации добавляют клетки и фрагменты ДНК, которые необходимо ввести в клетки (рис. 46). Через среду пропускают высоковольтные импульсы (напряжение 200–350 В, длительность импульса 54 мс), приводящие к образованию пор (электропробой) в цитоплазматической мембране, время существования и размер которых достаточны, чтобы такие макромолекулы, как ДНК, могли из внешней среды войти в клетку в результате действия осмотических сил. При этом объем клетки увеличивается.

Рис. 46. Метод электропорации
Напряженность электрического поля и продолжительность его действия, концентрации трансформирующей ДНК и реципиентных клеток для каждой системы клеток подбирают экспериментально с тем, чтобы достичь высокого процента поглощения ДНК выжившими клетками. Показано, что в оптимальных условиях электропорации количество трансформантов может достигать 80 % выживших клеток. Электропорация — физический, а не биохимический метод, и это, по-видимому, обусловливает его широкое применение. Многочисленные исследования продемонстрировали, что электропорация может успешно использоваться для введения молекул ДНК в разные типы клеток, такие как культивируемые клетки животных, простейшие, дрожжи, бактерии и протопласты растений. Электропорирующий эффект высоковольтного разряда на бислойную липидную мембрану, по-видимому, зависит от радиуса ее кривизны. Поэтому мелкие бактериальные клетки эффективно поглощают ДНК при значительно большей напряженности (10 кВ/см и более), чем крупные животные и растительные клетки, эффективно поглощающие ДНК при напряженности поля 1–2 кВ/см. Электропорация — наиболее простой, эффективный и воспроизводимый метод введения молекул ДНК в клетки. Однако до недавнего времени этот метод использовался в ограниченном числе лабораторий в связи с отсутствием серийных приборов — электропораторов. Появление и совершенствование таких приборов в ближайшие годы приведет к широкому применению данного подхода в генетической инженерии самых разных типов клеток. «Мини-клетки» получают путем блокирования донорных клеток митозе колцемидом. При продолжительной обработке клеток колцемидом в них вокруг каждой хромосомы формируется новая ядерная мембрана. Обработка цитохалазином В и центрифугирование приводит к образованию мини-клеток, представляющих микроядра, инкапсулированные в цитоплазматическую мембрану. Полученные мини-клетки очень чувствительны к разного рода воздействиям, поэтому для слияния подбирают специальные мягкие условия. Метод трудный, капризный, эффективность низкая — 10-6-10-7. Упаковка в липосомы используется для защиты экзогенного генетического материала от разрушающего действия рестриктаз. Липосомы — сферические оболочки, состоящие из фосфолипидов. Получают их путем резкого встряхивания смеси водного раствора и липидов, либо обрабатывая ультразвуком водные эмульсии фосфолипидов. Липосомы, состоящие из фосфатидилсерина и холестерина наиболее пригодны для введения ДНК в клетки животных и растений. Системы переноса с помощью липосом низкотоксичны по отношению к клеткам. Метод биологической баллистики (биолистики) является одним из самых эффективных на сегодняшний день методов трансформации растений, особенно однодоль- Суть метода заключается в том, что на мельчайшие частички вольфрама, диаметром 0,6–1,2 мкм, напыляется ДНК вектора, содержащего необходимую для трансформирования генную конструкцию. Вольфрамовые частички, несущие ДНК, наносятся на целлофановую подложку и помещаются внутрь биолистической пушки. Каллус или суспензия клеток наносится в чашку Петри с агаризированной средой и помещается под биолистическую пушку на расстоянии 10–15 см. В пушке вакуумным насосом уменьшается давление до 0,1 атм. В момент сбрасывания давления вольфрамовые частички с огромной скоростью выбрасываются из биолистической пушки и, разрывая клеточные стенки, входят в цитоплазму и ядро клеток. Обычно клетки, располагающиеся непосредственно по центру, погибают из-за огромного количества и давления вольфрамовых частиц, в то время как в зоне 0,6–1 см от центра находятся наиболее удачно протрансформированные клетки. Далее клетки осторожно переносят на среду для дальнейшего культивирования и регенерации. С помощью биолистической пушки были протрансформированы однодольные растения, такие, как кукуруза, рис, пшеница, ячмень. При этом были получены стабильные растения-трансформанты. Кроме успехов в получении трансгенных однодольных, биолистическая трансформация применяется для прямого переноса ДНК в эмбриогенную пыльцу и дальнейшего быстрого получения трансгенных дигаплоидных растений, которые являются важным этапом в селекционной работе. В настоящее время этим методом была проведена трансформация растений табака, и после регенерации гаплоидных растений, получены стабильные трансформанты.
Генетические манипуляции с бактериальными клетками В настоящее время бактерия Е. coli является самой изученной клеткой из всех существующих. У большинства наиболее полно изученных фагов клеткой — хозяином является также Е. coli. Протопласт Е. coli одет в муреиновый мешок, прилегающий к внешней мембране. Е. coli относится к микроорганизмам, не обладающим физиологической компетентностью к поглощению экзогенной ДНК. Поэтому необходимо создать условия, позволяющие преодолеть барьер клеточной стенки. Сначала получают сферопласты путем обработки клеток лизоцимом в изотоническом растворе. Липополисахаридный слой внешней мембраны грамотрицательной бактерии стабилизирован двухвалентными катионами, поэтому для разрыхления внешней мембраны Е. coli используется комплексообразователь этилендиаминтетрауксусная кислота (EDTA), которая связывает двухвалентные катионы. При обработке EDTA часть липополисахаридов высвобождается из внешней мембраны клетки, и лизоцим может достигнуть муреинового мешка и гидролизовать его. Это ведет к повышению проницаемости клеточной оболочки. Усовершенствование методов получения сферопластов Е. coli и их трансфекции позволили достичь достаточно высокой эффективности трансформации молекулами ДНК различных фагов. Обнаружено, что на инфекционность существенное влияние оказывает форма молекул фаговых ДНК, которую они принимают in vivo. Фаги с кольцевой или линейной, но быстро замыкающейся ДНК (лямбоидные фаги) характеризуются наибольшей эффективностью трансфекции. Успешное проведение экспериментов на кишечной палочке стало стимулом для проведения аналогичных исследований с другими прокариотическими организмами. Наибольших успехов удалось достичь с клетками Bacillus subtilis. В. subtilis — непатогенный почвенным микроорганизм, растущий в строго аэробных условиях. Бациллы не образуют токсинов и непатогенны ни для животных, ни для человека, тогда как клеточная стенка Е. coli содержит эндотоксин, который довольно трудно отделить от продуктов генной инженерии. Кроме того, клеточная стенка бацилл имеет простую структуру и бактерии могут секретировать многие белки в культуральную жидкость. 20 различных видов бацил секретируют в культуральную жидкость более 40 ферментов с внеклеточной локализацией. Е. coli секретирует в среду относительно мало белков, а выделение и очистка их затруднены. В бациллах также обнаружены плазмиды и фаги, которые к настоящему моменту уже хорошо изучены. Чужеродные гены клонируют в так называемых челночных векторах. Эти вектора с одинаковым успехом реплицируются в клетках нескольких хозяев, в данном случае, в клетках Е. coli и В. subtilis. Векторы были получены комбинацией in vitro фрагментов этих плазмид. Гены Е. coli со своими регуляторными районами не функционируют в В. subtilis, поэтому были использованы собственные гомологичные районы В. subtilis. Для конструирования рекомбинантной ДНК, содержащей в своем составе ген, который должен экспрессироваться, придерживаются следующей стратегии. Синтезируют кДНК или из клонотеки выделяют клетки, несущие фрагмент генома с нужным геном, и клонируют их в соответствующем векторе. Фрагменты геномной ДНК подвергают модификации — удаляют из них некодирующие области и участки соседних генов. Часто для проведения этой операции необходимо секвенирование данного фрагмента ДНК. Затем конструируются промежуточные рекомбинантные ДНК, в которых ген помещается под контроль бактериальных регуляторных элементов (промотор, оператор, точка связывания с рибосомами). Эти регуляторные элементы выделяют из гибридных плазмид, сконструированных специально как источники регуляторных элементов. Полученная конструкция встраивается в подходящий вектор, например, pBR 322, и ген экспрессируется в бактериальной клетке. Однако удобнее встраивать ген в специальный вектор для экспрессии, который уже содержит регуляторные элементы, обеспечивающие активную экспрессию после введения рекомбинантной плазмиды в бактериальную клетку. К таким эффективным регуляторным участкам относится, например, сильный промотор гена бэта-лактамазы (ген устойчивости к пенициллину, входящий в состав плазмиды pBR 322). Ряд генов, в том числе и ген инсулина, встраивали в сайт рестрикции PstI, который расположен в структурной части гена. Промотор этого гена обеспечи вает эффективную транскрипцию, которая продолжается до тех пор, пока РНК-полимераза не дойдет до сигнала терминации встроенного гена. В качестве примера маркирования вектора могут служит первые эксперименты с Е. coli, а точнее с одной из ее плазмид pBR322, проведенные Гилбертом для получения инсулина. Плазмида pBR322 содержит 2 гена, которые определяют устойчивость к ампициллину и тетрациклину. Рестриктаза PstI расщепляет плазмиду в средней части гена, кодирующего фермент устойчивости к апициллину. После расщепления плазмиды на ее концы с помощью концевой трансферазы надстраивали последовательность из четырех нуклеотидов с остатками гуанина. Затем, как обычно, с помощью лигаз "вшивали" ген проинсулина, получая рекомбинантную ДНК. Встроенный в плазмиду фрагмент ДНК нарушал синтез фермента, разрушающего ампициллин, но ген, обеспечивающий устойчивость к тетрациклину, оставался активным. Трансформированные таким образом клетки Е. coli синтезировали гибридный белок, содержащий последовательности пенициллазы и проинсулина, поэтому биологически активный инсулин получали путем отщепления пенициллазы и средний сегмент проинсулина. С другой стороны, если фрагмент чужеродной ДНК встраивается в один из генов устойчивости, то последний инактивируется. Следовательно, успешное встраивание фрагмента чужеродной ДНК в один из этих генов легко детектировать по исчезновению у бактерий устойчивости к данному антибиотику.
ВВЕДЕНИЕ ГЕНОВ В КЛЕТКИ МЛЕКОПИТАЮЩИХ
Манипуляции с клетками млекопитающих можно разделить на 2 большие группы: эксперименты с соматическим клетками и эксперименты по трансформации половых клеток. В последнем случае конечный результат — получение трансгенных организмом.
Характеристика векторов для переноса генов в животные клетки Одними из лучших носителей для введения чужеродной информации в животную клетку являются вектора на основе ретровирусов, например, на основе вируса лейкоза мышей. Они обеспечивают высокоэффективный перенос генов и их стабильное встраивание в хромосому клеток-мишеней. В основном трансформации животных клеток осуществляют либо с помощью ретровирусов (около 40 % от всех трансформаций), либо путем упаковки ДНК в липосомы (25 %), реже используют аденовирусы, так как они могут вызывать сильный иммунный ответ, кроме того, невозможно их повторное введение. Если проблема доставки чужеродной ДНК in vitro практически решена, а ее доставка в клетки-мишени разных тканей in vivo успешно решается (главным образом путем создания конструкций, несущих рецепторные белки, в том числе и антигены, специфичные для тех или иных тканей), то другие характеристики существующих векторных систем — стабильность интеграции, регулируемая экспрессия, безопасность — все еще нуждаются в серьезных доработках. Прежде всего, это касается стабильности интеграции. До настоящего времени интеграция в геном достигалась только при использовании ретровирусных либо аденоассоциированных векторов. Повысить эффективность стабильной интеграции можно путем совершенствования генных конструкций типа рецептор-опосредованных систем, либо путем создания достаточно стабильных эписомных векторов (то есть ДНК-структур, способных к длительной персистенции внутри ядер). В последнее время особое внимание уделяется созданию векторов на базе искусственных хромосом млекопитающих (MAC — mammalian artificial chromosomes). Благодаря наличию основных структурных элементов обычных хромосом такие минихромосомы длительно удерживаются в клетках и способны нести полноразмерные (геномные) гены и их естественные регуляторные элементы, которые необходимы для правильной работы гена, в нужной ткани и в должное время. Такие искусственные хромосомы уже созданы для дрожжей (YAK), так как геном дрожжей полностью картирован. Для идентификации модифицированных клеток, необходимы маркеры. Если трансформируют соматические клетки, то применяют обычно селективные маркеры. Аксель с коллегами из колледжа терапии и хирургии Колумбийского университета исправили, таким образом, генетический дефект клеток мыши. Они взяли фрагмент ДНК, содержащий ген тимидинкиназы (ТК), который получен из вируса герпеса, смешали эту ДНК с несколькими миллиграммами ДНК-носителя из спермы лосося и осадили ДНК на культуру L-клеток мыши, в которых ген ТК отсутствовал (ТК-). С частотой 1 на 100000 клетки приобретали ген ТК, поэтому на селективной среде, которая не позволяла расти ТК-клеткам, росли и нормально размножались ТК+ — клетки. Другой селективный маркер — ген, кодирующий дигидрофолатредуктазу (ДГФР), можно использовать при трансформации немутантных линий клетки. Благодаря экспрессии многих копий этого гена животная клетка вместе с плазмидой приобретает устойчивость к высоким концентрациям ингибитора фермента, и таким образом трансформантов можно отбирать при высоких концентрациях ингибитора. Разработано еще два универсальных вектора, содержащих генные маркеры, работающие в нормальных клетках. Они построены по одному и тому же принципу: прокариотические гены, определяющие фенотип трансфецированных клеток, соединены с эукариотическими регуляторными сигналами. Один из векторов состоит из прокариотического гена устойчивости к антибиотику неомицину, встроенного в раннюю область генома SV-40. Эукариотические клетки чувствительны к аналогу неомицина G418, который инактивируется продуктом гена. Таким образом, трансфецированные клетки приобретают способность расти на среде, содержащей G418.
Генетическая транформация соматических клеток млекопитающих Культуры трансформированных клеток млекопитающих используют для получения различных веществ. Хотя культуры клеток животных, особенно при массовом выращивании, гораздо менее экономичны, чем бактериальные или дрожжевые культуры, они обладают существенным преимуществом — способностью осуществлять мелкие, но весьма важные модификации белков — продуктов гена млекопитающих. Например, для эффективного функционирования ряда белков необходимо присоединение к ним цепочек из молекул углеводов или липидов. Образование и присоединение таких цепочек — обычный процесс для клеток млекопитающих, тогда как бактериальная клетка не способна производить подобные модификации. Помимо создания клеток-продуцентов, трансформация соматических клеток млекопитающих позволяет изучать тонкие механизмы регуляции экспрессии генов и целенаправленно модифицировать генетический аппарат клетки животных, а при необходимости и человека, что имеет огромное значение для медицинской генетики. Культуры клеток млекопитающих могут оказаться эффективным источником выделения некоторых вирусных антигенов с целью получения вакцин для животных и человека. Получение таких вакцинных культур клеток осуществимо при помощи техники рекомбинантных ДНК и эффективных векторов экспрессии для клеток млекопитающих и человека. При использовании ДНК-вакцин в организм вводится не антиген, а ген, кодирующий синтез этого антигена. Ген встраивается в плазмиду, а плазмида вводится организм путем обыкновенной инъекции. ДНК-вакцины имеют хорошие перспективы в животноводстве. Фибер — белок вирусной оболочки. Эпитоп фибера кодирует синтез протективных антител. Одно из заболеваний птиц — синдром снижения яйценоскости (ССЯ) вызывается вирусом. После анализа ДНК этого вируса был выделен ген, кодирующий фибер, проклонирован и встроен в плазмиду. Рекомбинантная вакцина при введении ее в организм принесет ДНК фибера в клетку, выработка вирусного белка спровоцирует синтез специфических антител, т. е. вызовет иммунный ответ. Достоинством таких вакцин является очень маленький объем — для иммунизации одной мыши достаточно 10–50 мкг плазмиды, одной коровы — 200–300 мкг. Плазмида сохраняется в организме до 1 года. В стадии клинических испытаний в настоящее время находятся ДНК-вакцины против микоплазм, возбудителя туберкулеза, сальмонеллеза, лейшманиоза. Развитие злокачественной опухоли в организме обычно подавляет иммунитет. Проблема в том, чтобы подхлестнуть иммунную систему в целом и направить ее действие против раковых клеток. Исследователи из Медицинской школы в Энн-Арборе (Мичиган) придумали метод борьбы с раком. В опухолевые клетки толстой кишки подопытных мышей ввели гены, кодирующие белки другой линии мышей. Это можно осуществить с помощью липосом или вируса. После появления на внешней стороне клеточной мембраны этих белков иммунная система атаковала такие клетки. 20 % больных мышей выздоровели, у 70 % опухоль уменьшилась, в контрольной группе все умерли. Лимфоциты боролись не только с «меченными» клетками опухоли, но и клетками метастаз, следовательно, иммунная система «проснулась». В настоящее время ведутся эксперименты на людях с раком кожи.
Генотерапия Лечение заболеваний с помощью генов получило название генотерапии. Сейчас в мире насчитывается порядка 400 проектов, посвященных лечению с помощью генотеропии. Разработке программы генной терапии предшествуют тщательный анализ тканеспецифической экспрессии соответствующего гена, идентификация первичного биохимического дефекта, исследование структуры, функции и внутриклеточного распределения его белкового продукта, а также биохимический анализ патологического процесса. Все эти данные учитываются при составлении соответствующего медицинского протокола. Апробацию процедуры генокоррекции наследственного заболевания проводят на первичных культурах клеток больного, в которых в норме функционально активен данный ген. На этих клеточных моделях оценивают эффективность выбранной системы переноса экзогенной ДНК, определяют экспрессию вводимой генетической конструкции, анализируют ее взаимодействие с геномом клетки, отрабатывают способы коррекции на биохимическом уровне. Используя культуры клеток, можно разработать систему адресной доставки рекомбинантных ДНК, однако проверка надежности работы этой системы может быть осуществлена только на уровне целого организма. Поэтому такое внимание в программах по генной терапии уделяется экспериментам in vivo на естественных или искусственно полученных моделях соответствующих наследственных болезней у животных. Успешная коррекция генетических дефектов у таких животных и отсутствие нежелательных побочныхэффектов генной терапии являются важнейшей предпосылкой для разрешения клинических испытаний. Таким образом, стандартная схема генокоррекции наследственного дефекта включает серию последовательных этапов. Она начинается созданием полноценно работающей (экспрессирующейся) генетической конструкции, содержащей смысловую (кодирующую белок) и регуляторную части гена. На следующем этапе решается проблема вектора, обеспечивающего эффективную, а по возможности и адресную доставку гена в клетки-мишени. Затем проводится трансфекция (перенос полученной конструкции) в клетки-мишени, оценивается эффективность трансфекции, степень коррегируемости первичного биохимического дефекта в условиях клеточных культур (in vitro) и, что особенно важно, in vivo на животных — биологических моделях. Только после этого можно приступать к программе клинических испытаний. Существует два типа генотерапии: заместительная и корректирующая. Заместительная генотерапия заключается во вводе в клетку неповрежденного гена. Внесенная копия заменит по функциям сохранившийся в геноме больного дефектный ген. Все проводимые сегодня клинические испытания используют внесение в клетку дополнительных количеств ДНК. При корректирующей терапии предполагается замена дефектного гена нормальным в результате рекомбинации. Пока этот метод на стадии лабораторных испытаний, так как эффективность его еще очень низка. В зависимости от способа введения экзогенных ДНК в геном пациента генная терапия может проводиться либо в культуре клеток (ex vivo), либо непосредственно в организме (in vivo). Клеточная генная терапия или терапия ex vivo предполагает выделение и культивирование специфических типов клеток пациента, введение в них чужеродных генов, отбор трансфецированных клеток и реинфузию их тому же пациенту. Примером может служить лечение комбинированного иммунодефииицита. Комбинированный иммунодефицит может быть результатом дефекта гена аденозиндезаминазы. Впервые попытка лечения такого больного методами генотерапии была предпринята в США в 1990 г. У больного ребенка извлекли Т-лимфоциты, трансформировали ретровирусным вектором, введя нормальный ген аденозиндезаминазы и вернули клетки в организм. Введение приходится повторять. Более эффективна аналогичная трансформация стволовых клеток костного мозга. Генная терапия in vivo основана на прямом введении клонированных и определенным образом упакованных последовательностей ДНК в специфические ткани больного. В настоящее время не существует общедоступного метода культивирования клеток легких, поэтому при легочных заболеваниях единственный способ доставить чужеродный ген — это ввести его прямо в организм. Муковисцидоз — весьма распространенное среди людей белой расы тяжелое наследственное заболевание легких, которое поражает, например, в семьях из Центральной Европы одного новорожденного из 2500 и для которого установлен дефектный ген, кодирующий белок-регулятор трансмембранной проводимости. Основное проявление дефектного гена — пневмония. Поражаются все эпителиальные клетки. Основная проблема — как доставить ген в клетки, покрытые слизью, которая препятствует трансформации. Неповрежденную копию "гена заболевания", включенную в аденовирусный вектор или липосому, вводят в форме аэрозоля в дыхательные пути больного. Для коррекции нарушения при прогрессирующей мышечной дистрофии Дюшенна (заболевании мальчиков, связанном с дефектами Х-хромосомы) нормальный ген, кодирующий белок дистрофии, пытались прямо вкалывать в мышечные волокна, используя либо "голую" ДНК, либо аденовирусный вектор. Другие исследователи трансплантировали больному миобласты после генетической коррекции. Ранее неподвижный ребенок приобретал способность двигаться! К сожалению, во всех этих опытах удается получить только временный терапевтический эффект, и процедура введения гена должна неоднократно повторяться. Список наследственных заболеваний, которые пытаются или планируют лечить генами, велик. Это и ревматоидный артрит, и фенилкетонурия, и заболевания, связанные с недостатком гормонов (инсулина, эритропоэтина, гормона роста). В случае хронической анемии, связанной с дефицитом эритропоэтина, на основании опытов на животных предлагается принципиально новый подход к лечению. Так как каждая из наших клеток содержит один и тот же геном, можно заставить фибробласты кожи, которые в норме не производят эритропоэтина, синтезировать этот гормон. Для этого нужно ввести в геном новую контролирующую область и тем самым снять запрет со считывания (экспрессии) гена эритропоэтина, присутствующего, но "молчащего" в фибробластах. Практически в любой области медицины либо начаты клинические испытания лечения наследственных заболеваний с помощью генотерапии, либо в опытах на животных разрабатываются подходы к такому лечению. По мере усовершенствования методов доставки генов и контроля их экспрессии список заболеваний, к которым можно применять генотерапию, будет безусловно расширяться. Генотерапия применима не только к наследственным заболеваниям. Предстоит решить проблему лечения генами "чумы XX века" — синдрома приобретенного иммунодефицита (СПИД), возникающего при заражении вирусом иммунодефицита человека (ВИЧ). ВИЧ представляет собой ретровирус, поражающий Т-лимфоциты и макрофаги. Болезнь удалось бы победить, если бы были найдены новые гены, введение которых в зараженные ВИЧ лимфоциты останавливало бы дальнейшее размножение вируса. Предложено множество хитроумных способов борьбы со СПИДом с помощью привнесенных генов. Все они основаны на новейших данных о строении и функционировании генома ретровируса. Например, вводя прямо в мышцы больного ретровирусные векторы, несущие отдельные гены ВИЧ, ученые рассчитывали на то, что гены ВИЧ после внедрения в ДНК хромосом хозяина смогут дать информацию для синтеза вирусных белков и произойдет "противоСПИДная" иммунизация больного этими белками. Однако еще не получено ощутимых результатов, которые сулили бы успех в борьбе с вирусом дикого типа, коварство которого заключается в его изменчивости. Огромные перспективы открывает использование генотерапии для лечения онкологических заболеваний. Многолетние усилия ученых привели к пониманию того, что рак — это генетическое заболевание и его развитие происходит многостадийно, в результате серии генетических нарушений, накапливающихся в клетке. Следовательно, каждый из таких отдельных генетических эффектов может стать точкой приложения генотерапевтического подхода.
Получение трансгенных животных Если вводить ДНК в клетки многоклеточного организма, то результатом трансформации будет изменение свойств лишь небольшого числа клеток, которые приобрели новый ген или гены. Следовательно, для изменения свойств всего организма следует изменять геном половых клеток, которые перенесут новые свойства потомкам. У растений и животных целесообразно изменять такие свойства, как скорость роста, устойчивость к заболеваниям, способность адаптироваться к новым внешним условиям. В качестве маркеров в этом случае можно использовать полиморфизм длины рестрикционных фрагментов (AFLP), анализ мини-сателлитов, анализ микросателлитной ДНК (SSR), гибридизацию и т. д. Разработаны способы введения генов в эмбриональные клетки млекопитающих, мух и некоторых растений. От работы с довольно крупными яйцами амфибий перешли к изучению яйцеклеток и эмбрионов мыши, которая представляет наиболее изученное в генетическом отношении млекопитающее. Микроинъекцию клонированных генов производят в один или оба пронуклеуса только что оплодотворенной яйцеклетки мыши. Чаще выбирают мужской пронуклеус, привнесенный сперматозоидом, так как его размеры больше. После инъекции яйцеклетку немедленно имплантируют в яйцевод приемной матери, или дают возможность развиваться в культуре до стадии бластоцисты, после чего имплантируют в матку. Можно вводить ген в сперматозоиды и затем проводить ими оплодотворение. Таким образом были инъецированы гены интерферона и инсулина человека, ген β-глобина кролика, ген тимидинкиназы вируса простого герпеса и кДНК вируса лейкемии мышей. Число молекул, вводимое за одну инъекцию, колеблется от 100 до 300 000, а их размер — от 5 до 50 кб. Выживает обычно 10–30 % яйцеклеток, а доля мышей, родившихся из трансформированных яйцеклеток варьирует от нескольких до 40 %. Таким образом, реальная эффективность составляет около 10 %. Интеграция чужеродных генов неспецифична по отношению к хромосомам, а число копий чужеродного гена может различаться от нескольких штук до 100 и более. Эти гены образуют группу тандемных повторов, объединенных по типу "голова к хвосту". Чужеродная ДНК после инъекции была обнаружена как в соматических, так и в половых клетках. Это означает, что интеграция проходит на самых ранних стадиях развития зиготы. В нескольких случаях гетерологичная ДНК наследовалась в трех поколениях мышей, что свидетельствует о стабильной интеграции. Интегрировавшая в половые клетки ДНК передается как менделевский ген. Установлено, что уровень экспрессии чужеродного гена зависит от места интеграции ДНК с хромосомами и от степени ее метилирования, а также от дифференцировки тканей. В некоторых случаях удалось получить тканеспецифическую экспрессию. Важно отметить что специфические чужеродные гены можно встраивать в геном клетки таким образом, что они подчиняются нормальным регуляторным сигналам. В 1981 году Константини и Лэси (Оксфорд) провели инъекцию в яйцеклетки мыши фрагменты хромосомной ДНК кролика длиной 19 килобаз. Эти фрагменты содержали ген р-глобина кролика. Яйцеклетки культивировали до стадии бластоцисты и имплантировали в матку. У 24 мышей, родившихся в результате развития имплантированных яйцеклеток, проведены частичная гепатоэктомия. Анализ ДНК из клеток печени показал, что у 9 мышей встречается от 1 до 20 копий на клетку гена β-глобина. После спаривания 4 трансформированных самцов с нормальными самками получили потомство из 18 животных. 6 из них также имели ген β-глобина. Установлено, что интеграция гена в клетки млекопитающих происходит случайным образом и не связана с конкретными областями хромосомы. Ген нестабилен, может быть утрачен или стать неактивным. Вместе с геном необходимо вводить регуляторные последовательности. Метод введения генов в эмбриональные клетки имеет ограничения. Не всегда удается встроить чужеродную ДНК в заданный участок хромосомы. Разработанные методические примы пока не позволяют заменить имеющийся в геноме ген, вытесняя его, не всегда удается подчинить новый ген системе регуляции организма. При трансгенозе могут возникать неожиданные проблемы. Например, одни из первых работ по генетической транформации животных проводились путем встраивания генов гормона роста. Перенос гена гормона роста крысы мышам увеличивал рост мышей в 2 раза. Эксперименты по трансгенозу генов гормона роста быка кроликам также увенчались успехом. А вот аналогичные эксперименты по модификации крупного рогатого скота привели к увеличению прироста всего на 10–20 %. Очевидно, это связанно с тем, что у мышей сохраняется широкая норма реакции, и встраивание генов, увеличивающих количество гормона, заставляет генотип реализоваться максимально полно. У домашнего скота в результате направленной селекции организмы работают на верхнем пределе нормы реакции, отсюда ожидаемый эффект не проявился. В нашей стране получены свиньи, несущие ген соматотропина. Они не отличались по темпам роста от нормальных животных, но изменение обмена веществ сказалось на содержании жира. У таких животных ингибировались процессы липогенеза и активировался синтез белка. К изменению обмена веществ приводило и встраивание генов инсулиноподобного фактора. Такие трансгенные свиньи были созданы для изучения цепочки биохимических превращений гормона, а побочным эффектом явилось укрепление иммунной системы. Самая мощная белоксинтезирующая система находится в клетках молочной железы. Если поставить гены чужих белков под контроль казеинового промотора, то экспрессия этих генов будет мощной и стабильной, а белок будет накапливаться в молоке (животное-ферментер). Уже получены трансгенные коровы, в молоке которых содержится человеческий белок лактоферрин. Этот белок планируют применять для профилактики гастроэнтерологических заболеваний у людей с низкой иммунорезистентностью. Это больные СПИДом, недоношенные младенцы, больные раком, прошедшие радиотерапию. Ведутся клинические испытания такого молока. Уже сейчас корпорация Genzyme Transgenics планирует исследования с целью создания трансгенного крупного рогатого скота, содержащего в молоке человеческий альбумин. Был куплен патент на получение эмбрионов, содержащих геном клеток соединительной ткани (фибробластов), включающий ген, ответственный за синтез человеческого белка. Подобная технология позволяет увеличить эффективность создания трансгенных молочных животных, так как при обычном впрыскивании генов в оплодотворенную яйцеклетку рождается от только 5 — 10 % трансформированных животных, из них — несколько самцов, не дающих молока. Использование новой технологии клонирования позволяет получать животных только женского пола, дающих трансгенный протеин. Альбумин используется в терапии для поддержания осмотического давления в крови. Ежегодно в мире требуется около 440 тысяч литров плазмы крови для выделения этого белка (стоимость около 1,5 млрд. $). Каждая молочная корова может произвести 80 кг рекомбинантного человеческого альбумина ежегодно. Genzyme Transgenics занимается разработкой аналогичных методов получения человеческого гормона роста и β-интерферона. В Англии созданы трансгенные овцы, молоко которых содержит фактор свертывания крови. В нашей стране были попытки создать овец, продуцирующих химозин (фермент для сыроварения). Было получено 2 овцы, у одной — ген не экспрессировался, у второй содержание химозина достигало 300 мг/л. Однако потомство этой овцы давало низкие удои — порядка 50 кг за период лактации. Причина заключалась в том, что химозин вырабатывается в виде предшественника — прохимозина, который превращается в активный фермент при рН=5. Было запланировано получать именно прохимозин, но в каких-то участках вымени происходило снижение pH, что приводило к активации химозина непосредственно в организме. Активный химозин свертывал молоко, а оно закупоривало протоки вымени. Сейчас пытаются решить эту проблему. В Подмосковье получены кролики, выделяющие γ-энтерферон/эритропоэтин, но кролики не являются традиционными продуцентами молока. Эксперименты же по трансформации сельскохозяйственных животных очень дорогостоящи — одно трансгенное животное стоит десятки и сотни тысяч долларов. Трансгенных животных получают и для целей ксенотрансплантации. Одним из излюбленных доноров органов являются свиньи, так как имеется анатомическое сходство органов и сходство иммунологических свойств. Реакции отторжения при трансплантации имеют сложный механизм. Одним из сигналов для атаки организма на чужой орган являются белки, локализованные на внешней поверхности мембраны. У трансгенных свиней эти белки заменены на человеческие. Еще одно направление трансгеноза — получение устойчивых к болезням животных. Животноводство держится на вакцинах, так как селекция ведется преимущественно на хозяйственно ценные признаки — шерстистость, молочность и т. д. Повышение устойчивости — дело генных инженеров. К защитным белкам относятся интерфероны, поэтому ген интерферона встраивали различным животным. Трансгенные мыши получили устойчивость, они не болели или болели мало, а вот у свиней такого эффекта не обнаружено. Другое направление — введение генов, кодирующих антисмысловую РНК. Для животноводства острой проблемой являются лейкозы, вызываемые РНК-вирусами. Трансгенные кролики, несущие гены, отвечающие за присутствие в клетке антисмысловой РНК, были устойчивы к лейкозам. Трансгенных животных можно использовать для изучения наследственных заболеваний мозга и нервной системы. Гены болезни Альцгеймера (отложение белка β-амилоида приводит к образованию характерных бляшек) и гены, отвечающие за развитие эпилепсии, болезней мозга вводятся в геном нормальных животных; при этом получают трансгенных животных-моделей, на которых можно испытывать различные терапевтические приемы. Трансгенных животных стали использовать для исследования воспалительных и иммунологических заболеваний человека, например, ревматоидного артрита. Моделируются болезни, связанные с липидным обменом.
ГЕННАЯ ИНЖЕНЕРИЯ РАСТЕНИЙ
Трансформация растительного генома Генетическая конструкция, вводимая в растительную клетку обычно включает: белок-кодирующую структурную последовательность, сигнальные элементы трансляции и транскрипции, а также маркерные гены. Наиболее важными из регуляторных последовательностей являются проксимальный участок промотора, связывающий РНК-полимеразу; участок, кодирующий 5'-конец мРНК, необходимый для связывания с рибосомой и инициации трансляции, и эукариотический сигнал полиаденилирования на 3'-конце мРНК. Среди эукариотических организмов эти конститутивные сигнальные элементы оказались, к счастью, высококонсервативными и достаточно универсальными, так что растительные клетки в основном правильно экспрессируют чужеродные гены не только растений других видов, но и млекопитающих, дрожжей и других эукариот. Однако для генов бактериального происхождения необходима замена их конститутивных сигнальных элементов на соответствующие эукариотические. Помимо этого, для лучшей экспрессии гена на уровне трансляции мРНК желательно приблизить набор кодонов к типичному для растения. Обычно для этого посредством направленных точечных мутаций заменяют "редкие" кодоны на синонимичные "частые", что не сказывается на первичной структуре белка. В результате экспрессия гена может быть усилена до 300 раз. Иногда в структурной части генов прокариотического происхождения могут присутствовать какие-либо нежелательные сигнальные последовательности, например, узнаваемые на уровне мРНК ферментами сплайсинга или деградации, либо ферментами модификации на уровне белка. Наличие таких скрытых ("криптических") сигналов ведет к резкому снижению экспрессии гена в растении, поэтому их обычно удаляют также путем точечных замен оснований. Минимальный промотор, связывающий РНК-полимеразу, как правило, недостаточен для обеспечения заметного, а тем более тканеспецифичного уровня транскрипции. Для усиления экспрессии встроенного гена и придания ей заданных характеристик используют полноразмерные промоторы, включающие энхансеры (усилители) и (или) факторзависимые цис-элементы. Это приводит к тому, что подготовленный для трансформации ген, как правило, является химерным, т. е. включает фрагменты ДНК из одного вида, соединенные с фрагментами ДНК из другого вида. Набор известных к настоящему дню промоторов достаточно разнообразен и постоянно пополняется. Конститутивные промоторы применяются для наработки существенных количеств продукта гена во всем растении. Для двудольных растений такими эффективными промоторами являются, например, З5S-промотор вируса мозаичности цветной капусты (CaMV) и поэ-промотор гена нопалин-синтазы агробактерий; для однодольных — промоторы гена алкогольдегидрогеназы кукурузы (Adh) и гена актина 1 риса (Act). Помимо конститутивных, известно большое число специфических промоторов, которые активны лишь в отдельных органах, тканях или клетках, либо на отдельных стадиях онтогенеза растения. Примером может служить промотор гена пататина картофеля, работающий практически только в клубнях. Имеются также промоторы, активность которых проявляется в листьях, корнях, меристемах и других местах специфической локализации. Интенсивно изучаются и используются также индуцибельные промоторы, которые активируются лишь при определенных условиях: тем пературы, освещения, концентрации фитогормонов и т. д. Многие из таких промоторов достаточно универсальны, например, некоторые промоторы генов теплового шока. В частности, промотор гена hsp70 из дрозофилы равно эффективен в клетках растений. Особый интерес представляют промоторы, индуцируемые низкомолекулярными химическими эффекторами, часто не свойственными растениям. В зависимости от типа промотора, индукторами могут служить тетрациклин, дексаметазон, бензотиадиазол, этанол, ионы меди и другие соединения. Эти промоторы очень важны для фундаментальных исследований трансгенных растений, позволяя четко дифференцировать первичные и вторичные эффекты изучаемого гена и тем самым прояснить его истинную биологическую функцию. Они перспективны также для биотехнологии, так как позволяют вызвать экспрессию гена в заданный период, когда она уже либо не препятствует нормальному росту и развитию растения, либо не вызывает иных отрицательных последствий. Регулируемые извне индуцибельные промоторы, контролирующие соответствующие гены, могут способствовать одновременному прохождению растениями основных стадий онтогенеза (переход к цветению, опадение листьев и др.), что важно для практики сельского хозяйства. Есть промотор, индуцирующийся при механическом стрессе (поранении) или при обработке растений элиситорами. Использование такого промотора, соединенного с целевым геном, дает возможность выращивать трансгенные растения как обычные вплоть до стадии уборки урожая, а далее срезка растений индуцирует экспрессию целевого гена, продукт которого накапливается в собранной биомассе.
Введение генов в растительные клетки Ввести чужеродную ДНК в растения можно различными способами. Для двудольных растений существует естественный вектор для горизонтального переноса генов: плазмиды агробактерий. Что касается однодольных, то, хотя в последние годы достигнуты определенные успехи в их трансформации агробактериальными векторами, все же подобный путь трансформации встречает существенные затруднения. Для трансформации устойчивых ("рекальцитрантных") к агробактериям растений разработаны приемы прямого физического переноса ДНК в клетку, многие из которых взяты из практики работы с клетками бактерий или животных. Эти методы достаточно разнообразны, они включают: бомбардировку микрочастицами или баллистический метод; электропорацию; обработку полиэтиленгликолем; перенос ДНК в составе липосом и др. Наиболее продуктивным и чаще всего используемым является метод бомбардиров ки микрочастицами. При достаточной скорости эти частицы могут непосредственно проникать в ядро, что сильно повышает эффективность трансформации. Этим же методом можно, впрочем, трансформировать и другие ДНК-содержащие клеточные органеллы — хлоропласты и митохондрии. В последнее время был разработан и успешно применен также комбинированный метод трансформации, названный агролистическим. При этом чужеродная ДНК вводится в ткани каким-либо физическим методом, например, баллистическим. Вводимая ДНК включает как Т-ДНК вектор с целевым и маркерным геном, так и агробактериальные гены вирулентности, поставленные под эукариотический промотор. Временная экспрессия генов вирулентности в растительной клетке приводит к синтезу белков, которые правильно вырезают Т-ДНК из плазмиды и встраивают ее в хозяйский геном, как и при обычной агробактериальной трансформации. После проведения тем или иным способом трансформации растительной ткани ее помещают in vitro на специальную среду с фитогормонами, способствующую размножению клеток. Среда обычно содержит селективный агент, в отношении которого трансгенные, но не контрольные клетки приобретают устойчивость. Регенерация чаще всего проходит через стадию каллуса, после чего при правильном подборе сред начинается органогенез (побегообразование). Сформированные побеги переносят на среду укоренения, часто также содержащую селективный агент для более строгого отбора трансгенных особей.
Экспрессия генетического материала в трансгенных растениях При изучении экспрессии чужеродных генов в растительных клетках исследователи столкнулись с рядом новых явлений. Выяснилось, что трансформированные идентичной конструкцией ДНК трансгенные клоны, полученные параллельно в одном и том же опыте, значительно различаются по уровню экспрессии введенного гена. Показано, что уровень экспрессии зависит от многих факторов и в значительной мере от того в какую область ядерного хроматина попал введенный ген. Экспрессия трансгена, как правило, высока при его попадании в область активного хроматина. Кроме того, оказалось, что при встраивании в ядерный геном конструкция ДНК нередко претерпевает существенные изменения (перестройки, дупликации, инверсии и т. д.), что также приводит в основном к снижению экспрессии. Было установлено также, что обычно применяемые процедуры трансформации вовсе не безразличны и для хозяйского генома. Во-первых, встраивание трансгена может нарушить первичную структуру какого-либо хозяйского гена и тем самым вызвать его инактивацию. Это событие, по-видимому, не так редко, особенно с учетом того, что трансгены чаще встраиваются в транскрибируемые области хроматина (эухроматин). В последующих поколениях такой инактивированный ген может перейти в гомозиготное состояние, выражаясь в непредусмотренной и обычно нежелательной фенотипической мутации. Во-вторых, при агробактериальном или физическом переносе генов в растительный геном в последнем нередко отмечаются разного рода перестройки, вплоть до транслокации фрагментов хромосом. Все это также может менять нормальное функционирование генома растения. Еще одна важная проблема, которая открылась благодаря генетической инженерии растений — это проблема замолкания генов (gene silencing). Было обнаружено, что у достаточно заметной доли трансгенных растений интродуцированный ген через какое-то время теряет свою активность, хотя физически сохраняется в геноме. Таким образом, было установлено, что растение обладает способностью активно противостоять экспрессии чужеродной ДНК. Проблема замолкания генов имеет большое практическое значение, так как у генетически трансформированных сельскохозяйственных культур трансгены должны функционировать стабильно. За последние годы удалось выяснить механизмы и некоторые условия, способствующие замолканию генов. Одним из основных факторов является число идентичных копий гена, встроенных в геном: чем больше таких копий и чем они протяженнее, тем больше вероятность замолкания генов. Откуда следуют практические рекомендации генным инженерам: трансгенные культуры должны содержать не более одного встроенного гена на гаплоидный геном; сигнальные части трансгена (промоторы, терминаторы и др.) не должны иметь длинных гомологий (более 100–300 п.о. ДНК) с участками хозяйского генома; фрагменты векторной плазмидной или вирусной ДНК должны быть по возможности полностью удалены из конструкции перед трансформацией. Уровень и стабильность экспрессии трансгенов в растении можно повысить при специальном конструировании концов вводимой ДНК. Так, добавление терминаторов транскрипции предотвращает нежелательное прохождение через трансген РНК-полимеразы, если промотор хозяйского генома случайно оказывается поблизости от места встраивания. При концевом добавлении сравнительно коротких (>300 п.о.) участков связывания ядерного матрикса (МА.Р) трансген имеет больше шансов образовать независимую петлю хроматина или включиться в состав транскрибируемого эухроматина. В обоих случаях это приводит к повышению и стабилизации уровня экспрессии трансгена. В ряде случаев, особенно у злаков, введение интрона в район 5'-конца мРНК целевого гена резко усиливает его транскрипцию. Помимо этого, в составе мРНК обнаружены цис-элементы, определяющие степень ее стабильности в растительной клетке. Правильный генно-инженерный дизайн таких элементов может также способствовать оптимальному накоплению мРНК в трансгенных растениях, предназначенных для биотехнологического применения. Стабильно введенный ген должен передаваться потомству при семенном размножении, поэтому важно определить этот параметр на практике. Уровень транскрипции введенного гена оценивается обычно с помощью традиционного Northern-блотинга или недавно предложенных количественных версий RT PCR (на мРНК с помощью обратной транскриптазы синтезируется комплементарная цепь ДНК, которая и размножается в полимеразной цепной реакции). В ряде случаев экспрессию гена проще и надежнее контролировать по конечному белковому продукту или тому эффекту, который этот белок вызывает. Однако необходимо использование разных методов для доказательства истинной трансгенности данного растения, чтобы исключить "псевдотрансгенные" черты, вызванные сомаклональной (эпигенетической) изменчивостью или неполной элиминацией агробактерий Для изучения молекулярной конституции генома трансгенных организмов можно использовать полиморфизм длины рестрикционных фрагментов. Метод основан на том, что гомологичные последовательности ДНК гибридов и их родителей при разрезании рестриктазами могут попасть во фрагменты разной длины. В качестве маркеров родительских геномов можно использовать и часто повторяющиеся последовательности ДНК. Повторяющиеся последовательности составляют значительную часть генома типичного растения. Большая часть последовательностей повторяется до 1000 раз на геном. У многих растений выявлены еще более частые повторы (100000 — 1000000 копий на геном), которые составляют до 20 % генома. Они имеют отличный от остальной клеточной ДНК "Г-Ц" состав и в градиенте хлористого цезия выявляются в виде сателлитного компонента. В геномах даже очень близких видов амплифицируются разные типы повторов, поэтому они становятся своеобразными "отпечатками пальцев" близкородственных геномов. Высокотандемные последовательности ДНК относительно легко клонируются, так как после расщепления рестриктазой, имеющей сайт узнавания в повторе, эти последовательности легко выявляются в виде полос при электрофоретическом разделении фрагментов и могут быть элюированы из геля в чистом виде. Их можно использовать в качестве зондов. Использование высокоповторяющихся последовательностей в качестве молекулярных зондов позволяет вести одновременный скрининг большого количества образцов грубоочищенной ДНК регенерировавших после слияния протопластов растений или каллусов методом дот-гибридизации. Как другой биохимический маркер при анализе соматических гибридов может использоваться рибулозо-1,5-бифосфат карбоксилаза/оксигеназа. Причины: необычайно высокое содержание его в листьях (до 50 % растворимого белка), хорошо разработанные методы выделения и анализа, а также то, что составляющие этот белок субъединицы кодируются как ядерным (мол. масса 14 кД), так и хлоропластным (большая субъединица, мол. масса 55 кД) геномами.
Характеристика Ti- и Ri-плазмид A. tumefaciens вызывает образование опухолей стебля двудольных растений — так называемых корончатых галлов. Бактерии прикрепляются к клетками растения в местах повреждений. Сайтами связывания на поверхности бактерий, видимо, являются молекулы β-глюкана и О-антигенной цепи липополисахарида внешней мембраны. Бактерии связываются с рецепторами высшего растения, состоящими из белка и пектина; лектины в данном случае не имеют значения. Бактериальные сайты связывания и рецепторы растений являются констуитивными, т. е. оба партнера обладают ими еще до момента взаимодействия. Первый шаг взаимодействия с растением — узнавание — следует рассматривать как специфическую адгезию растений. Как только бактерии прикрепились к поверхности клеток растения, они начинают образовывать целлюлозные фибриллы. Эти фибриллы можно увидеть в сканирующем электронном микроскопе уже через 90 минут после добавления бактерий к суспензии культуры клеток ткани моркови. К 10 часам инкубации фибриллы формируют сеть, покрывающую поверхность растительных клеток. Фибриллы служат более прочному закреплению бактерий на поверхности хозяина. За целлюлозные фибриллы могут зацепиться свободно плавающие клетки бактерий. Фиксируя их у поверхности растения, фибриллы увеличивают множественность заражения. В результате размножения образуются скопления бактерий на поверхности растения. Клеточная стенка растения повреждается вследствие выделения бактериями пектолитических ферментов, что обеспечивает плотный контакт бактерий с плазмалеммой растительной клетки. Этот контакт необходим для передачи ДНК от бактерий в растительную клетку. Передача ДНК происходит без нарушения целостности мембраны растительной клетки, но требует определенного её состояния — компетентности. Способность A. tumefaciens индуцировать у растений образование опухолей типа "корончатого галла" коррелирует с наличием у них Ti-плазмиды. Опухолевая трансформация проявляется в гипертрофии возникающей после проникновения агробактерий в пораненные участки (сайты) растений (рис. 47). Трансформация является результатом стабильного ковалентного включения (инсерции или интеграции) сегмента («transferred» или Т-ДНК) большой плазмиды (pTi — tumor inducing или pRi — root inducing) бактерий в ядерную ДНК растительной клетки.

Рис. 47. Генетическая колонизация растения A. tumefaciens: 1 — агробактерии существуют в ризосфере; 2 — строение A. tumefaciens; 3 — встраивание Т-ДНК в геном; 4 — образование опухоли.
Другой вид агробактерий — A. rhizogenes, — вызывает заболевание, именуемое "бородатый корень", при котором в зоне повреждения корня образуется масса новых корешков. A. rubi обычно индуцируют неорганизованные опухоли (тератомы), штаммы A. radiobacter авирулентны. В отличие от большинства тканей взятых из нормальных растений, трансформированные ткани в культуре in vitro в асептических (стерильных) условиях способны неограниченно расти в отсутствие экзогенно добавленных ауксинов и цитокининов. Кроме того, трансформированные ткани часто синтезируют одну или более групп соединений, названных опинами, которые обычно не обнаруживаются в нетрансформированных растительных тканях. Наиболее подробно изучены опухоли — корончатые галлы, индуцируемые Agrobacterium tumefaciens. Они представляют собой истинно злокачественные опухоли, которые могут расти в культуральной среде в отсутствие стимуляторов роста — фитогормонов, необходимых для роста нормальных тканей. Опухоли можно поддерживать в течение многих лет in vitro, и при их использовании они способны вызывать опухоли у здоровых растений. В природных условиях корончатые галлы образуются в месте соединения корня со стеблем (у корневой шейки), откуда и произошло их название корончатый галл. Однако корончатые галлы могут развиваться и на подземных частях растения, например, на корнях плодовых деревьев, и на надземных, например, на стебле винограда. В лаборатории эти заболевания можно вызвать у здоровых растений экспериментально, путем инфицирования их бактериями. Растения перед инокуляцией должны быть поранены, при этом опухоли возникают в поврежденных сайтах растения, обычно на стебле или листьях растения. Кроме целых растений в качестве тест-объектов используются экспланты, например ломтики моркови и кусочки других органов растений. Ткани корончатых галлов содержат более высокие уровни ауксина и цитокининов. Выявлено еще одно наследуемое изменение в клетках корончатых галлов — это синтез опинов. Необычное для растений соединение, производное аргинина, обнаруженное лишь в определенных опухолевых линиях, было названо октопином. Затем было показано, что другими опухолевыми линиями синтезируется еще одно соединение — нопалин, также производное аргинина. В зависимости от типа индуцируемого в опухоли опина штаммы A. tumefaciens и находящиеся в них Ti-плазмиды получили соответствующее обозначение — октопиновые или нопалиновые. Агробактерии, индуцирующие опухоли, в которых не обнаруживается ни нопалин, ни октопин, ранее обозначались как штаммы нулевого типа. Позднее было показано, что в опухолях нулевого типа синтезируются опины третьего класса — агропины. Обнаружены также и другие типы опинов. Поскольку все опины обнаруживаются только в опухолевых клетках и отсутствуют в клетках нормальных растений или клетках растительных опухолей других типов, то опины могут рассматриваться как специфические биохимические маркеры для клеток корончатых галлов. Опухоли, развивающиеся из одной или нескольких клеток, быстро разрастаются в крупные образования, диаметр которых на определенных видах деревьев может достигать одного метра. Типичная неорганизованная опухоль представляет собой более или менее округлую дедифференцированную массу клеток (каллус), которая может иметь гладкую или шероховатую поверхность, быть паренхиматозной или одревесневшей. Иногда на периферии таких опухолей формируются листовидные структуры (тератомы), иногда — придаточные корни. Нередко на зараженных растениях наблюдаются вторичные опухоли, значительно удаленные от первичных. Обычно они обнаруживаются выше первичной опухоли, что предполагает движение бактерий или трансформирующего агента в направлении транспирации. Распространение Agrobacterium и других фитопатогенных бактерий по межклетникам и ксилеме является хорошо доказанным фактом. Агробактерии могут передвигаться на большие дистанции со значительной скоростью. Очевидно, это не является единственной причиной индукции вторичных опухолей. Организацию опухолей, а именно форму, величину и характер развития, определяют три фактора: • штамм агробактерий, • генотип растения-хозяина, • физиологическое состояние инфицируемых растительных клеток. Agrobacterium имеет очень широкий круг растений-хозяев и может инфицировать практически все двудольные растения. Долгое время считалось, что однодольные растения не чувствительны к агробактериальной инфекции. В настоящее время показано, что при соблюдении определенных условий агробактерии могут инфицировать однодольные растения, в частности представителей таких семейств, как Amaryllidaceae, Liliaceae, Gramineae, Iridaceae и некоторых других. Однако существуют определенные вариации круга хозяев для различных штаммов Agrobacterium: некоторые штаммы способны вызывать галлообразование на отдельных видах растений, но не инфицируют другие. Различные сорта одного и того же растения также могут иметь различную чувствительность к данному бактериальному штамму. Невозможность заражения в природе обуславливается отсутствием соответствующих рецепторов, необходимых для взаимодействия с бактериями. Другим фактором, препятствующим инфицированию однодольных агробактериями, возможно, является отсутствие в клетках растений низкомолекулярных индукторов вирулентности Agrobacterium, например ацетосирингона, которые обычно присутствуют в клеточном соке при поранении двудольных растений. Подробная информация о структуре плазмид Agrobacterium получена путем их рестрикционного или физического картирования. В результате исследований обнаружено четыре основных области гомологии между октопиновой и нопалиновой плазмидами. Две консервативные (области А и D) вовлечены в онкогенность, еще одна (В) соответствует области контроля репликации плазмиды, в то время как последняя (С) кодирует функции конгьюгативного переноса (рис. 48).

Рис. 48. Структура Ti-плазмиднопалинового и октопинового типа
Таким образом, кроме Т-ДНК в плазмидах имеются область, кодирующая функцию конъюгации (Тrа), область репликации (Ori V) и область вирулентности (Vir). Последовательности Ti-плазмиды, фланкирующие Т-ДНК (пограничные или концевые области), играют важную роль в интеграции в растительный геном и содержат несовершенные прямые повторы по 24–25 п.н. Делеция левой границы Т-ДНК не влияет на опухолеобразование, но удаление правой пограничной области приводит практически к полной утрате вирулентности. Показано, что делеция правого повтора или его части приводит к потере способности Т-ДНК включаться в растительную ДНК. Учитывая важную роль концов Т-области в переносе Т-ДНК, можно предположить, что любой сегмент ДНК, встроенный между этими концами, может быть перенесен в растения как часть Т-ДНК. Плазмиды модифицируют таким образом, чтобы удалить все онкогенные последовательности, так как они не принимают участие ни в переносе, ни в интеграции в геном клетки-хозяина. На место этих генов можно встроить чужеродную ДНК, а плазмида теряет свои онкогенные свойства. Неонкогенные Т-ДНК, присутствующие в растениях — регенерантах, передаются согласно законам Менделя.
Введение ДНК в клетки растений с помощью Ti- и Ri-плазмид Разработаны два метода для введения Ti-плазмидных последовательностей, содержащих нужный ген, в растение. Первый метод — метод «промежуточных векторов» (коинтегративных векторов) — основан на использовании плазмиды кишечной палочки pBR 322 (рис. 49).

Рис. 49. Создание коинтегративного вектора на основе Ti-плазмиды: Рр — расщепление рестриктазой
Т-ДНК вырезают из Ti-плазмиды с помощью рестриктаз и встраивают в плазмиду pBR 322 для клонирования в Е. coli. Бактерии, содержащие плазмиду с Т-ДНК, размножают, после чего эту плазмиду выделяют. Затем в клонированную Т-ДНК с использованием рестриктаз встраивают нужный ген. Эту рекомбинантную молекулу, содержащую Т-ДНК со встроенным в нее геном, снова размножают в большом количестве, то есть клонируют в кишечной палочке. Затем с помощью конъюгации вводят в клетки агробактерии, несущие полную Ti-плазмиду. Между Т-сегментами нативной Ti-плазмиды и промежуточного вектора происходит гомологичная рекомбинация. В результате этого Т-ДНК со встроенным геном включается в нативную Ti-плазмиду, замещая нормальную ДНК. Получаются клетки А. tumefaciens, несущие Ti-плазмиды со встроенными в Т-сегмент нужными генами. Далее их перенос в клетки растения осуществляется обычным способом, характерным для агробактерий. Второй метод основан на созданиисистемы бинарных (двойных) векторов. Последние исследования показали, что для заражения и трансформации не нужна целая Ti-плазмида, а достаточны только пограничные области Т-ДНК и один участок Ti-плазмиды, ответственный за вирулентность. Причем эти два участка ДНК не обязательно должны находиться в одной и той же плазмиде. Если клетки агробактерий содержат Ti-плазмиду с сегментом vir и другую плазмиду с Т-ДНК, эти бактерии могут трансформировать клетки растений. При этом Т-ДНК с любыми встроенными в нее генами интегрирует с геномом растения, для этого не нужна гомологичная рекомбинация в бактериальных клетках. Для осуществления экспрессии чужеродных генов, нужен специфический промотор из Т-ДНК, например, промотор нопалинсинтетазы. Показано, что он функционирует в клетках растений и может быть легко соединен с кодирующей последовательностью чужеродного гена в широко распространенных субклонах Ti-плазмид. Другое преимущество данного промотора заключается в том, что он функционирует в каллусах и в большинстве органов растений. Эффективность трансформации с помощью модифицированной Т-ДНК агробактерий превосходит на сегодняшний день все другие способы переноса генов в растение. О механизмах, с помощью которых агробактерия переносит Т-ДНК ядра растений, известно очень мало: Т-сегменты ДНК октопиновых и нопалиновых плазмид встраиваются в разные, по-видимому, случайные, точки хромосом хозяина, но при этом они никогда не интегрируют с ДНК митохондрий и хлоропластов. Для введения сконструированных Ti-плазмид в растительную клетку может быть использовано несколько методов. Наиболее простой из них природный способ — это инокуляция сконструированных штаммов в поврежденные (пораненные) области растения. Другой метод состоит в трансформации протопластов путем сокультивирования их с агробактериями. Методика сокультивации может рассматриваться как индукция опухолей в искусственных условиях: вирулентные агробактерии временно совместно культивируются с протопластами. Если агробактерии добавляются к свежевыделенным или однодневным протопластам, не наблюдается ни присоединения бактерий, ни трансформации. Существенным условием для трансформации является наличие вновь образуемых клеточных стенок у 3-дневных протопластов. Это подтверждается применением ингибиторов образования клеточной стенки, которые ингибируют и присоединение бактерий. После периода сокультивации (более суток), в течение которого наступает агрегация протопластов с бактериями, свободные бактерии удаляются повторным отмыванием. Далее растительные клетки культивируются на среде с добавлением гормонов, а через 3–4 недели небольшие колонии высеваются на безгормональную среду. На этой среде выживают только колонии трансформированных клеток. Так были получены трансформированные растения-регенеранты табака и петунии. Этот метод дает возможность существенно расширить круг хозяев агробактерий, включая виды семейства злаковых. Эффективность кокультивирования может быть повышена применением индукторов слияния клеток (ПЭГ, кальций и др.). Трансформация протопластов может быть проведена также сокультивированием их непосредственно с Ti-плазмидами, такие опыты были проведены с протопластами петунии, табака. Очень низкая эффективность включения Т-ДНК в протопласты, наблюдавшаяся в первых экспериментах, была затем увеличена благодаря химической стимуляции (ПЭГ). Из трансформированных клеток были получены трансгенные растения. Преимуществом этого метода является то, что отпадает необходимость в промежуточных векторах.
Достижения генной инженерии растений Первые трансгенные растения (растения табака со встроенными генами из микроорганизмов) были получены в 1983 г. Первые успешные полевые испытания трансгенных растений (устойчивые к вирусной инфекции растения табака) были проведены в США уже в 1986 г. После прохождения всех необходимых тестов на токсичность, аллергенность, мутагенность и т. д. первые трансгенные продукты появились в продаже в США в 1994 г. Это были томаты Flavr Savr с замедленным созреванием, созданные фирмой "Calgen", а также гербицид-устойчивая соя компании "Monsanto". Уже через 1–2 года биотехнологические фирмы поставили на рынок целый ряд генетически измененных растений: томатов, кукурузы, картофеля, табака, сои, рапса, кабачков, редиса, хлопчатника. В настоящее время получением и испытанием генетически модифицированных растений занимаются сотни коммерческих фирм во всем мире с совокупным капиталом более ста миллиардов долларов. В 1999 г. трансгенные растения были высажены на общей площади порядка 40 млн. га, что превышает размеры такой страны, как Великобритания. В США генетически модифицированные растения (GM Crops) составляют сейчас около 50 % посевов кукурузы и сои и более 30–40 % посевов хлопчатника. Это говорит о том, что генно-инженерная биотехнология растений уже стала важной отраслью производства продовольствия и других полезных продуктов, привлекающей значительные людские ресурсы и финансовые потоки. В ближайшие годы ожидается дальнейшее быстрое увеличение площадей, занятых трансгенными формами культурных растений. Первая волна трансгенных растений, допущенных для практического применения, содержала дополнительные гены устойчивости (к болезням, гербицидам, вредителям, порче при хранении, стрессам). Нынешний этап развития генетической инженерии растений получил название "метаболическая инженерия". При этом ставится задача не столько улучшить те или иные имеющиеся качества растения, как при традиционной селекции, сколько научить растение производить совершенно новые соединения, используемые в медицине, химическом производстве и других областях. Этими соединениями могут быть, например, особые жирные кислоты, полезные белки с высоким содержанием незаменимых аминокислот, модифицированные полисахариды, съедобные вакцины, антитела, интерфероны и другие "лекарственные" белки, новые полимеры, не засоряющие окружающую среду и многое, многое другое. Использование трансгенных растений позволяет наладить масштабное и дешевое производство таких веществ и тем самым сделать их более доступными для широкого потребления.
Улучшение качества запасных белков Запасные белки основных культурных видов кодируются семейством близкородственных генов. Накопление запасных белков семян — сложный биосинтетический процесс. Первая генно-инженерная попытка улучшения свойства одного растения путем введения гена запасного белка от другого была, проведена Д. Кемпом и Т. Холлом в 1983 г. в США. Ген фазеолина бобов с помощью Ti-плазмиды был перенесен в геном подсолнечника. Результатом этого опыта было лишь химерное растение, получившее название санбин. В клетках подсолнечника были обнаружены иммунологически родственные фазеолиновые полипептиды, что подтверждало факт переноса гена между растениями, относящимися к различным семействам Позднее ген фазеолина был передан клеткам табака: в растениях-регенерантах ген экспрессировался во всех тканях, хотя и в малых количествах. Неспецифическая экспрессия фазеолинового гена, так же как и в случае переноса его в клетки подсолнечника, сильно отличается от экспрессии этого гена в зрелых семядолях бобов, где фазеолин составлял 25–50 % от общего белка. Этот факт указывает на необходимость сохранения и других регуляторных сигналов этого гена при конструировании химерных растений и на важность контроля экспрессии генов в процессе онтогенеза растений. Ген, кодирующий запасной белок кукурузы — зеин, после интеграции его в Т-ДНК был перенесен в геном подсолнечника следующим образом. Штаммы агробактерий, содержащие Ti-плазмиды с геном зеина, использовали для индукции опухолей в стеблях подсолнечника. Некоторые из полученных опухолей содержали мРНК, синтезируемые с генов кукурузы, что дает основание рассматривать эти результаты как первое доказательство транскрипции гена однодольного растения в двудольном. Однако присутствие зеинового белка в тканях подсолнечника не обнаружилось. Более реальной задачей для генетической инженерии считается улучшение аминокислотного состава белков. Как известно, в запасном белке большинства злаковых наблюдается дефицит лизина, треонина, триптофана, у бобовых — метионина и цистеина. Введение в эти белки дополнительных количеств дефицитных аминокислот могло бы ликвидировать аминокислотный дисбаланс. Методами традиционной селекции удалось существенно повысить содержание лизина в запасных белках злаковых. Во всех этих случаях часть проламинов (спирторастворимые запасные белки злаковых) заменялась другими белками, содержащими много лизина. Однако у таких растении уменьшались размеры зерна и снижалась урожайность. По-видимому, проламины необходимы для формирования нормального зерна, и их замена другими белками отрицательно влияет на урожайность. Учитывая это обстоятельство, для улучшения качества запасного белка зерновых нужен такой белок, который не только отличался бы высоким содержанием лизина и треонина, но и мог полноценно заменить определенную часть проламинов при формировании зерна. Растения могут производить и белки животного происхождения. Так, встраивание в геном растений Arabidopsis thaliana и Brassica napus химерного гена, состоящего из части гена запасного 25-белка арабидопсиса и кодирующей части для нейропептида — энкефалина, приводило к синтезу химерного белка до 200 нг на 1 г семени. Два структурных белковых домена были связаны последовательностью, узнаваемой трипсином, что давало возможность в дальнейшем легко изолировать чистый энкефалин. В другом эксперименте удалось после скрещивания трансгенных растений, в одном из которых был встроен ген гамма-субъединицы, а во втором — ген каппа-субъединицы иммуноглобулина, получить у потомства экспрессию обеих цепей. В результате растение формировало антитела, составляющие до 1,3 % суммарного белка листьев. Также было показано, что в растениях табака могут собираться полностью функциональные секреторные моноклональные иммуноглобулины. Секреторные иммуноглобулины обычно выделяются в ротовую полость и желудок человека и животных и служат первым барьером на пути кишечных инфекций. В упомянутой выше работе получили продукцию в растениях моноклональных антител, которые были специфичны для Streptococcus mutans — бактерий, вызывающих зубной кариес. Предполагается, что на основе таких моноклональных антител, продуцируемых трансгенными растениями, удастся создать действительно антикариесную зубную пасту. Из других белков животного происхождения, которые представляют интерес для медицины, показана продукция в растениях человеческого β-интерферона. Разработаны также подходы, позволяющие получать бактериальные антигены в растениях и использовать их в качестве вакцин. Получен картофель, экспрессирующий олигомеры нетоксичной субъединицы β-токсина холеры. Эти трансгенные растения могут быть использованы для получения дешевой вакцины от холеры.
Жиры Важнейшим сырьем для получения разного рода химических веществ являются жирные кислоты — основной компонент растительного масла. По своей структуре это углеродные цепи, которые обладают различными физико-химическими свойствами в зависимости от своей длины и степени насыщения углеродных связей. В 1995 году была закончена экспериментальная проверка и получено разрешение от федеральных властей США на выращивание и коммерческое использование трансгенных растений рапса с измененным составом растительного масла, включающего вместе с обычными 16- и 18-членными жирными кислотами также и до 45 % 12-членной жирной кислоты — лаурата. Это вещество широко используется для производства стиральных порошков, шампуней, косметики. Экспериментальная работа заключалась в том, что был клонирован ген специфической тиоэстеразы из растения Umbellularia califomica, где содержание лаурата в жире семян достигало 70 %. Структурная часть гена этого фермента под контролем промотора-терминатора гена белка, специфического для ранней стадии семяобразования, была встроена в геном рапса и арабидопсиса, что и привело к увеличению содержания лаурата в масле этих растений. Из других проектов, связанных с изменением состава жирных кислот, можно упомянуть работы, ставящие целью повышение или снижение содержания ненасыщенных жирных кислот в растительном масле. Интересными представляются эксперименты с петрозелиновой кислотой — изомером олеиновой кислоты, где двойная связь находится за шестым углеродным членом. Эта жирная кислота входит в со став масла кориандра и определяет его более высокую температуру плавления (33 °C), в то время как при наличии олеиновой кислоты температура плавления составляет только 12 °C. Предполагается, что после переноса генов, определяющих синтез петрозелиновой кислоты, в растения — продуценты растительного масла удастся производить диетический маргарин, содержащий ненасыщенную жирную кислоту. Кроме того, из петрозелиновой кислоты очень легко получать лаурат путем окисления озоном. Дальнейшее изучение специфики биохимического синтеза жирных кислот, по-видимому, приведет к возможности управлять этим синтезом с целью получения жирных кислот различной длины и различной степени насыщения, что позволит значительно изменить производство детергентов, косметики, кондитерских изделий, затвердителей, смазочных материалов, лекарств, полимеров, дизельного топлива и многого другого, что связано с использованием углеводородного сырья.
Полисахариды Проводится работа по созданию трансгенных растений картофеля и других крахмалнакапливающих культур, в которых это вещество будет находиться в основном в виде амилопектина, то есть разветвленной форме крахмала, или же в основном только в виде амилозы, то есть линейных форм крахмала. Раствор амилопектина в воде более жидкий и прозрачный, чем у амилозы, которая при взаимодействии с водой образует ригидный гель. Так, например, крахмал, состоящий в основном из амилопектина, по-видимому, будет иметь спрос на рынке производителей различных питательных смесей, где сейчас в качестве наполнителя используется модифицированный крахмал. Генетической модификации могут подвергаться также геномы пластид и митохондрий. Такие системы позволяют значительно увеличить содержание продукта в трансгенном материале.
Создание гербицидоустойчивых растений В новых, интенсивных сельскохозяйственных технологиях гербициды применяются очень широко. Это связано с тем, что на смену прежним экологически опасным гербицидам широкого спектра действия, обладающим токсичностью для млекопитающих и длительно сохраняющимся во внешней среде, приходят новые, более совершенные и безопасные соединения. Однако они обладают недостатком — подавляют рост не только сорняков, но и культурных растений. Такие высокоэффективные гербициды, как, глифосат, атразины интенсивно изучаются на предмет выявления механизма толерантности к ним некоторых сорняков. Так, на полях, где широко используют атразин, довольно часто появляются атразинустойчивые биотипы у многих видов растении. Изучение механизма устойчивости к гербицидам с целью получения методами генетической инженерии культурных растений, обладающих этим признаком, включает следующие этапы: выявление биохимических мишеней действия гербицидов в растительной клетке: отбор устойчивых к данному гербициду организмов в качестве источников генов устойчивости: клонирование этих генов: введение их в культурные растения и изучение их функционирования Существуют четыре принципиально различных механизма, которые могут обеспечивать устойчивость к тем или иным химическим соединениям, включая гербициды: транспортный, элиминирующий, регуляционный и контактный. Транспортный механизм устойчивости заключается в невозможности проникновения гербицида в клетку. При действии элиминирующего механизма устойчивости вещества, попавшие внутрь клетки, могут разрушаться с помощью индуцируемых клеточных факторов, чаще всего деградирующих ферментов, а также подвергаться тому или иному виду модификации, образуя неактивные безвредные для клетки продукты. При регуляционной резистентности белок или фермент клетки, инактивирующийся под действием гербицида, начинает усиленно синтезироваться, ликвидируя, таким образом, дефицит нужного метаболита в клетке. Контактный механизм устойчивости обеспечивается изменением структуры мишени (белок или фермент), взаимодействием с которым связано повреждающее действие гербицида Установлено, что признак гербицидоустойчивости является моногенным, то есть признак детерминируется чаще всего одним-единственным геном. Это очень облегчает возможность использования технологии рекомбинантной ДНК для передачи этого признака. Гены, кодирующие те или иные ферменты деструкции и модификации гербицидов, могут быть с успехом использованы для создания гербицидоустойчивых растении методами генетической инженерии. Традиционные методы селекции создания сортов, устойчивых к гербицидам, очень, длительны и малорезультативны. Наиболее широко применяемый за рубежом гербицид глифосат (коммерческое название Roundup) подавляет синтез важнейших ароматических аминокислот, воздействуя на фермент 5-енолпирувилшикимат-3-фосфатсинтазу (ЕПШФ-синтаза). Известные случаи устойчивости к этому гербициду связаны либо с повышением уровня синтеза этого фермента (регуляционный механизм), либо с возникновением мутантного фермента, нечувствительного к глифосфату (контактный механизм). Из устойчивых к глифосфату растений был выделен ген ЕПШФ-синтазы и поставлен под промотор вируса мозаики цветной капусты. С помощью Ti-плазмиды эта генетическая конструкция была введена в клетки петунии. При наличии одной копии гена в регенерированных из трансформированных клеток растениях синтезировалось фермента в 20–40 раз больше, чем в исходных растениях, но устойчивость к глифосфату увеличилась только в 10 раз. К числу наиболее распространенных гербицидов, используемых при обработке зерновых культур, относится атразин. Он подавляет фотосинтез, связываясь с одним из белков фотосистемы II и прекращая транспорт электронов. Устойчивость к гербициду возникает в результате точечных мутаций в этом пластохинон связывающем белке (замена серина на глицин), вследствие чего он теряет способность взаимодействовать с гербицидом. В ряде случаев удалось осуществить перенос гена мутантного белка в чувствительные к атразину растения с помощью Ti-плазмиды. Интегрированный в хромосому растений ген устойчивости был снабжен сигнальной последовательностью, которая обеспечивала транспорт синтезируемого белка в хлоропласты. Химерные растения проявляли значительную устойчивость к таким концентрациям атразина, которые вызывали гибель контрольных растений с геном белка дикого типа. Некоторые растения способны инактивировать атразин путем отщепления остатка хлора ферментом глутатион-3-трансфераза. Этот же фермент инактивирует и другие родственные гербициды триазинового ряда (пропазин, симазин и др.). Существуют растения, естественная устойчивость которых к гербицидам основана на детоксикации. Так, устойчивость растений к хлорсульфурону может быть связана с дезактивацией молекулы гербицида путем его гидроксилирования и последующего гликозилирования введенной гидроксильной группы. Создание растений, устойчивых к патогенам и вредителям Устойчивость растений к тем или иным патогенам чаще всего является сложным мультигенным признаком. Одновременная передача нескольких локусов трудна даже методами генной инженерии, не говоря о классических методах селекции. Более простым является другой путь. Известно, что у устойчивых растений при атаке патогенов изменяется метаболизм. Накапливаются такие соединения, как Н2О, салициловая кислота, фитоаллексины. Повышенный уровень этих соединений способствует противостоянию растения в борьбе с патогенами. Вот один из примеров, доказывающий роль салициловой кислоты в иммунном ответе растений. Трансгенные растения табака, которые содержат бактериальный ген, контролирующий синтез салицилат гидролазы (этот фермент разрушает салициловую кислоту), были неспособны к иммунному ответу. Поэтому изменение генно-инженерным путем уровня салициловой кислоты или выработки в растениях в ответ на патоген Н2О2 может быть перспективным для создания устойчивых трансгенных растений. В фитовирусологии широко известен феномен индуцированной перекрестной устойчивости растений к вирусным инфекциям. Сущность этого явления состоит в том, что заражение растения одним штаммом вируса предотвращает последующую инфекцию этих растений другим вирусным штаммом. Молекулярный механизм подавления вирусной инфекции пока неясен. Показано, что для иммунизации растений достаточно введения отдельных вирусных генов, например генов капсидных белков. Так, ген белка оболочки вируса табачной мозаики перенесли в клетки табака и получили трансгенные растения, у которых 0,1 % всех белков листьев был представлен вирусным белком. Значительная часть этих растений при инфицировании вирусом не проявляла никаких симптомов заболевания. Возможно, что синтезирующийся в клетках белок оболочки вируса мешает вирусной РНК нормально функционировать и формировать полноценные вирусные частицы. Установлено, что экспрессия капсидного белка вируса табачной мозаики, вируса мозаики люцерны, вируса огуречной мозаики, Х-вируса картофеля в соответствующих трансгенных растениях (табак, томаты, картофель, огурцы, перцы) обеспечивает высокий уровень их защиты от последующей вирусной инфекции. Причем у трансформированных растений не отмечалось снижения фертильности, нежелательного изменения ростовых и физиологических характеристик исходных экземпляров и их потомства. Полагают, что индуцированная устойчивость растений к вирусам обусловлена особым антивирусным белком, очень похожим на интерферон животных. Представляется возможным методом генетической инженерии усилить экспрессию гена, кодирующего этот белок, путем его амплификации или подстановки под более сильный промотор. Следует отметить, что использование генетической инженерии для защиты растений от различных патогенных микроорганизмов в значительной мере сдерживается недостаточностью знаний о механизмах защитных реакций растений. Для борьбы с насекомыми-вредителями в растениеводстве используются химические средства — инсектициды. Однако они оказывают вредное влияние на млекопитающих, убивают и полезных насекомых, загрязняют окружающую среду, дороги, и, кроме того, насекомые довольно скоро приспосабливаются к ним. Известно более 400 видов насекомых, устойчивых к используемым инсектицидам. Поэтому все большее внимание привлекают биологические средства борьбы, обеспечивающие строгую избирательность действия и отсутствие адаптации вредителей к применяемому биопестициду. Уже довольно давно известна бактерия Bacillus thuringiensis, продуцирующая белок, являющийся очень токсичным для многих видов насекомых, в то же время безопасный для млекопитающих. Белок (дельта-эндотоксин, CRY-белок) продуцируется различными штаммами В. thuringiensis. Взаимодействие токсина с рецепторами строго специфично, что усложняет подбор комбинации токсин — насекомое. В природе найдено большое количество штаммов В. thuringiensis, чьи токсины дей ствуют только на определенные виды насекомых. Препараты В. thuringiensis в течение десятилетий использовали для контроля насекомых на полях. Безопасность токсина и его составных белков для человека и других млекопитающих полностью доказана. Встраивание гена этого белка в геном растений дает возможность получить трансгенные растения, не поедаемые насекомыми. Кроме видоспецифичности по действию на насекомых встраивание прокариотических генов дельта-токсинов в геном растений даже под контролем сильных эукариотических промоторов не привело к высокому уровню экспрессии. Предположительно такое явление возникло в связи с тем, что эти бактериальные гены содержат значительно больше адениновых и тиминовых нуклеотидных оснований, чем растительная ДНК. Эта проблема была решена путем создания модифицированных генов, где из природного гена вырезали и добавляли те или иные фрагменты с сохранением доменов, кодирующих активные части дельта-токсина. Так, например, с помощью таких подходов был получен картофель, устойчивый к колорадскому жуку. Получены трансгенные растения табака, способные синтезировать токсин. Такие растения были нечувствительны к гусеницам Manduca sexta. Последние погибали в течение 3 суток контакта с токсинпродуцирующими растениями. Токсинообразование и обусловленная им устойчивость к насекомым передавалась по наследству как доминантный признак. В настоящее время так называемые Bt-растения (от В. thuringiensis) хлопка и кукурузы занимают основную долю в общем объеме генетически модифицированных растений этих культур, которые выращивают на полях США. В связи с возможностями генной инженерии конструировать энтомопатогенные растения на основе токсина микробного происхождения еще больший интерес к себе вызывают токсины растительного происхождения. Фитотоксины являются ингибиторами белкового синтеза и осуществляют защитную функцию, направленную против насекомых-вредителей микроорганизмов и вирусов. Лучше всех среди них изучен рицин, синтезируемый в клещевине: его ген клонирован и установлена нуклеотидная последовательность. Однако высокая токсичность рицина для млекопитающих ограничивает генно-инженерные работы с ним только техническими культурами, не используемыми в пищу человека и на корм животным. Токсин, вырабатываемый фитолаккой американской, эффективен против вирусов и безвреден для животных. Механизм его действия заключается в инактивации собственных рибосом при проникновении в клетки различного рода патогенов, в том числе фитовирусов. Пораженные клетки некротизируются, предотвращая размножение патогена и его распространение по растению. В настоящее время проводятся исследования по изучению гена этого белка и передаче его в другие растения. Вирусные болезни широко распространены среди насекомых, поэтому для борьбы с насекомыми-вредителями можно использовать природные вирусы насекомых, препараты которых называют вирусными пестицидами. В отличие от ядохимикатов они обладают узким спектром действия, не убивают полезных насекомых, они быстро разрушаются во внешней среде и не опасны для растений и животных. Наряду с вирусами насекомых используются как биопестициды некоторые грибы, поражающие насекомых-вредителей. Применяемые сейчас биопестициды являются природными штаммами энтомопатогенных вирусов и грибов, однако не исключена возможность создания в будущем методами генетической инженерии новых эффективных биопестицидов.
Повышение устойчивости растений к стрессовым условиям Растения очень часто подвергаются воздействию различных неблагоприятных факторов окружающей среды: высокие и низкие температуры, недостаток влаги, засоление почв и загазованность среды, недостаток или, напротив, избыток некоторых минеральных веществ и т. д. Этих факторов множество, поэтому и способы защиты от них многообразны — от физиологических свойств до структурных приспособлений, позволяющих преодолевать их пагубное действие. Устойчивость растений к тому или иному стрессовому фактору является результатом воздействия множества разных генов, поэтому говорить о полной передаче признаков толерантности от одного вида растения другому генно-инженерными методами не приходится. Тем не менее у генетической инженерии имеются определенные возможности для повышения устойчивости растений. Это касается работы с отдельными генами, контролирующими метаболические ответы растений на стрессовые условия, например сверхпродукцию пролина в ответ на осмотический шок, на действие засоления, синтез особых белков в ответ на тепловой шок и т. д. Дальнейшее углубленное изучение физиологической, биохимической и генетической основы ответной реакции растения на условия среды, несомненно, позволит применять методы генетической инженерии для конструирования устойчивых растений. Пока можно отметить лишь косвенный подход для получения морозоустойчивых растений, основанный на генно-инженерных манипуляциях с Pseudomonas syringae. Этот микроорганизм, сосуществующий с растениями, способствует их повреждению ранними заморозками. Механизм явления связан с тем, что клетки микроорганизма синтезируют особый белок, локализующийся во внешней мембране и являющийся центром кристаллизации льда. Известно, что формирование льда в воде зависит от веществ, могущих служить центрами образования льда. Белок, вызывающий формирование кристаллов льда в различных частях растения (листья, стебли, корни), является одним из главных факторов, ответственных за повреждение тканей растений, чувствительных к ранним заморозкам. Многочисленные эксперименты в строго контролируемых условиях показали, что стерильные растения не повреждались заморозками вплоть до — 6–8 °C, тогда как у растений, имеющих соответствующую микрофлору, повреждения возникали уже при температурах — 1,5–2 °C. Мутанты этих бактерий, потерявшие способность синтезировать белок, вызывающий формирование кристаллов льда, не повышали температуру образования льда, и растения с такой микрофлорой были устойчивы к заморозкам. Штамм таких бактерий, распыленный над клубнями картофеля, конкурировал с обычными бактериями, что приводило к повышению морозоустойчивости растений. Возможно, такие бактерии, созданные с помощью методов генной инженерии и используемые в качестве компонента внешней среды, будут служить для борьбы с заморозками.
Повышение эффективности биологической азотфиксации Хорошо изучен фермент ответственный за восстановление молекулярного азота до аммония — нитрогеназа. Структура нитрогеназы одинакова у всех азотфиксирующих организмов. При фиксации азота непременным физиологическим условием является защита нитрогеназы от разрушения под действием кислорода. Лучше всех среди азотфиксаторов изучены ризобии, образующие симбиоз с бобовыми растениями, и свободноживущая бактерия Klebsiella pneumoniae. Установлено, что у этих бактерий за фиксацию азота ответственно 17 генов — так называемых nif-генов. Все эти гены сцеплены друг с другом и расположены в хромосоме между генами ферментов биосинтеза гистидина и генами, определяющими усвоение шикимовой кислоты. У быстрорастущей ризобии nif-гены существуют в форме мегаплазмиды, содержащей 200–300 тысяч пар нуклеотидов. Среди генов азотфиксации выявлены гены, контролирующие структуру нитрогеназы, белковый фактор, принимающий участие в транспорте электронов, регуляторные гены. Регуляция генов азотфиксации довольно сложна, поэтому генно-инженерный перенос азотфиксирующей функции от бактерий непосредственно высшим растениям в настоящее время уже не обсуждается. Как показали эксперименты, даже в самом простом эукариотическом организме — дрожжах не удалось добиться экспрессии nif-генов, хотя они и сохранялись в течение 50 генераций. Эти опыты показали, что диазотрофность (азот-фиксация) свойственна исключительно прокариотическим организмам, и nif-гены не смогли преодолеть барьер, разделяющий прокариоты и эукариоты, из-за слишком сложной своей структуры и регуляции генами, расположенными вне nif-области. Возможно, более удачным окажется перенос nif-генов с помощью Ti-плазмид в хлоропласты, поскольку механизмы экспрессии генов в хлоропластах и в клетках прокариот близки. В любом случае нитрогеназа должна быть защищена от ингибирующего действия кислорода. Кроме того, фиксация атмосферного азота — очень энергоемкий процесс. Вряд ли растение под влиянием nif-генов может так кардинально изменить свой метаболизм, чтобы создать все эти условия. Хотя не исключено, что в будущем методами генетической инженерии можно будет создать более экономно работающий нитрогеназный комплекс. Более реально использование генноинженерных методов для решения следующих задач: повышение способности ризобии колонизировать бобовые растения, повышение эффективности фиксации и ассимиляции азота путем воздействия на генетический механизм, создание новых азотфиксирующих микроорганизмов путем введения в них nif-генов, передача способности к симбиозу от бобовых растений к другим. Первостепенной задачей генетической инженерии для повышения эффективности биологической фиксации азота является создание штаммов ризобии с усиленной азотфиксацией и колонизирующей способностью. Колонизация бобовых растений ризобиями протекает очень медленно, лишь единичные из них дают начало клубенькам. Это происходит потому, что местом инвазии ризобии является только одна небольшая область между точкой роста корня и ближайшим к ней корневым волоском, находящимся на стадии формирования. Все остальные части корня и развившиеся корневые волоски растения нечувствительны к колонизации. В ряде случаев сформировавшиеся клубеньки оказываются неспособными фиксировать азот, что зависит от многих растительных генов (выявлено не менее пяти), в частности от неблагоприятного сочетания двух рецессивных генов. Традиционными методами генетики и селекции удалось получить лабораторные штаммы ризобий с более высокой колонизирующей способностью. Но они в полевых условиях испытывают конкуренцию со стороны местных штаммов. Повышение их конкурентоспособности, видимо, можно осуществить генноинженерными методами. Повышение эффективности процесса азотфиксации возможно применением генноинженерных приемов, основанных на увеличении копий гена, усилении транскрипции тех генов, продукты которых образуют «узкое» место в каскадном механизме азотфиксации, путем введения более сильных промоторов и т. п. Важно повышение коэффициента полезного действия самой нитрогеназной системы, осуществляющей непосредственное восстановление молекулярного азота в аммиак.
Повышение эффективности фотосинтеза С4-растения характеризуются высокими темпами роста и скоростью фотосинтеза, у них практически отсутствует видимое фотодыхание. У большинства сельскохозяйственных культур, относящихся к С3-растениям, высокая интенсивность фотодыхания. Фотосинтез и фотодыхание — тесно связанные процессы, в основе которых лежит бифункциональная активность одного и того же ключевого фермента — рибулозобисфосфат-карбоксилазы (РуБФК). РуБФ-карбоксилаза может присоединять не только СО2, но и О2, то есть осуществляет реакции карбоксилирования и оксигенирования. При оксигенировании РуБФ образуется фосфогликолат, который служит основным субстратом фотодыхания — процесса выброса СО2 на свету, в результате чего теряется часть фотосинтетических продуктов. Низкое фотодыхание у С4-растений объясняется не отсутствием ферментов гликолатного пути, а ограничением оксигеназной реакции, а также реассимиляцией СО2 фотодыхания. Одной из задач, стоящих перед генетической инженерией, является исследование возможности создания РуБФК с преобладающей карбоксилазной активностью.
Получение растений с новыми свойствами В последние годы ученые используют новый подход для получения трансгенных растений с "antisense RNA" (перевернутой или антисмысловой РНК), который позволяет управлять работой интересуемого гена. В этом случае при конструировании вектора копию ДНК (к-ДНК) встраиваемого гена переворачивают на 180°. В результате в трансгенном растении образуется нормальная молекула мРНК и перевернутая, которая в силу комплементарности нормальной мРНК образует с ней комплекс и закодированный белок не синтезируется. Такой подход использован для получения трансгенных растений томатов с улучшенным качеством плодов. Вектор включал к-ДНК гена PG, контролирующего синтез полигалактуроназы — фермента, участвующего в разрушении пектина, основного компонента межклеточного пространства растительных тканей. Продукт гена PG синтезируется в период созревания плодов томатов, а увеличение его количества приводит к тому, что томаты становятся более мягкими, что значительно сокращает срок их хранения. Отключение этого гена в трансгенах позволило получить растения томатов с новыми свойствами плодов, которые не только значительно дольше сохранялись, но и сами растения были более устойчивы к грибным заболеваниям. Такой же подход можно применить для регулирования сроков созревания томатов, а в качестве мишени в этом случае используют ген EFE (ethylene-forming enzyme), продуктом которого является фермент, участвующий в биосинтезе этилена. Этилен — это газообразный гормон[76], одной из функций которого является контроль за процессом созревания плодов. Стратегия антисмысловых конструкций широко применима для модификации экспрессии генов. Эта стратегия используется не только для получения растений с новыми качествами, но и для фундаментальных исследований в генетике растений. Следует упомянуть еще об одном направлении в генной инженерии растений, которое до недавнего времени в основном использовали в фундаментальных исследованиях — для изучения роли гормонов в развитии растений. Суть экспериментов заключалась в получении трансгенных растений с комбинацией определенных бактериальных гормональных генов, например только iaaM или ipt т. д. Эти эксперименты внесли существенный вклад в доказательство роли ауксинов и цитокининов в дифференцировке растений. В последние годы этот подход стали использовать в практической селекции. Оказалось, что плоды трансгенных растений с геном iaaM, находящимся под промотором гена Def (ген, который экспрессируется только в плодах), являются партенокарпическими, то есть сформировавшимися без опыления. Партенокарпические плоды характеризуются либо полным отсутствием семян, либо очень небольшим их количеством, что позволяет решить проблему "лишних косточек", например в арбузе, цитрусовых и т. д. Уже получены трансгенные растения кабачков, которые в целом не отличаются от контрольных, но практически не содержат семян. Обезоруженную, лишенную онкогенов Ti-плазмиду ученые активно используют для получения мутаций. Этот метод носит название Т-ДНК-инсерционного мутагенеза. Т-ДНК, встраиваясь в геном растения, выключает ген, в который она встроилась, а по утрате функции можно легко отбирать мутанты (явление сайлесинга — замолкания генов). Этот метод замечателен также тем, что позволяет сразу обнаружить и клонировать соответствующий ген. В настоящее время таким способом получено множество новых мутаций растений и соответствующие гены клонированы. М. А. Раменской на основе Т-ДНК мутагенеза получены растения томатов с неспецифической устойчивостью к фитофторозу. Не менее интересен и другой аспект работ — получены трансгенные растения с измененными декоративными свойствами. Один из примеров — это получение растений петунии с разноцветными цветками. На очереди голубые розы с геном, контролирующим синтез голубого пигмента, клонированным из дельфиниума.
Проблемы биобезопасности трансгенных растений Одним из главных возражений против употребления "трансгенных" продуктов питания является наличие во многих из них генов устойчивости к антибиотику (в частности, к канамицину), которые содержались в исходной конструкции ДНК в качестве селективных. Предполагается, что эти гены устойчивости могут при переваривании пищи передаваться эндогенной микрофлоре, в том числе патогенной, в результате чего микробы могут приобрести резистентность к данному антибиотику. Однако в реальности вероятность такого события ничтожно мала — многочисленные эксперименты и наблюдения в природе относительно подобного горизонтального переноса генов до сих пор давали только отрицательные результаты. Не стоит забывать, что встраиваемые в растения гены устойчивости "настроены" для экспрессии лишь в эукариотических, но не бактериальных клетках. Надо учесть и то, что эти селективные гены взяты из природных популяций микроорганизмов, где они сейчас широко распространены в результате активного применения антибиотиков в медицинской практике. Поэтому вероятность попадания гена устойчивости к антибиотику в микрофлору человека из природного резервуара несравнимо реальнее, чем при употреблении трансгенных растений. Однако, учитывая настроения общественности, разрабатываются подходы, для исключения присутствия "подозрительных" генов в коммерциализированных трансгенных формах. В большинстве случаев маркерные гены устойчивости к антибиотикам сейчас заменяют на гены устойчивости к гербицидам. Правда, применение "гербицидных" генов также встречает возражения, но уже защитников окружающей среды. Предложено несколько способов избирательной элиминации маркерного гена после получения желаемого трансгенного растения, когда он фактически уже не нужен. Очень перспективным представляется замена селективных генов на репортерные при отборе трансгенных форм растений, либо использование альтернативных селективных генов, таких как гены синтеза фитогормонов или гидролиза особых форм полисахаридов при выращивании растений в культуральной среде. Таким образом, даже эта виртуальная опасность, связанная с генами устойчивости к антибиотику, в скором времени перестанет существовать. Что касается возможной токсичности или аллергенности трансгенных растений, то здесь применяют те же жесткие стандарты, как и для полученных традиционным путем новых сортов культурных растений или новых видов продуктов питания. Никаких особых отличий трансгенных растений от обычных по этим параметрам ожидать не приходится (разве что в лучшую сторону при блокировании синтеза токсинов или аллергенов), да и действительно, как правило, не наблюдается на практике. Проблема возможного ущерба для окружающей среды имеет несколько аспектов. Во-первых, существует опасение, что устойчивые к гербицидам культурные растения могут при межвидовом опылении передавать эти гены близкородственным сорнякам, которые могут превратиться в неистребимые суперсорняки (superweeds). Хотя вероятность такого нежелательного развития событий для большинства сельскохозяйственныхкультур очень мала, генные инженеры и ученые-аграрии активно разрабатывают подходы для исключения подобной опасности. Здесь, правда, надо отметить, что данный вопрос также не нов, так как в практике сельского хозяйства уже давно используется ряд устойчивых к гербицидам сортов, полученных путем обычной селекции. При этом никакой экологической катастрофы широкое использование таких устойчивых сортов до сих пор не вызвало[77]. Тем не менее, и в этом случае, чтобы отвести любые возражения от трансгенных растений, пробуют, например, вводить в растения не один, а сразу несколько генов устойчивости к разным гербицидам. Передача нескольких генов сорнякам гораздо менее вероятна, чем одного гена. Кроме того, мультигербицидная устойчивость позволит чередовать разные гербициды при обработке посевов, что не даст возможности для распространения какого-либо определенного гена устойчивости в сорняках. Предлагается также вводить гены устойчивости не в ядерный, а в хлоропластный геном. Это может предотвратить нежелательный дрейф генов с помощью пыльцы, так как хлоропласты наследуются только по материнской линии. Еще один генно-инженерный путь борьбы с сорняками без использования генов резистентности к гербицидам вообще — биотрансгенный. Речь идет об использовании мелких животных, например, кроликов, для поедания сорняков на полях. При этом чтобы оградить от поедания культурные растения, в них можно ввести какой-либо ген, делающий их непривлекательными (запах, вкус) для данного животного. Такой биотрансгенный подход сразу снял бы большинство выдвигаемых сейчас возражений против трансгенных культур. Близкие по сути экологические возражения касаются трансгенных растений со встроенными "инсектицидными" генами, способных, как считают, спровоцировать у насекомых-вредителей возникновение массовой резистентности. Здесь также предложены действенные способы для уменьшения этой опасности, например, использование генов нескольких разных токсинов и/или индуцибельных промоторов, быстро активирующихся при нападении насекомых на растение. Данная проблема, в общем, не нова, так как многие из инсектицидов, используемых сейчас на "генном уровне", давно применяют в виде чистого вещества для опрыскивания посевов. Еще одно нежелательное следствие использования трансгенных растений с генами инсектицидов заключается в том, что пыльца этих растений может быть токсичной и для полезных насекомых, которые данной пыльцой питаются. Некоторые экспериментальные данные говорят о том, что такая опасность действительно существует, хотя о ее возможных масштабах говорить пока трудно. Однако и здесь уже предложены и испытаны адекватные генно-инженерные решения, например, использование трансгеноза через хлоропластную ДНК, или промоторов, не работающих в пыльце.
ЛИТЕРАТУРА К РАЗДЕЛУ «ГЕННАЯ ИНЖЕНЕРИЯ»
1. Алберте Б., Брей Д., Льюис Дж. и др. Молекулярная биология клетки. Т. 1–3. М.: Мир, 1994. 2. Анализ генома. Методы / Под ред. К. Дейвиса. М.: Мир, 1990. 246 с. 3. Атанасов А. Биотехнология в растениеводстве. Новосибирск: ИЦиГСО РАН, 1993. - 241 с. 4. Барановов В. С. Генная терапия — медицина XXI века // Соросовский образовательный журнал. № 3. 1999. С. 3 — 68. 5. Бекер М. Е., Лиепиньш Г.К., Райпулис Е.П. Биотехнология. М.: Агропромиздат, 1990. 334 с. 6. Борисюк Н.В. Молекулярно — генетическая конституция соматических гибридов // Биотехнология. Итоги науки и техники ВИНИТИ АН СССР. М., 1988. Т. 9. С. 73 -113. 7. Валиханова Г. Ж. Биотехнология растений. Алматы: Конжык, 1996. 272 с. 8. Глеба Ю. Ю. Биотехнология растений // Соросовский образовательный журнал. № 6. 1998. С. 3–8. 9. Глебов О. К. Генетическая трансформация соматических клеток // Методы культивирования клеток. Л.: Наука, 1988. 10. Гольдман И. Л., Разин С. В., Эрнст Л. К., Кадулин С. Г., Гращук М. А. Молекулярно-биологические аспекты проблемы позиционно-независимой экспрессии чужеродных генов в клетках трансгенных животных // Биотехнология. 1994. № 2. 11. Дыбан А. П., Городецкий С. И. Интродукция в геном млекопитающих чужеродных генов: пути и перспективы // Молекулярные и клеточные аспекты биотехнологии. Л.: Наука, 1986. С. 82–97. 12. Егоров Н. С., Самуилов В. Д. Современные методы создания промышленных штаммов микроорганизмов // Биотехнология. Кн. 2. М.: Высшая школа, 1988. 208 с. 13. Зверева С. Д., Романов Г. А. Репортерные гены для генетической инженерии растений: хара-пктеристика и методы тестирования // Физиология растений. 2000. Т. 47, № 3. С. 479–488. 14. Лещинская И. Б. Генетическая инженерия // Соросовский образовательный журнал. 1996. № 1. С. 33–39. 15. Ли А., Тинланд Б. Интеграция т-ДНК в геном растений: прототип и реальность // Физиология растений. 2000, том 47, № 3. С. 354–359 16. Лутова Л. А., Проворов Н. А., Тиходеев О. Н. и др. Генетика развития растений. СПб.: Наука, 200. 539 с. 17. Льюин Б. Гены. М.: Мир, 1987. 544 с. 18. Пирузян Э. С., Андрианов В. М. Плазмиды агробактерий и генная инженерия растений. М.: Наука, 1985. 280 с. 19. Пирузян Э. С. Генетическая инженерия растений. М.: Знание, 1988. 64 с. 20. Пирузян Э. С. Основы генетической инженерии растений. М.: Наука, 1988. 304 с. 21. Пирузян Э. С. Проблемы экспрессии чужеродных генов в растениях // Итоги науки и техники ВИНИТИ. Сер. Биотехнология. 1990. Т. 23. 176 с. 22. Попов Л. С., Языков А. А. Трансгенные животные как модели для изучения репродукции эмбрионального развития и заболеваний человека // Успехи современной биологии.1999. Т 119, № 1. С. 30–41. 23. Романов Г. А. Генетическая инженерия растении и пути решения проблемы биобезопасности // Физиология растений, 2000. Том 47, № 3. С. 343–353 24. Сельскохозяйственная биотехнология: Учеб. /В. С. Шевелуха, Е. А. Калашникова, С. В. Дегтярев и др.: Под ред. В. С. Шевелухи. М.: Высш. школа, 1998. 416 с. 25. Сингер М., Берг П. Гены и геномы. Т. 1–2. М.: Мир, 1998. 26. Томилин Н. В., Глебов О. К. Генетическая трансформация клеток млекопитающих // Молекулярные и клеточные аспекты биотехнологии. Л.: Наука, 1986. С. 62–82. 27. Фаворова О. О. Лечение генами — фантастика или реальность? // Соросовский образовательный журнал. № 2. 1997. С. 21–27. 28. Щелкунов С. Н. Генетическая инженерия. Ч. 1. Новосибирск: Изд-во Новосибирского ун-та, 1994. 304 с.
ЛАБОРАТОРИЯ
Практическая биотехнология[78] для начинающих
Кузьмина Н.А.
Википедия Биотехнология — это интеграция естественных и инженерных наук, позволяющая наиболее полно реализовать возможности живых организмов или их производные для создания и модификации продуктов или процессов различного назначения. Чаще всего применяется в медицине, пищевой промышленности, также для решение проблем в области энергетики, охране окружающей среды, и в научных исследованиях. В последние десятилетия биология бурно развивается и создаёт новые научные направления. Новое комплексное направление — физико-химическая биология, включающая в себя биохимию, биофизику, молекулярные биологию и генетику, биоорганическую химию и некоторые другие дисциплины, — не только помогает решать задачи, которые давно ставила перед биологией производственно-техническая практика, но и намечает пути принципиально нового биологического производства. В результате стремительного прогресса разных составных частей физико-химической биологии, возникло новое направление в науке и производстве, получившее наименование биотехнологии. Это направление сформировалось за последние два десятка лет и уже сейчас получило мощное развитие. Особенно интенсивно биотехнология стала развиваться с 1981 года. Задачи физико-химической биологии очень обширны. Объединяет их то, что основу, суть каждой задачи составляет познание природы живого и использование в практике знаний о процессах и материальных структурах живых организмов. Стремительно расширяющиеся знания о процессах жизнедеятельности позволяют не только приспосабливать эти процессы для практических целей, но и управлять ими, а также создавать весьма перспективные в практическом отношении новые системы, не существующие в природе, хотя и аналогичные существующим. Биотехнология в целом представляет собой систему приёмов направленного использования процессов жизнедеятельности живых организмов для получения промышленным способом ценных продуктов.
Меня часто спрашивали — и учителя, и школьники — нельзя ли завести какие-либо простенькие культуры, иллюстрирующие биотехнологические процессы для использования на уроках? Можно, и в этом нет ничего суперсложного. Знаменитому физику Резерфорду, который и сам ставил блестящие эксперименты, имея порой в распоряжении простое оборудование, приписывают высказывание о том, что настоящий ученый сможет провести эксперимент при помощи палки и веревки. Ну, например, для стерилизации сред нужен автоклав. Чем его можно заменить? Очень просто — скороварка. А если вы найдете скороварку с 2 режимами работы — на 1 и 1,5 атмосферы — будет вообще замечательно. Ах, скажете вы — а где мы возьмем агар-агар[79]? Можно заменить его крахмалом или желатином. Конечно, придется немного поэкспериментировать с концентрацией и режимами стерилизации, но в конце концов, уверена, вы подберете себе что-нибудь подходящее. Да, состав пищевого желатина и пищевого крахмала далек от химически чистого, но для простых экспериментов сойдут и эти пищевые продукты. Желатин не переносит стерилизацию под давлением? Попробуйте прием "дробной стерилизации" — 3 раза с интервалом в день по полчаса обрабатывайте пробирки со средой паром в скороварке. Обычно для убивания вегетативных форм микроорганизмов этого достаточно. Если у вас нет ни агара, ни желатина, ни крахмала — культивируйте на плотиках из фильтровальной бумаги. Когда мы проводили работы по селекции каллусов на среде с ПЭГ (очень сильный осмотик, имитирующий засуху в этих экспериментах), то столкнулись с проблемой — агар в присутствии высоких концентраций полиэтиленгликоля не застывал после автоклавирования. Выход был найден — на дно пробирок или баночек клали комочек ваты (рыхлый), сверху — вырезанный по диаметру сосуда кусочек фильтровальной бумаги, заливали жидкой питательной средой так, чтобы ее уровень не превышал 1 мм над поверхностью бумаги. Высота столбика среды в пробирке диаметром 2 см — примерно 1–1,5 см, в баночке из под детского питания — 0,5 см. В качестве мерной посуды можно использовать то, что продается в хозяйственных магазинах для домовитых хозяек — мерные стаканы. Для более мелких доз — в аптеке вы найдете шприцы в ассортименте, которые прекрасно заменят вам мерные пробирки и пипетки. Ультрафиолетовые лампы сейчас можно купить в хозяйственном магазине. Там же вы найдете хлорный отбеливатель для стерилизации экспланта. Сухожаровой шкаф с успехом заменит обычная духовка газовой или электрической плиты.
Правила стерильной работы в лаборатории Выращивание изолированных клеток, тканей, органов, растений-регенерантов, водных культур и грибов, используемых в биотехнологии, проводят в условиях полной асептики, т. е. стерильно. Особое внимание следует обратить на чистоту посуды, предназначенной для приготовления питательных сред и их компонентов; на подготовку объектов к пересадке, пассированию и культивированию. Только некоторые объекты (хлорелла, азолла) можно выращивать в нестерильных условиях.
Приемы и методы стерилизации Стерилизация — полное уничтожение микроорганизмов и их покоящихся форм (например, спор). Существуют разные методы стерилизации: с помощью влажного пара, сухого пара, облучения ультрафиолетовыми лучами, обработки химическими веществами и микрофильтрации. Обработка влажным паром производится в автоклавах. Вегетативные клетки бактерий и грибов гибнут через 5-10 минут уже при температуре около 60 °C; для гибели спор дрожжей и грибов требуется температура 120 °C в течение 15 минут. Продолжительность автоклавирования зависит от величины (теплоемкости) пробирок, колб и объема питательной среды в них. Иногда автоклавируют несколько раз — дробная стерилизация. Этот прием используют для стерилизации, как питательных сред, так и посуды.
Стерилизация посуды. Большинство культур в лабораторных условиях выращивают в пробирках, колбах Эрленмейера различного объема и чашках Петри одно- или многоразового использования. Вначале посуду тщательно моют с использованием детергентов, а также раствора двухромовокислого калия в серной кислоте (хромпика[80]). Вымытую посуду ополаскивают водопроводной, затем дистиллированной водой и высушивают в сушильном шкафу. Чтобы избежать заражения стерильных предметов из воздуха, перед стерилизацией их закрывают ватными пробками, заворачивают в оберточную бумагу или закрывают фольгой (у стаканов, колб достаточно завернуть только горлышко). Затем посуду можно стерилизовать двумя способами: 1. Посуду выдерживают в автоклаве под давлением в течение 20–40 минут при температуре 100–130 °C. Продолжительность автоклавирования зависит от его режима: при давлении 0.5 атмосферы — 20–40 минут, при 1 атм. — 15 минут. 2. При сухом способе стерилизации чашки Петри, колбы, стаканы, завернутые в плотную бумагу, стерилизуют в сушильном шкафу при температуре 140 °C в течение 2 часов, при температуре 180 °C — 30 минут. При более высоких температурах ватные пробки буреют, а бумага становится ломкой.
Стерилизация инструментов. Инструменты (скальпели, пинцеты, иглы и т. д.) стерилизуют в сушильном шкафу способом № 2. Шприцы, ножницы, пробочные сверла удобнее кипятить. Металлические предметы нельзя автоклавировать: под воздействием пара они ржавеют и тупятся. Непосредственно перед работой и в процессе её инструменты помещают в стакан со спиртом и обжигают в пламени спиртовки. Стерильный инструмент используют только для одноразовой манипуляции! Перед повторным употреблением его снова окунают в спирт и обжигают.
Стерилизация материалов. Вату, марлю, ватные пробки, фильтровальную бумагу, халаты, косынки стерилизуют в автоклаве под давлением 2 атм. в течение 25–30 мин.
Стерилизация питательных сред. Автоклавирование питательных сред для выращивания культур тканей проводят после их разлива в пробирки или колбы под давлением 0.7–0.8 атм. при температуре 115–120 °C в течение 15–30 минут, в зависимости от объема среды. Если в результате стерилизации среда помутнела, следовательно, неправильно выбран режим стерилизации[81].
Холодная стерилизация. Органические жидкости, не выносящие нагревания, освобождаются от бактерий при пропускании через стерильные мелкопористые бактериальные фильтры с диаметром пор 0.45 мкм.
Как изготовить микробиологическую пробку

Для предотвращения заражения культур из воздуха и их преждевременного высыхания пробирки и колбы закрывают пробками. Пробки для посуды условно можно разделить на ватные (традиционные микробиологические) и крышечки из фольги. С ватными пробками проще работать начинающим — меньше риска получить ожог. Пробки должны плотно входить в пробирку (колбу) на 2–3 см, или на 2/3 своей длины; при открывании хорошо закрытой посуды раздается характерный хлопок. Недостатки микробиологических пробок — довольно трудоемки в изготовлении, часто прожигаются, не очень удобны при долговременном культивировании. Если для микробиологических культур сроки культивирования редко превышают 2 недели, то для растительных клеток продолжительность субкультивирования — как минимум 3 недели. За это время среда начинает подсыхать. Поэтому в таких случаях важно закрывать ватную пробку сверху герметично парафильмом или пищевой пленкой. Крышечка из фольги проще в работе — легко изготавливается (вырезается кружочек или квадратик по диаметру горлышка пробирки или колбы + один — полтора сантиметра для отгибания вниз). Фольгу нужно брать плотную. Если нет плотной — тонкая фольга складывается в 3–4 раза. При обжигании краешков такой пробки нужно быть аккуратным, чтобы не получить ожог, так как фольга металлическая и хорошо проводит тепло. После одевания пробки самый краешек, загнутый вниз на пробирку или колбу, оборачивается узенькой полоской парафильма или пищевой пленки в 2–3 оборота, таким образом краешки плотно прилегают к посуде, и крышка держится надежнее, герметичнее закрывает, снижается риск контаминации (занесения посторонней микрофлоры)

Изготовление микробиологической пробки Сложить вдвое кусок марли, накрыть им верхнюю часть пробирки. На середину куска, закрывающего отверстие пробирки, положить вату, протолкнуть в пробирку на глубину примерно 3–4 см. Вату очень плотно утрамбовать, затем концы марли скрутить в центре, обвязать ниткой, лишнее обрезать. Обрезки марли можно использовать вместе с ватой при изготовлении пробок. Проверить качество пробок: хлопок в момент открывания пробирки, очень твердая ("каменная") на ощупь. Для колб пробки готовят следующим образом: прямоугольный слой ваты скатать в виде очень плотного, до твердости валика нужных размеров, учитывая диаметр и длину горла колбы, обернуть его двойным марлевым слоем. Не использовать пробки, изготовленные только из ваты — они легко воспламеняются при обжигании!
Работа в ламинарном боксе Все операции, связанные с разливом питательных сред, пересадкой каллусов, тканей, микроорганизмов, ведут в специальных комнатах или ламинарных боксах, где обеспечиваются стерильные условия работы. Ламинарный бокс (ламинар) приспособление для работы в стерильных условиях. Асептические условия в ламинаре создаются с помощью тока воздуха. Ламинарное движение воздуха — движение, при котором струйки воздуха двигаются параллельно, обтекая препятствие равномерными слоями. Ток воздуха, проходя через ламинар, движется к исследователю[82], что позволяет освобождать внутреннее пространство ламинара от спор микроорганизмов.
Подготовка бокса к работе В специальных комнатах (микробиологических боксах) проводят влажную уборку с дезинфицирующими агентами. Для надежности стерилизации перед началом работы помещение лаборатории и внутреннее пространство ламинара облучают УФ-лучами. Облучение ультрафиолетовыми лучами (260 нанометров) — наиболее часто используется в лабораториях для стерилизации помещений, настольных боксов. При длительном воздействии эти лучи вызывают гибель всех бактерий. Бактерии погибают очень быстро, а споры грибов значительно медленнее. Поэтому в боксах устанавливают бактерицидные лампы БУФ-15 или БУФ-30, которые включаются на 30 минут за 1 час до работы. Кроме того, рекомендуется проводить профилактическое облучение боксов в течение 2 часов.

Непосредственно перед работой необходимо протереть настольный бокс или внутренние поверхности ламинара этиловым спиртом[83], разложить в нем необходимые инструменты и материалы: спирт в закрытой посуде, спиртовку (горелку), спички, простерилизованный инструмент и посуду.
Рабочее место при отсутствии ламинара Наличие ламинара — это отлично, просто замечательно. Но что делать, если его нет? Или появится в обозримо-необозримом будущем, а культуры клеток заводить нужно уже сейчас? Шаг первый — постарайтесь найти малопосещаемое людскими массами помещение — чем меньше посетителей — тем меньше источников загрязнения. Шаг второй — всегда перед работой проводите влажную уборку всей комнаты и стерилизуйте ее кварцевой (ультрафиолетовой) лампой. А теперь, собственно — о рабочем месте. В нашем случае оно выглядело примерно так: стол, покрытый ламинатом. Лучше — железный, но сойдет и покрашенный масляной краской. В идеале — поверхность стола должна быть негорючей. Именно поэтому железный стол — лучше всего. Ламинат тоже загорается не сразу, хуже всего в этом отношении масляная краска, а еще пуще всякие дерматины и сукно. Они противопоказаны категорически. Прямо перед вами на столе должны лежать следующие предметы: кафельная плитка или кусок стекла размером 15x15 см или чуть больше. Кафельная плитка — лучше, она выдерживает нагрев, а стекло может лопнуть. Спиртовка, стаканчик с 96 % спиртом (или 200–300 граммовая высокая баночка из-под кофе, детского питания, фруктовых пресервов), широкая кисточка для рисования, мокрая марлевая салфетка или тряпочка размером 30х30, свернутая в несколько раз (смочена водой!), пинцет, скальпель, пробирки/баночки с питательной средой, пробирки/чашки Петри с материалом для размножения.
Порядок работы Протираете влажной тряпочкой поверхность стола, ополаскиваете тряпочку и выжимаете ее. Кладете свернутую под ПРАВУЮ руку, справа от кафельной плитки. Располагаете подручные средства. Кафельная плитка лежит прямо перед вами, в центре рабочего места. За кафельной плиткой по центру стоит спиртовка. Рядом с ней, на расстоянии 10 см или чуть больше стоит стаканчик со спиртом, получается, он будет находиться сразу за тряпочкой справа от вас. Слева от плитки располагаются пробирки со средой и материал, который вы будете вводить в культуру или пассировать. Расстояние от плитки до среды и от плитки до тряпочки не менее 10 см влево и вправо, соответственно. Скальпель и пинцет погружены в спирт примерно на 2/3 своей длины (поэтому важно, чтобы баночка была узкой и высокой и была почто до верха наполнена спиртом. Кисточка также стоит в стаканчике со спиртом. Берете кисточку, мажете спиртом поверхность плитки. Ставите кисточку в спирт. Зажигаете спиртовку, вынимаете скальпель, поджигаете в пламени спиртовки и поджигаете горящим скальпелем спирт на поверхности плитки. Кладете скальпель справа на кафельную плитку (она может еще продолжать гореть) так, чтобы острие скальпеля смотрело вверх (то есть на ребро плитки), но так, чтобы большая часть скальпеля, включая острие, находилось именно над плиткой, в пламени. Правой рукой берете пинцет из баночки со спиртом, перекладываете в левую руку, поджигаете от спиртовки и кладете слева на плитку — аналогично скальпелю. ВНИМАНИЕ — если вы проводите эти процедуры впервые — отрабатывайте навыки, окуная инструменты в спирт на четверть длины и не пугайтесь пламени, оно исчезает в течение минуты. Влажная тряпочка — ваша страховка на случай, если вы подожжете что-то не то, например, запаникуете и бросите горящий инструмент в баночку со спиртом. Тогда сразу накрывайте влажной тряпкой источник огня, без доступа кислорода огонь тухнет моментально. В результате этих манипуляций вы имеете абсолютно стерильную поверхность плитки со стерильными инструментами на ней. Постарайтесь контролировать свои движения и доставать экспланты, открывать пробирки не над плиткой непосредственно, а в зоне между плиткой и горящей спиртовкой, то есть за плиткой. На стерильной плитке вы можете резать эксплант, класть его пинцетом на кончик скальпеля для последующего переноса на питательную среду и она должна оставаться стерильной. Старайтесь не трясти над ней рукавами халата. Использованные инструменты снова погружаете в спирт. Перед каждым новым эксплатном все операции по стерилизации, начиная с обжигания поверхности плитки, повторяются. Помните, что скальпель вы держите указательным и большим пальцем (он лежит на указательном и среднем, зажатый большим пальцем, а крышки с пробирок снимаете мизинцем, предварительно пронеся горлышко пробирки через пламя спиртовки.
Стерилизация растительного экспланта Стерильная культура — культура, свободная от эпифитных и ризосферных микроорганизмов. Работающий должен вымыть руки с мылом и протереть их спиртом, надеть стерильный халат, завязать волосы стерильной косынкой. Перед стерилизацией объекта его тщательно моют теплой водой с мылом, промывают дистиллированной водой, очищают от излишних тканей, снимают кожуру у корнеплодов и корней, кору у побегов, поверхностные листья у верхушек побегов, промывают дистиллированной водой и помещают на несколько секунд в 70 % спирт (семена на 1–2 минуты). После этого сегменты корней, побегов, стеблей, клубней или семена переносят в стерилизующий раствор. Вид стерилизующего агента, его концентрация и время действия, зависящие от особенностей тканей исходных растений, необходимо подобрать таким образом, чтобы убить микроорганизмы и не повредить ткани экспланта. Для поверхностной стерилизации растительных объектов применяют следующие средства стерилизации: сулему (двуххлористая ртуть) (0.1 %), хлорамин (2-10 %), гипохлорит кальция (7-10 % Са(ClO)2) или натрия (NaOCl), перекись водорода (13–18 %) и др. Хлорамин можно купить в аптеке. Для стерилизации можно использовать также хлорсодержащие растворы отбеливателей из хозяйственных магазинов, например, средство "Белизна". Свежие растворы отбеливателей при стерилизации нежных эксплантов, таких как молодые листочки, нужно разводить в 2 или даже в 3 раза. Эффективность стерилизации возрастает при добавлении нескольких капель твина-80 и твина-20 на литр стерилизующего раствора. Продолжительность стерилизации меристем сулемой — 5 минут, семян — 15–20 минут, пергидролем соответственно 15 и 30 минут. После стерилизации материал переносят в стерильную дистиллированную воду, выдерживают 10 минут, затем меняют воду ещё два раза, выдерживая в каждой порции по 15–20 минут. В стерильных чашках Петри или на стерильных листах бумаги, или на обожженной кафельной плитке обрезают стерильным скальпелем концы сегментов исходного материала, где клетки могут быть повреждены, и из средних зон нарезают кусочки тканей, которые высаживают на инициальную среду для образования каллусов. При стерилизации отрезков стебля или верхушечных почек в растворах сулемы или гипохлорита рекомендуется парафинировать срезы, чтобы стерилизатор не проник в сосуды, что может привести к интоксикации ткани.
Ход работы Простерилизовать инструменты, посуду и питательные среды, необходимые для проведения работ по выращиванию стерильных проростков и выращивания каллусных тканей. 1. Завернуть в плотную бумагу и простерилизовать сухим жаром в сушильном шкафу стеклянную посуду — колбы, чашки Петри. 2. Подвергнуть предварительной стерилизации в сушильном шкафу скальпели и пинцеты. Перед помещением в сушильный шкаф завернуть инструменты в плотную бумагу. 3. Простерилизовать в автоклаве дистиллированную воду в колбе. Для получения стерильной воды налейте в колбу 1/3 часть объема дистиллированной воды, закройте ватной пробкой, а сверху плотной бумагой или целлофаном, можно обернуть горлышко фольгой. Автоклавировать 30 минут при 1 атмосфере. 4. Простерилизовать в автоклаве в течение 20 мин при давлении 1 атм. пробирки с питательной средой, закрытые фольгой или ватными пробками, обернутыми плотной бумагой или целлофаном.
Питательные среды для растительных биотехнологических объектов Успех в культивировании объектов зависит от правильного выбора питательной среды и тщательности её приготовления. В состав питательных сред входят макро- и микроэлементы (N, Р, К, Са, S, Mg, Fe, В, Zn, Сu, Со, Mn, J, Мо); витамины B1, В6, В12, РР и другие; углеводы (сахароза, глюкоза, маннит); фитогормоны (чаще всего цитокинины и ауксины в определенном соотношении). Ауксины вызывают клеточную дедифференцировку, цитокинины индуцируют деление дедифференцированных клеток и необходимы для получения каллусных тканей. На средах без гормонов растут "привыкшие" и опухолевые ткани. Для получения стеблевого морфогенеза снижают содержание ауксинов. Из ауксинов чаще всего применяют 2,4-дихлорфеноксиуксусную кислоту (2,4-Д) — 0.1-10 мг/л, нафтилуксусную кислоту (НУК) — 0.1–2 мг/л, ИУК — 1-30 мг/л. Для индукции каллусогенеза используют более высокие концентрации ауксинов, в дальнейшем ткань может расти при более низком содержании ауксинов. В качестве цитокининов используют кинетин, 6-бензиламинопурин (ЕАП), зеатин (0.001-10 мг/л); из них кинетин наименее активен. В состав некоторых сред входит аденин. Иногда используют гибберелловую кислоту (ГК). В качестве ростактиваторов применяют также кокосовое молоко, дрожжевой экстракт, гидролизат казеина и др. Состав питательных сред для культивирования биотехнологических объектов зависит от типа их питания: — среда без глюкозы — для хлореллы, цианобактерий; — среда без витаминов и гормонов — для грибов (фузариума, ботритиса) и для ряски (Lemna); — среда с азотом, сахарами, витаминами и гормонами — для культивирования клеток и тканей высших растений. Разработано много питательных сред, но большинство из них представляют модификации основных: Мурасиге-Скуга (МС), Уайта, Шенка-Хильдебрандта, Гамборга (В5), Линсмайера-Скуга, Хеллера, Чапека и др. Составы питательных сред, получивших наибольшее распространение, приведены в справочниках по физиологии растений и биотехнологии. Ниже вы найдете прописи для базовых сред для культивирования растительных клеток: среда Мурасиге-Скуга и среда Гамборга, некоторые микробиологические среды.
Состав питательных сред для растительных клеток
Среда Мурасиге-Скуга
Компоненты ∙ Концентрация солей в 1 литре готового маточного раствора, мг ∙ Объем маточного р-ра, мл для приготовления 1 литра среды — маточный раствор макроэлементов NH4NO3 ∙ 33000 ∙ 50 KNO3 ∙ 38000 СаСl2*2Н2O ∙ 8800 MgSO4*7H2O ∙ 7400 КН2РO4 ∙ 3400 — маточный раствор микроэлементов KJ ∙ 166 ∙ 5 Н3ВО3 ∙ 1240 MnSO4*H2O ∙ 4460 ZnSO4*7H2O ∙ 1720 Na2MoO4*2H2O ∙ 50 CuSO4*5H2O ∙ 5 СоСl2*6Н2O ∙ 5 — маточный раствор хелатного железа FeSO4*7H2O ∙ 5560 ∙ 5 Nа2ЭДТА*2Н2O 7460 — витамины и органические вещества Мезоинозит ∙ 20000 ∙ 5 Никотиновая кислота ∙ 100 Пиридоксин-НСl ∙ 100 Тиамин-НСl ∙ 100 Глицин ∙ 400 — Добавлять в виде порошка в среду перед варкой: Сахароза ∙ 30 г/л Агар-агар ∙ 7 г/л pH готовой среды ∙ 5.6–5.8 * для получения стерильных проростков на 1 литр среды берут 1/2 часть маточных растворов, т. е. вместо 50 мл раствора макроэлементов берут 25 мл и т. д.
Среда Гамборга (В5)
Компоненты ∙ Концентрация солей в 1 литре готового маточного раствора, мг ∙ Объем маточного р-ра, мл для приготовления 1 литра среды — маточный раствор макроэлементов (NH4)2SO4 ∙ 2680 ∙ 50 KNO3 ∙ 50000 СаСl2*2Н2O ∙ 3000 MgSO4*7H2O ∙ 5000 NaH2PO4*H2O ∙ 3000 — маточный раствор микроэлементов KJ ∙ 75 ∙ 10 Н3ВО3 ∙ 3300 MnSO4*H2O ∙ 1000 ZnSO4*7H2O ∙ 200 Na2MoO4*2H2O ∙ 25 CuSO4*5H2O ∙ 25 СоСl2*6Н2O ∙ 2.5 — маточный раствор хелатного железа FeSO4*7H2O ∙ 5560 ∙ 5 Nа2ЭДТА*2Н2O ∙ 7460 — витамины и органические вещества Мезоинозит ∙ 10000 ∙ 10 Никотиновая кислота ∙ 100 Пиридоксин-НСl ∙ 100 Тиамин-НС1 ∙ 1000 — Добавлять в виде порошка в среду перед варкой: Сахароза ∙ 30 г/л Агар-агар ∙ 7 г/л pH готовой среды ∙ 5.5
Питательные среды для микроорганизмов и их приготовление[84] 1. Приготовление бобового агара В 1 литре воды кипятят 100 г белой фасоли или белых бобов до готовности, избегая растрескивания бобов и превращения крахмала в клейстер. Отвар фильтруют в горячем виде и добавляют 2 % агара. Стерилизуют в автоклаве как обычно. 2. Приготовление среды для хлореллы В качестве среды для культивирования зеленых водорослей можно использовать раствор Кнопа (г/литр раствора):
Са (NO3)2 — 0.25 MgSO4*7H2O — 0.06 КН2РO4 — 0.06 КСl — 0.08 Fе2Сl6 — одна капля 1 % раствора (можно заменить раствором хелатного железа из среды МС, 50 мл маточного раствора).На растворе Кнопа можно также выращивать и ряску.
3. Приготовление среды для цианобактерий Для культивирования цианобактерий рекомендуется среда Чу N 10 (г/литр раствора):
Са(NO3)2 — 0.04 MgSO4*7H2O — 0.025 KH2PO4 — 0.01 Na2CO3 — 0.02 Na2SiO3*9H20–0.025 FeCl3*6H2O — 0.0008 (также можно заменить хелатом железа)
4. Приготовление питательного агара для грибов (агар Чапека) Состав среды Чапека (г/литр раствора):
Сахароза — 30 г NaNO3 — 3 г КН2РO4 — 1 г MgSO4*7H2O — 0.5 г КСl — 0.5 г FeSO4*7H2O — 0.01 г Агар-агар — 15 гНа дно колбы насыпать агар (можно брать пол-литровые обычные бутылки, тогда на 1 литр среды необходимы 3 бутылки, в каждую их которых насыпать 1/3 агара, т. е. 5 грамм) и залить раствором солей. Колбу или бутылки поставить в автоклав на стерилизацию на 20 минут. После стерилизации, покачивая колбу, тщательно перемешать содержимое, иначе агар останется на дне, и застынет только нижний слой среды. В боксе среду разлить по стерильным чашкам Петри или пробиркам. Проверить на бактериальное заражение, выдержав 3–4 дня при комнатной температуре. В дальнейшем лучше хранить в холодильнике, так как там среда меньше испаряется.
5. Среда Прата для хранения коллекции культур водорослей Длительно выдерживать зеленые водоросли без пересева не рекомендуется, так как водоросли могут прекратить свой рост в результате самоотравления продуктами жизнедеятельности. Признаками такого угнетения могут служить пожелтение культуры, появление белесого оттенка на среде и ее помутнение. Сохранить коллекцию водорослей можно на твердых питательных средах, где рост культуры замедлен, поэтому частые пересевы не требуются (раз в месяц, а если хранить при слабом рассеянном свете при температуре 10–15 °C, то и раз в два месяца). Состав среды Прата (г/литр раствора):
KNO3 — 0.10 MgSO4*7H2O — 0.01 К2НРO4 — 0.01 FeCl3*6H2O — 0.001 агар-агар — 12 гСреду простерилизовать.
Методики культивирования клеток Начнем с того, что растительные и животные клетки предъявляют совершенно различные требования к условиям культивирования. Для животных клеток диапазон оптимального существования очень узок. Это касается и температурных условий, и состава питательных сред (необходимо строго выдерживать pH, молярность и т. д.), и газового состава. Пожалуй, наименее требовательны они к освещению. Для выращивания животных клеток всегда используют готовые фабричные питательные среды, а для длительного поддержания линии нужен термостат. Для некоторых видов клеток газовый состав воздуха можно обеспечить, поместив флаконы с культурой в эксикатор (емкость с притирающейся крышкой) в который также помещают горящую свечку. Когда эксикатор плотно закрыт, свеча затухает сама при концентрации углекислого газа в воздухе 5 %. Подробнее о условиях культивирования животных клеток и культивировании органов можно прочесть в учебнике по биотехнологии. Заводить и вести культуру растительных клеток гораздо проще. Растительные культуры поддерживают при температурах от 22 до 27 °C, в темноте или при освещении. Среды для них — несложные, и их легко приготовить самому. Культивировать можно на твердых питательных средах с добавлением агар-агара, крахмала (методика изложена в главе "питательные среды" или на жидких питательных средах, с использованием мостиков из ваты и фильтровальной бумаги. Суспензионные культуры растительных клеток поддерживать гораздо сложнее — необходима мешалка для постоянного перемешивания среды. Иначе эксплант погиб нет, в первую очередь, от отсутствия доступа кислорода. Условия освещения различны в зависимости от поставленных вами задач. Каллусную ткань обычно получают в темноте. Клональное микроразмножение проводят при освещении. В качестве источника света лучше пригодны лампы дневного света. Они не нагревают камеру и имеют более или менее подходящий для растений спектральный состав света.

ПОЛУЧЕНИЕ КАЛЛУСНОЙ ТКАНИ У РАЗНЫХ РАСТИТЕЛЬНЫХ ОБЪЕКТОВ
Каллус — определенным образом организованная масса ткани, состоящая из дедифференцированных клеток. Образование и рост каллусной ткани контролируется фитогормонами типа ауксинов и цитокининов. Под действием ауксинов происходит дедифференцировка, а под влиянием цитокининов — интенсивное деление, в результате которого образуется ткань. Спонтанное образование каллусной ткани в природе мы можем наблюдать на месте повреждения. Каллусные клетки затягивают рану, а впоследствии дифференцируются в утраченные ткани растения. Например, когда вы черенкуете розы или другие древесные растения, то на месте нижнего среза, погруженного в воду вырастает мозолеобразое образование — каллус, из которого затем формируются корни, происходит укоренение черенка. Очень часто каллус формируется на поврежденных корнеплодах моркови (особенно в месте среза листьев), на черешках кочанов капусты. Каллусную ткань также можно получить на искусственных питательных средах, включающих фитогормоны, используя фрагменты самых разных частей растения: стеблей, корней, тканей клубня, листьев, зародышей, семядолей и т. д. Такие фрагменты называются эксплантами.
Получение каллусной ткани из листьев табака Табак — классический объект для получения каллусной ткани и дальнейших ее исследований. Клетки табака легко дедифференцируются и переходят к делению, образуя быстро растущую каллусную ткань. Кроме того, в каллусной ткани табака легко вызвать регенерацию. Каллусообразование очень хорошо идет у основания листьев табака, особенно в зоне, примыкающей к срединной жилке. Для получения каллуса пригодны как стерильные, так и нестерильные растения. Нестерильные растения стерилизуют в растворе хлорамина, перекиси водорода или других стерилизующих агентов.
Материалы и оборудование Растения табака любого возраста, стерильные или нестерильные, этанол, 4 % раствор хлорамина, пробирки со стерильной водой (3 штуки), скальпель, пинцет, спиртовка, кафельная плитка, стаканчик, подставки для инструмента, кисточка, чашка Петри, спички, пробирки со средой МС с добавлением гормонов: 2 мг 2,4-Д и 0,2 мг кинетина на литр среды.
Ход работы Листья табака промыть в мыльном растворе, отмыть от мыла водопроводной водой, ополоснуть дистиллированной. Погрузить на 1 минуту в стаканчик с этанолом, вынуть, после чего поместить в чашку Петри и залить стерилизующим раствором на 10 минут. Раствор слить, промыть трижды стерильной дистиллированной водой из пробирок, в каждой из порций держать не менее 5 минут. Если берутся пробирочные растения, то предварительная стерилизация не требуется. Стерильные листья табака положить на предварительно обожженую кафельную плитку, стерильным скальпелем вырезать фрагменты у основания листьев, прилегающие к средней жилке, длиной 1–1.5 см, шириной 1 см. Перенести подготовленные участки листьев в пробирки или чашки Петри с питательной средой МС, содержащей гормоны, поместить в термостат при температуре 26 °C. Через 3 недели рассмотреть результат.
Получение и культивирование каллуса из стебля картофеля Материалы и оборудование Стерильные растения картофеля в пробирке или нестерильные проростки клубней картофеля, длиной 10–15 см., этанол, 4 % раствор хлорамина, пробирки со стерильной водой (3 штуки), скальпель, пинцет, спиртовка, кафельная плитка, стаканчик, подставки для инструмента, кисточка, стерильная чашка Петри, спички, чашки Петри или колбы со средой МС (с добавлением гормонов — 4 мг 2,4-Д и 0.2 мг кинетина на литр среды). В ответ на поранение паренхимные клетки, помещенные в питательную среду, содержащую фитогормоны, дедифференцируются, переходят к делению и образуют каллус. У картофеля каллус может быть получен из тканей стебля, листа, клубня, пыльника.
Ход работы Если используется стерильное растение из пробирки, то горлышко пробирки обжечь спиртом. Пинцетом вынуть стерильное растение из пробирки и поместить его в стерильную чашку Петри. Если берется нестерильное растение, то после промывки его оставляют в чашке Петри. Придерживая крышку в полуоткрытом состоянии, стерильным скальпелем вырезать скальпелем участки стебля длиной 5-10 мм, не захватывая междоузлия. Обжечь инструменты, перенести вырезанный участок на плитку, надсечь экспланты в нескольких местах для появления в дальнейшем раневого каллуса. Поместить их в колбу или чашку Петри со стерильной средой, чуть вдавливая их пинцетом, для усиления контакта со средой. В одну чашку Петри помешают 5-10 эксплантов. Закрыть колбу или чашку Петри. Чашки Петри заклеить парафилмом или пищевой пленкой. Поместить в термостат без освещения и выдержать 3 недели при температуре 22–25 °C. Через 3 недели рассмотреть образовавшийся каллус.
Получение и культивирование каллусной ткани из зародышей пшеницы Материалы и оборудование Зрелые зерновки пшеницы, пробирки со стерильной средой Гамборга с добавлением 5 мг на литр 2,4-Д (см. прописи питательных сред для растительных клеток), стерильный пинцет, скальпель, кисточка, стаканчик с этанолом, 4 % хлорамин, чашка Петри, кафельная плитка, колба со стерильной водой, спиртовка, спички. Каллусы, полученные из зародышей пшеницы, используют для селекции растений на солеустойчивость, засухоустойчивость, устойчивость к другимнеблагоприятным факторам.
Ход работы Семена пшеницы промыть водой с мылом, затем водопроводной водой, ополоснуть дистиллированной водой. Поместить в чашку Петри, залить этанолом на 5 минут, этанол слить в стаканчик, залить хлорамином на 15 минут, трижды промыть стерильной дистиллированной водой, выдерживая по 5 минут в каждой смене. Зерновку пинцетом поместить на плитку, вычленить зародыш, разрезать его поперек на 2 части. Верхнюю половинку зародыша, обращенную в зерновке к центру, поместить срезом на поверхность питательной среды в пробирке. Поместить в климокамеру при температуре 25 °C. Через три недели рассмотреть результаты.
Напоминание: не забудьте ознакомиться с методикой стерильной работы и методикой стерилизации растительного экспланта.

МИКРОКЛОНАЛЬНОЕ РАЗМНОЖЕНИЕ
Микроразмножение томатов или табака черенкованием побегов Материалы и оборудование Пробирочные растения томатов или табака, пробирки с питательной средой МС, стерильные скальпели, пинцеты, чашки Петри. Размножать некоторые растения можно путем черенкования в пробирочной культуре. Для этого растения разрезают на части. Черенки пересаживают в пробирки с питательной средой МС. Размножение черенкованием основано на подавлении апикального доминирования и активации пазушных меристем путем удаления верхушечного побега. При помещении побега на питательную среду из пазушных меристем появляется побег. Черенкование проводят с интервалом 14–21 день. Из 1 растения получают 5–8 черенков, за 2–3 месяца это составит 3–5 тысяч растений (при условии, что вы все делаете правильно и в условиях абсолютной стерильности).
Ход работы Пробирочное растение вынуть из пробирки, поместить в стерильную чашку Петри, разрезать на части (отрезок стебля с листом и пазушной почкой), часть стебля над листом должна быть при этом в 2–3 раза меньше, чем часть ниже лис та. Необходимо соблюдать строгую стерильность. Черенки перенести на свежую питательную среде в пробирки. Для предотвращения попадания микроорганизмов в пробирки обжечь горлышко и ватную пробку в пламени спиртовки. Пробирку поставить в световую камеру. Наблюдать за развитием побега.
Клональное микроразмножение роз Материалы и оборудование Однолетние побеги роз, скальпель, кисточка, пинцет, стаканчик с этанолом, пробирки со стерильной водой, 4 % раствор хлорамина, чашка Петри, кафельная плитка, спиртовка, спички, пробирки с модифицированной питательной средой МС, ее компоненты:
— макроэлементы по прописи МС — микроэлементы по прописи МС — хелат железа витамины по прописи МС — источник углерода: глюкоза — 40 г/л — агар-агар 7 г/л — гормоны: гиббереллин — 0.1 мг/л, 6-ЕАП — 2 мг/л — pH готовой среды 5.6–5.8).
Ход работы В качестве экспланта используют боковые почки из средней части 1-летнего побега. Побеги промывают водопроводной водой с мылом, ополаскивают дистиллированной водой, вырезают почку. Стерилизуют сначала 70 % этанолом в течение 1 минуты, а затем в растворе хлорамина 10 минут. Трижды промывают стерильной дистиллированной водой (в каждой порции по 10 минут). Срезы обновляют, снимают почечные чешуи и переносят почки на агаризованную питательную среду в пробирки. В некоторых случаях экспланты переносят на свежую питательную среду, так как в среду выделяются полифенолы. Культивируют при температуре 18–25 °C, освещенности 500-1000 люкс, 16-часовом фотопериоде. Пассируется через 3–4 не дели. Для укоренения побеги переносят на среду Гамборга (В5) с добавлением 1 мг/л ИУК и увеличивают интенсивность освещения до 2000 люкс.
Перенос посадочного материала в почву Материалы и оборудование Пробирки с растениями, горшки с почвой; почву желательно прогреть или проавтоклавировать до посадки растений.
Ход работы Сделать пробиркой углубление в увлажненной почве в горшке. Пробирочное растение вынуть из пробирки, поместить в горшки с почвой, не отмывая агаровой среды. Уплотнить почву вокруг растения. Первые 3–5 дней желательно прикрывать растения стеклянным колпаком. Через 10 дней растения пересаживают на постоянное место в теплице.

ПОДДЕРЖАНИЕ И СОХРАНЕНИЕ КЛЕТОЧНЫХ КУЛЬТУР
Поддержание и сохранение культуры картофеля Материалы и оборудование Пробирки с растениями картофеля, стерильный пинцет, скальпель, кисточка, стаканчик с этанолом, чашка Петри, кафельная плитка, спиртовка, спички, пробирки с питательной средой МС. Для замедления роста растений из черенков ТУР (ретардант, применяемый в сельском хозяйстве против полегания стеблей растений — хлорхолинхлорид, ССС) в концентрации 0.4–0.5 мл/л добавляют в среду МС, при этом промежуток между двумя последующими черенкованиями увеличивается с 1 до 4 месяцев, а высота со сближенными междоузлиями не превышает 3–4 см. Растения приобретают темно-зеленую окраску. Одним из способов сохранения и поддержания коллекции in vitro является клубнеобразование в пробирках. На процесс образования клубней in vitro оказывает влияние фотопериод, температура, ростовые вещества, содержание сахарозы в питательной среде.
Ход работы Черенки переносят на модицифированную питательную среду МС для образования клубней картофеля, ее компоненты:
Макроэлементы по прописи МС Микроэлементы по прописи МС Источники железа Витамины по прописи МС + аденин — 0.25 мг/л Источник углерода: сахароза — 70 г/л Агар-агар — 7 г/л Гормоны: ИУК — 1 мг/л pH готовой среды — 5.6–5.8.Культивирование черенков проводится под люминесцентными лампами при 10 часовом фотопериоде, освещенности 3.5–4 тыс. люкс, влажности 70 %. После трехнедельного выдерживания на 10-часовом фотопериоде растения переносят на 2 суток в темноту, а затем на 16 часовой период и оставляют в этих условиях до завершения клубнеобразования. После отмирания растений и достижения микроклубнями размера 0.7–1.4 см их извлекают из пробирок (в стерильных условиях) и хранят в пробирках, закрытых ватными пробками, в холодильнике, при температуре +2 — +4 °C.
КУЛЬТИВИРОВАНИЕ ЖИВОТНЫХ КЛЕТОК
Культивирование фибробластов человека Культуральная посуда и оборудование Пипетки мерные от 1 до 10 мл, пипетки Пастера, пенициллиновые флаконы, пробирки Лейтона, мерные флаконы от среды разных объемов, пробки резиновые № 10, 12.5, 14.5, 24; стекла покровные для микроскопии, предварительно нарезанные для помещения их в пенициллиновые флаконы, пинцеты глазные анатомические и хирургические, зажим Кохера, бритва безопасная, препаровальная игла из нержавеющей стали, резиновая груша для работы с пипетками (с надетой резиновой трубкой), инвертированный микроскоп, ламинар-окс, термостат.
Питательная среда Фибробласты успешно культивируют на разных средах, но рекомендуется использовать следующий состав питательной среды — среда Игла — 90–85 %, сыворотка крупного рогатого скота — 5-10 %, пуповинная сыворотка человека (фетальная сыворотка) — 5 %, глютамин — 150 мг на 500 мл готовой среды. Готовую среду и её компоненты рекомендуется хранить при 4 °C. При культивировании эксплантов рекомендуется использовать антибиотики (40–60 мкг канамицина на 1 литр среды).
Ход работы 1. Взятие биопсии. Фибробласты можно получать из многих тканей и органов. Проще всего — фибробласты дермального слоя кожи. Наиболее удобно брать биопсию со внутренней стороны предплечья левой руки (нижняя треть). Кожу дезинфицируют 70 % этиловым спиртом. Не рекомендуется употреблять другие дезинфицирующие растворы, так как в случае проникновения в биопат они могут оказать токсическое действие. После дезинфекции кожу захватывают глазным хирургическим пинцетом, оттягивают и отрезают кусочек кожи непосредственно под браншами пинцета. Не следует стремиться взять глубокие слои кожи. При правильно взятой биопсии, захватывающей эпидермис и сосочковый слой кожи, кровотечение минимально. Отсечение лучше проводить лезвием безопасной бритвы, простерилизованной (как и пинцет) путем погружения в этиловый спирт с последующим поджиганием. Отсеченный кусочек погружается в заранее приготовленный пенициллиновый флакон со средой. На рану накладывают бактерицидный пластырь. Фрагмент кожи может храниться в питательной среде при 4 °C в течение 3–4 суток без утраты фибробластами способности к росту. 2. Получение экспланта. В стерильную чашку Петри с каплей 0.25 % раствора трипсина или питательной среды помещают фрагмент кожи. Придерживая пинцетом, нарезают на мелкие кусочки величиной с булавочную головку и меньше стерильным лезвием безопасной бритвы, зажатой в зажим Кохера или Пеана. При помощи изогнутой препаровальной иглы кусочки извлекают из капли и помещают на покровное стекло, находящееся в пенициллиновом флаконе или пробирке Лейтона. Уложенные кусочки накрывают другим покровным стеклом, слегка прижимая препаровальной иглой. Наливают в культуральный сосуд питательную среду так, чтобы она покрыла верхнее стекло. Покровные стекла с зажатыми между ними кусочками кожи при помощи пинцета кладут на боковую стенку пенициллинового флакона. Флаконы плотно закрывают пробкой и устанавливают на подставке под углом примерно в 30°. Культуры инкубируют в термостате при 37 °C. В течение недели следят за цветом среды. Изменение цвета индикатора к желто-оранжевому свидетельствует о хорошем состоянии экспланта. Через неделю ежедневно начинают просматривать культуру под микроскопом. Можно использовать как инвертированный, так и обычный микроскоп. В последнем случае следует осторожно повернуть флакон, при этом стекла с культурой прилипают к боковой стенке флакона и могут быть исследованы при малом увеличении. Вокруг кусочков вначале появляются единичные фибробласты, которые заметны в виде длинных веретеновидных клеток, либо появляется слой эпителия, окружающий фрагмент в виде "кружевного" ореола. При появлении зоны выроста клеток следует сменить среду. Когда клетки займут все пространство между кусочками (это видно невооруженным глазом или при помощи лупы), их пересевают.
Пересев фибробластов человека Материалы и оборудование Флаконы с питательной средой, трипсин, покровные стекла, пипетки, препаровальные иглы. Пересевы производят для получения количества клеток, достаточного для цитогенетического или биохимического исследования, а также для криоконсервации. Клеточный штамм считают установившимся, если после пассирования в течение первых 5 пассажей прирост клеток спустя 3–4 суток после пересева превосходит количество посеянных минимум вдвое, что позволяет пересевать такие клетки в отношении 1:2 (из одного сосуда в два).
Ход работы Удаляют питательную среду из культурального сосуда и наливают 2–3 мл подогретого 0.25 % раствора трипсина (или смесь версен-трипсин). Через 2 минуты удаляют трипсин, оставив его только между стеклами. Инкубируют в термостате 15–20 минут при 37 °C. Ход трипсинизации можно контролировать, просматривая культуры под микроскопом. По окончании трипсинизации при помощи препаровальной иглы отделяют одно стекло от другого и при помощи тонко оттянутой пипетки смывают клетки небольшим количеством среды (1 мл). При этом несколько раз пропускают через пипетку суспензию клеток и кусочки. Суспензию клеток переносят в пенициллиновый флакон и добавляют среду до 2–3 мл. Оставшиеся в прежнем сосуде кусочки снова укладывают между двух покровных стекол и заливают средой. От этих вторично культивируемых эксплантов может быть получен второй "урожай" клеток. Флакон с пересаженными растущими фибробластами просматривают ежесуточно под микроскопом. Когда образуется слившийся слой клеток, производят второй пересев (обычно через 4–7 суток).
Контроль штаммов Ежедневный просмотр культуральных сосудов для оценки цвета среды и её прозрачности. Резкое изменение цвета и появление мути может свидетельствовать о бактериальном или грибковом заражении. Если цвет индикатора среды не изменяется после пересева или сдвигается в сторону малинового, это может быть связано с отсутствием размножения клеток, с недостаточным количеством для данного объема или гибелью. При помощи инвертированного микроскопа оценивается морфология клеток и слоя в целом. В нормальных условиях фибробласты — веретенообразные вытянутые клетки с прозрачной цитоплазмой. Отсутствие рисунка на субстрате (фибробласты обладают ориентированным ростом), появление зернистости в цитоплазме, пустые места в слое клеток, распластанность клеток свидетельствует об их дегенерации в результате токсических или иных вредных воздействий. Небольшое количество среды после каждого пересева отливают для проверки на стерильность, которую производят путем посева на агар.
ПРАКТИКА
Разведение пивных дрожжей в домашних условиях
Е. Свидерский
Довольно часто перед домашним пивоваром встает вопрос, где взять качественные пивные дрожжи. Сейчас достаточно много фирм предлагают сухие дрожжи низового брожения, которые великолепно подходят для домашнего пивоварения. Однако их ассортимент в России очень ограничен. Как правило, дрожжи «интересных» сортов привозятся из заграничных командировок. При этом их количества весьма ограничены и без принятия соответствующих мер все расходуется за пару варок, поэтому становится актуальна проблема размножения дрожжей дома. Самый простой способ практически не требует усилий. Достаточно после брожения собрать дрожжевой осадок в банку и хранить в холодильнике под слоем чистой воды. Однако при этом дрожжи очень быстро деградируют, в них попадают посторонние бактерии и через пару варок их смело можно выбрасывать, ничего приличного с их помощью уже сбродить не удастся. Если хочется сохранить дрожжевую культуру надолго и с минимальными потерями качества, придется основательно потрудиться. Во-первых, очень возрастают требования к стерильности. Если маленькое попадание посторонних бактерий в ферментатор обычно не фатально, то в данном случае оно способно сильно испортить ситуацию. Для начала готовим питательную среду. Кипятим примерно 6–8 % раствор Малтакса[85] минут 15–20 до его стерилизации. Остужаем раствор, сливаем чистую жидкость от белковых хлопьев на дне. Кипятим еще раз, прямо в банке на водяной бане (водяная баня — это обычная кастрюля, в которую налито сантиметров 5-10 воды), при этом простерилизуется как банка, так и жидкость. Охлаждаем банку в закрытом виде. Дальше зажигаем газ, ставим банку рядом с огнем (сантиметров 5-10), открываем крышку и стерильной ложкой (можно ее прокалить на том же газу) взбалтываем Малтакс, чтобы немного насытить его кислородом. Огонь нужен для того, чтобы создать восходящий поток воздуха. При этом есть гарантия, что в процессе перелива никакая пылинка или злобная бактерия не упадут в нашу банку сверху. Засыпаем дрожжи, закрываем, но не герметично и ставим в темное место. Спустя пару дней, не дожидаясь полного окончания брожения, необходимо поставить банку в холодильник, чтобы остановить брожение. Ждем еще с день-два, дрожжи оседают на дно. Аккуратно сливаем жидкость (совсем хорошо перед этим протереть спиртом горловину банки и делать переливы у огня). После этого промываем дрожжи. Для этого сливаем с дрожжевого осадка жидкость, наливаем чистую стерильную охлажденную воду, закрываем, взбалтываем, и оставляем на день в холодильнике. Повторяем процедуру 2–3 раза. Теперь дрожжевой осадок нужно поставить на хранение. Вариант попроще: на лить немножко воды (разумеется, прокипяченной, стерильной и охлажденной), взболтать и разлить по стерильным баночкам, после чего хранить в холодильнике примерно при плюс 5 градусах. Более продвинутый вариант — добавить в воду процентов 10–15 глицерина и хранить в нулевой камере при температуре около нуля. В первом случае срок хранения примерно месяца 2, во втором — около 4 месяцев, потом требуется процедуру размножения дрожжей повторить. Перед употреблением дрожжи нужно разбраживать, т. к. их концентрация намного меньше, чем в случае применения сухих дрожжей. Разбраживать дрожжи желательно за день до варки пива. Для этого нужно прокипятить в баночке раствор Малтакса, как мы делали для размножения дрожжей. Остудить и влить туда дрожжи из хранения. Перед этим их очень желательно подержать при комнатной температуре хотя бы час, чтобы перепад температур дрожжей и питательной среды был не более 10 градусов. Потом закрываем банку (не герметично) и оставляем на ночь. На следующий день проверяем, появился ли в банке характерный слой пены. Если нет, варку лучше отложить, т. к. все равно полученное сусло нечем будет сбродить. Если же брожение началось, проводим варку сусла и после его охлаждения вливаем в него содержимое банки с дрожжами (перед переливом взболтать). Если хочется еще продлить сроки использования дрожжей, придется потрудиться дополнительно и провести очистку дрожжевой культуры от посторонних примесей, которые туда все равно попадут, несмотря на все старания. Понадобится чашка Петри (штуки 2–3). Готовим питательную среду. Для этого в вышеупомянутый раствор Малтакса засыпаем желатин из расчета примерно 10–15 % по массе. Тонким слоем наливаем на чашку (разумеется, все стерильное и около огня). Накрываем и ждем затвердевания. Разбавляем раствор дрожжей раз этак в 1000. Капаем каплю на застывший желатин, равномерно распределяем прокаленной ложкой, накрываем и оставляем. Потом разбавляем еще на порядок и еще на порядок, окучивая оставшиеся чашки. Через пару дней на желатине прорастут колонии из попавших туда клеток. Выбираем ту чашку, где эти колонии пространственно разделены (при большой концентрации исходных дрожжей там сплошная каша будет). Дрожжи там смотрятся как маленькие точки, а всякая бяка как маленькие мутные пятнышки. Несколько точек соскребаем иголкой (опять все стерильное) и кидаем в небольшой бутылёк с раствором Малтакса, там они слегка размножаются, после чего переносим их в большую банку и продолжаем процесс[86]. При выполнении этих мер вполне можно в домашних условиях получать чистую дрожжевую культуру. Разумеется, это не самый простой путь, но разве кто-то обещал легкую жизнь?
Жарим зерно дома
Г. Сандерс и С. Лэсей
ЗАЧЕМ[87]?
В дебрях Северного Куинсленда[88] просто полным-полно тропических фруктов, но ни одной солодовни, сколько ни кричи «куи!»[89]. На заре пивоварения считайте, что Вам повезло, если удавалось достать хотя бы солод Schooner[90]. И когда это случалось, он был столь же свежим, как и проеденный червями кенгуру. Вот тогда то я и начал экспериментировать. Быстрая обжарка этого зерна в духовке приводила самое несвежее зерно хотя бы в более-менее приличное состояние. С годами обстоятельства менялись к лучшему, стал появляться кристаллический солод, а также свежий солод Schooner. Все бы ничего, да у нашего кристаллического были (и до сих пор еще остаются) неизвестно какие характеристики и цветность. Сегодня предлагается довольно хороший ассортимент, хотя практически невозможно заполучить специальное зерно, такое как Special В, копченый или коричневый солод, а поставки других видов солода в лучшем случае нестабильны. Поэтому для меня, домашняя обжарка основывалась на необходимости. Но также она обладает одним преимуществом, которое многие пивовары-цеховики на самом деле упускают. Это все равно, что покупать свежеиспеченный или суточной хлеб, разница ЧУВСТВУЕТСЯ. Так что, испытываете ли Вы трудности с поставками жареных солодов или просто хотите попробовать добавить особую нотку в ваше пиво, Вы можете прийти к выводу, что Вам действительно хочется попытаться пожарить солод дома.
КАКОЕ ОБОРУДОВАНИЕ ПОНАДОБИТСЯ?
Всего лишь духовка, термометр для духовки и противни. Вот теперь можно было бы купить ртутный стеклянный термометр, который меряет до 300 °C, но есть вариант намного дешевле. Зайдите в какой-нибудь магазин для кулинаров и поищите плоский металлический термометр для духовки. Они, может быть, не очень точные, но достаточно хороши для рассматриваемой работы, и довольно дешевы. С ними можно работать в горячем виде, их можно ронять, швыряться ими в гневе, они идеальны. Обычные термометры также могут подойти, но они должны быть в состоянии выдерживать высокие температуры. У меня есть ртутный термометр, который выдерживает до 350 °C. Главный их недостаток в том, что на них непросто прочитать показания и когда достаешь их из духовки, они очень горячие. Это верный способ их разбить.

"Зачем термометр?" спросите Вы. "У меня чудесная духовка, я могу настроить на ней нужную температуру, местную радиостанцию и даже поймать полицейскую частоту". Ну, эти шкалы редко бывают точными, а датчик на самом деле находится на стенке духовки, а не на самом зерне. Вам нужно мерить температуру на поверхности зерна. Вот где также хорошо подойдут эти металлические термометры. Вы можете положить их сверху на зерно, и следить через окно вашей духовки за температурой зерна. Разберитесь с Вашей духовкой, Вы должны знать, что в ней происходит на самом деле. Установите ваш поднос примерно посередине и выставите регулятор на 100 °C. Пронаблюдайте, когда температура стабилизируется, проделайте это снова скажем при 150 °C и так далее. Обратите внимание, какие изменения происходят с температурой. А также посмотрите, как различная высота, на которую устанавливается противень, влияет на изменение температуры. Так, что же Вам нужно? Две вещи. 1. Вам нужно узнать, что на самом деле нужно выставить на ручке-регуляторе, чтобы получить требуемую температуру. 2. Вам нужно добиться минимальных колебаний температуры. Многие духовки имеют так называемые «горячие пятна», их нужно избегать. Обычно следует установить поддон(ы) посередине духовки. Вам также понадобится противень для зерна. Идеально подойдет что-нибудь такое, во что можно было бы насыпать слой примерно в один дюйм толщиной. Если он не совсем чист, выложите его алюминиевой фольгой.
ПРОЦЕСС
То, что происходит на солодовне, на самом деле довольно простой процесс. В основном, как только ячмень прорастает, происходит одна из двух следующих вещей. 1. Ячмень сушат и затем жарят. Чем дольше Вы его жарите, и чем выше температура, тем зерно получается темнее. Привожу базовые солода. В порядке повышения цветности это pilsener, пэйл, венский, мюнхенский, янтарный, бискит, коричневый, шоколадный, темный (или могут быть похожие названия); либо 2. Ячмень оставляют влажным и жарят. Опять же, чем дольше и горячее жарите, тем темнее результат. Таким образом Вы получаете карамельный солод. Опять же в порядке повышения цветности: carapils, caravienna, caraMunich, различные кристаллические солода, специальный В. Существует еще пара других вариантов. Прокоптите зерно, и Вы получите rauch (раух) солод, опять же обжарка просто сырого ячменя добавит пиву дополнительный вкус. Жарьте его до тех пор, пока он не станет черным, и у Вас будет жареный ячмень для стаута. Зачем же останавливаться на ячмене, чем не подходит пшеница или рожь? Варианты бесконечны.

На этой фотографии показано, что Вы можете жарить зерно даже в микроволновой печи, если в ней установлен специальный датчик.
Обратите внимание, преимущество здесь за простым металлическим термометром для духовки. Он легко читается, и его не нужно убирать. Он показывает 130 °C, в то время как я поджариваю кристаллический солод.
Но давайте рассмотрим основные солода. Все они в основном происходят от одного из базовых солодов, pilsner и carapils[91]. То, что солодовник делает с ними впоследствии, определяет конечный продукт. То есть, Вы можете взять carapils и сделать Belgian special В (бельгийский специальный В), если Вы знаете, как этого добиться. Но Вы можете также использовать кара-мюнхенский и сделать специальный В. Фактически, пока Вы идете "по линейке вверх", Вы можете сделать любой солод из более светлого. Так какие же солода мне нужно иметь дома, из тех которые понадобятся. Можно обзавестись pilsener солодом, как импортным, так и отечественным. Он не дешевый, даже если берете оптом, и он особенно хорош для светлых континентальных лагеров. Поэтому я его оставляю, чтобы использовать самому. Дальше идет пэйл солод (обычно Schooner). Теперь это солод для пивзаводов-гигантов, который производится в промышленных масштабах. Он не столь же хорош, как и pilsener солод, мне кажется, он слегка декстриновый, но он хорошо модифицированный, дешевый и легкодоступный. Это мой любимый солод для получения обычных жареных солодов. У меня с ним нет проблем при получении из него солодов вплоть до шоколадного. Для карамельных солодов я использую темный carapils (или очень светлый каравенский, выбор за Вами). Я предпочитаю темноватый carapils, чтобы гарантировать наличие в нем карамельного вкуса. Из него я без проблем получаю все виды карамельного солода, даже бельгийский Специальный В. Так каковы же методы, которыми мы можем воспользоваться, чтобы пожарить свое зерно. Один метод, который я не одобряю, носит название "быстрый и резкий". При этом методе Вы производите сильный нагрев за короткий промежуток времени. Метод прекрасно работает, зерно жарится очень быстро. Но я его терпеть не могу, потому что на самом деле он не соответствует тому, что происходит на солодовне, он не дает однородного результата, используя его слишком легко все испортить, и Вам необходимо почти постоянно перемешивать солод во избежание появления горячих пятен и горения. НО, этот метод действительно позволяет Вам в критической ситуации быстро получить немного жареного зерна. Его стоит попробовать, когда больше уже ничего не поделаешь, но я не собираюсь распространяться на эту тему.
ШАГ ЗА ШАГОМ
На самом деле в этом нет ничего сложного. Все, что Вам нужно сделать, это выставить нужную температуру и дать духовке прогреться. Тем временем на подносе разложите зерно толщиной примерно в дюйм[92]. Не кладите слишком много. Лучше сделать несколько партий, чем выложить зерно более толстым слоем. Термометр положите на зерно, и поставьте его в духовку. На прогрев зерна до нужной температуры понадобится около двадцати минут (в зависимости от духовки). Не ВКЛЮЧАЙТЕ таймер, пока все зерно не прогреется до нужной температуры. Быстро перемешайте и включите таймер.

Фотография закрытой дверцы духовки. Хотя на фотографии не очень хорошо видно, преимущества такого типа термометра очевидны. Нет необходимости открывать дверь духовки. Вы можете легко прочитать показания температуры через окошко.
Все, что Вам остается теперь делать, это перемешивать каждые полчаса, пока не будет готово. Ну, пока что все и ёжику понятно. Если Вы используете больше одного поддона, неплохо, если они при этом бы еще и вращались. Так, что за зерно, сколько времени и какая температура. Потребуется немного поэкспериментировать в зависимости от вашей духовки, но чтобы все то, что Вы делаете, получилось, нужно достать образцы промышленного солода для сравнения их между собой (если получится). На самом деле Вы удивитесь, насколько близкий результат у Вас может получиться. Но что делать, если нет никаких образцов. Тогда Вы примерно соблюдаете температурные режимы, принятые на солодовнях. Если Вы не особенно любознательны и хотите копировать то, что делает кто-то еще, тогда я скажу Вам температуры, которые использую я.
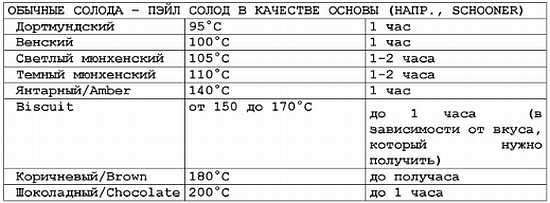
При жарке сырого ячменя мне нравится подержать его 45 минут при 190 °C, затем 30 минут при 230 °C, или пока 10 % зерен не станут очень темными, а 10 % светло-коричневыми. Вы заметите, что чем сильнее нагрев, тем фактор времени становится более гибким. Это происходит оттого, что при более низкой температуре можно не заметить, что имеют место значительные изменения, в то время как они действительно происходят. При более высокой температуре Вы заметите, что изменения происходят довольно быстро, и Вы даже можете определить цвет вашего солода. Поэтому, если в Вашем рецепте уже заложено много темных добавок, Вы делаете свой жареный солод более светлым. Ну и конечно, если рецепт предполагает светлый цвет, Вы также можете сделать его более темным. Все это замечательно подходит для создания рецепта. Существует один фокус, позволяющий добиться наличия в венских и мюнхенских солодах вкусных меланоидинов. Я обнаружил что, если зерно предварительно немного увлажнить приблизительно на час, часть сахара выступает на поверхности. В течение сушки он снова высыхает, но смачивание способствует появлению дополнительного цвета, но что еще более важно, лучшей выработке меланоидинов. Для получения раух (rauch) солода я также немного сильнее увлажняю зерно и копчу его в горячем дыму. Поглощенный солодом аромат дыма получается намного сложнее и насыщенней (поэтому я могу использовать гораздо меньше). Ну а теперь, что же нам делать с карамельными солодами? Ну, если Вы их немного увлажните, то сохраните ту самую карамельную образующую, таким образом Вы можете получить собственный кара-венский, кара-мюнхенский и бельгийский специальный В солода. Пожарите их без увлажнения, и Вы получите очень хорошую замену кристаллическому солоду.
НАЧИНАЕМ С БОЛЕЕ ТЕМНОГО КАРАПИЛСА (ВСЕ ТЕМПЕРАТУРНЫЕ РЕЖИМЫ ОКОЛО 12 °C) Cara-Vienna… до 1 часа Cara-Munich… 1–2 часа Светлый crystal… 1–2 часа Темный Crystal… 2–3 часа Special В… 2–3 часа
Теперь последний совет. Я всегда жарю зерно накануне его использования. Это как раз тот самый синдром свежего хлеба. Чем свежее, тем лучше. На самом деле я делаю исключение для своего "беспощадного" раух солода. Я даю ему проветриться несколько недель, с тем, чтобы из него ушли некоторые наиболее летучие вещества. Солод получается мягче. Существует также противоположное мнение, из которого следует, что любому солоду домашней жарки необходимо дать "полежать" неделю-другую для "созревания". Само собой разумеется, я не подписался на эту теорию, но, честно говоря, Вы должны понимать, что некоторые люди верят этому, и Вы вольны сами для себя все решить. Ну, вот, пожалуй, и все. Ваш выход!
ТЕХНОЛОГИИ
Эли бочкового созревания
Д. Буш
Англия имеет много отличительных особенностей, которые внесли свой вклад в мир пивоварения, но ни одна из них не отличается той же уникальностью и важностью как традиция производства элей бочкового созревания. Бочковой, или как его часто называют, риэл эль является специальным пивом, которое подается особым образом, при помощи ручного насоса из подвала. Несмотря на то, что Вам, возможно, рассказывали в Соединенных Штатах, бочковой эль не теплый и не без газа. Его разливают при температуре подвала, от 54 до 59 градусов F (от 12 до 15 градусов С), и естественно, хоть и слегка, но он газированный. В результате, по палатальным ощущениям (mouthfeel) он отличается от "газированного" пива из кега. Температура и карбонизация оказывают большое влияние на вкусовое восприятие пива, и комбинация подвальной температуры, низкого содержания СО2 и зачастую резкого хмельного аромата и вкуса — все это смешивается во рту — и выявляет совершенно особый и приятный эль. В то время как весь спектр бочковых элей обобщить довольно нелегко, карбонизация, температура, степень охмеления и продукты брожения обычно являются определяющими факторами во вкусовом восприятии всех бочковых элей. Многие бочковые эли обладают многочисленными фруктовыми нотками, которые рождаются в ферментере и постепенно ослабевают и перемешиваются в процессе созревания и выдержки. Важным моментом является то, что они должны присутствовать и то, что они проявляют себя в различной степени сложности на протяжении всего нахождения в бочке. Одним из замечательных аспектов бочкового эля является то, что это пиво живет, дышит, и оно изменится за неделю или около того в промежутке между шпунтованием или закупориванием и последним движением ручного насоса.
ПРОИЗВОДСТВО БОЧКОВЫХ ЭЛЕЙ
Бочковые эли, изготавливаемые в Англии, являются пивом верхового брожения, часто производимым в открытых ферментерах. Открытые ферментеры — это то, что и кажется — емкость без верхней части. Ферментеры часто представляют собой большой цилиндр с навесной крышкой. Многие из них терморегулируются (охлаждаются) при помощи системы труб, которые погружены в бродящее сусло. Через трубопровод прокачивается охлажденная вода или охлажденный гликоль, что позволяет пивовару контролировать температуру в процессе брожения. В то время как закрытые танки используются на некоторых более крупных пивоваренных заводах, открытые ферментеры являются традиционной практикой, и некоторые известные пивзаводы полагаются на старые системы открытого брожения Burton Union и Yorkshire Squares. Оба типа этих открытых бродильных аппаратов разработаны таким образом, что бродильному оттеку или кройзену (kraeusen, шапка бродящей пены) дают вытечь из ферментера в сборный резервуар, затем его удаляют или возвращают в главный ферментер. Эта техника обычно способствует тому, что в ферментер поступает дополнительный кислород, что в свою очередь приводит к слегка повышенному уровню диацетила в пиве. Обычно это не является отрицательным моментов для данного пива. Использование открытого брожения может показаться странным пивоварам, которые идут на все, лишь бы не допустить воздушного заражения, но это не является причиной для беспокойства в английском пивоварении. Как и все пивовары, английские элевые пивовары очень внимательны в вопросах санобработки всего, что контактирует с полученным суслом, особенно, когда его температура становится ниже 170 градусов F (76.5 градусов С). Чистый и продезинфицированный ферментер в сочетании с чистыми, здоровыми дрожжами, заданными в количестве от 6 до 12 миллионов клеток на миллилитр, обеспечит быстрое начало брожения, и соответственно выработку большого количества СО2, который покроет бродящее сусло, и таким образом защитит пиво от витающей в воздухе инфекции. Как только брожение активизируется, pH пива быстро понизится с начального уровня, который равен 5.0, где-то до середины диапазона, составляющего 4, а при использовании некоторых рас, вплоть до 4.1. Это подкисление сусла наряду с большим образованием СО2 приводит к возникновению среды, весьма неблагоприятной для большинства бактерий, находящихся в воздухе. Главное, и это относится ко всему пивоварению, состоит в том, чтобы внести достаточное количество чистой, здоровой суспензии дрожжевой культуры. Используя открытые бродильные аппараты, пивовар должен снимать с пива дрожжевую шапку между вторым и третьим днем нормального брожения. То, что поднимается на поверхность после первого дня, удаляется с целью уменьшения количества твердых частиц, которые могут привести к возникновению проблем с терпкостью. Использование открытых ферментеров обеспечивает легкий способ наблюдения и выполнения требований по снятию пены в элях верхового брожения. При использовании обычных рас верхового брожения, здоровые белые дрожжи собирают на третий или четвертый день брожения и сохраняют их для повторного использования. Дрожжи, собранные в процессе здорового брожения, могут быть повторно использованы на протяжении сотен поколений, при условии, что на пивзаводе соблюдается чистота, а пивовар внимательно следит за характеристиками брожения. Любое ухудшение работы дрожжей должно быть исправлено заменой штамма свежим. Брожение обычно завершается за пять-семь дней при температуре от 60 до 70 градусов F (от 15.5 до 21 градуса С). После этого пиво переливают в танки для выдержки, где оно может находиться, проходя непродолжительный период созревания. В качестве альтернативы, негазированное пиво можно перелить непосредственно в бочку. Важным моментом является то, что перелив необходимо производить так, чтобы в негазированном пиве оставалось бы около 1° Плато (1.004) остаточного экстракта, количество дрожжевых клеток составляло бы от 0.25 до 2 миллионов на миллилитр негазированного пива. Остаточный экстракт также может быть обеспечен в виде праймер-сахаров. Это достигается путем приготовления раствора пивоваренного сахара (глюкозы) плотностью 1.150 (34 ° Плато) и добавления его в бочку в количестве от 0.35 до 1.75 литра на гектолитр. Также может быть использован оттек сусла или кройзен пиво, но в последнем случае, чрезмерное количество дрожжевых клеток может помешать процессу осветления в бочке. Когда бочки будут заполнены, в емкость добавляют осветлитель, обычно в форме рыбьего клея (isinglass) в количествах от одного до пяти литров на британский баррель. Isinglass состоит из коллагеновых молекул, которые обладают общим положительным зарядом. Поскольку дрожжи имеют общий отрицательный заряд, в результате образуется электростатическое притяжение, которое приводит к слипанию дрожжей и частиц рыбьего клея, а затем к выпадению их в осадок. В процессе осветления понадобится около суток для достижения "отменного блеска" пива. Пиво с остаточным уровнем дрожжей от 2 и более миллионов клеток на миллилитр осветлить будет сложнее. Многие пивовары в момент наполнения бочки также добавляют хмель в виде шишек в количестве от половины до трех унций на баррель. С появлением современных способов тарирования, таких как бочонки polypin, некоторые пивовары стали использовать экстракты хмелевого масла для того, чтобы имитировать некоторые характеристики, присущие элям с бочковым охмелением. Теперь бочковой эль можно отправлять в подвал бара.
СОЗРЕВАНИЕ БОЧКОВЫХ ЭЛЕЙ
После того, как бочка отправлена в местный паб, работа по окончательному созреванию пива уже не является прямой ответственностью пивовара. Эта задача возлагается на владельца бара или погребного мастера (cellarmaster). В старые времена в обязанность погребного мастера входило добавление осветлителей в бочки по прибытии их с пивзавода, но сегодня это не принято. После доставки в паб бочку помещают на стеллаж и дают ей отстояться два-три дня. В это время в бочке происходит вторичное брожение, или бочковое созревание. За день перед началом использования, бочку нужно подготовить для розлива. Это делается путем вбивания твердой пробки (непористой деревянной втулки) в шпунт (круглое пробочное устройство на верхней части бочки или эквивалент затычки на старых американских кегах). Втулка — это по существу примитивный СО2 клапан, непористая втулка используется для закрывания бочки на ночь на хранение, в то время как пористая втулка используется при розливе, впуская воздух и позволяя пиву вытекать при помощи пивного насоса. Когда втулку первый раз вколачивают в бочку, погребной мастер дает углекислому газу выйти из бочки, предотвращая таким образом повышение уровня СО2, который не приветствуется любителями риэл эля. Заключительный шаг в подготовке бочки к использованию состоит в забивании крана в keystone (собственно отверстие, через которое "выходит" пиво). Как минимум сутки потребуются для отстаивания, чтобы получить гарантию, что процесс подготовки не взболтал слишком много дрожжей. На следующий день, погребной мастер попробует пиво, чтобы определять, что оно готово. Это чрезвычайно важная часть процесса и основная причина, почему многие бочковые эли не подаются при наступлении своего вкусового пика. Некоторому пиву требуется немного больше времени, чем другим, для достижения этого максимума.
РОЗЛИВ БОЧКОВЫХ ЭЛЕЙ
Когда погребной мастер решит, что новая бочка готова к розливу, подсоединяют линию розлива пива, которая соединяет бочку и устройство beer engine. Beer engine — это всего лишь причудливый ручной насос, который "тянет" пиво из бочки. По мере того, как пиво уходит из бочки, воздух просачивается через пористую втулку. По этой причине бочковые эли лучше всего в первые несколько дней розлива, и известны тем, что становятся все более и более непригодными для питья после трех или четырех дней. Окисленное пиво в любой стране не очень приятно, и бочки, которые простояли слишком долго, демонстрируют сильный эффект окисления. В попытке противостоять некоторым отрицательным последствиям окисления, пивовары и владельцы баров изобрели несколько методов введения СО2 в бочку. Наименее спорным является метод создания защитного слоя из углекислоты, когда в бочку впрыскивается чрезвычайно малое количество (от 1 до 2 psi) СО2 газа. Поскольку СО2 тяжелее воздуха, он образует "покров" над пивом, определенным образом защищая его от кислорода. Другой метод использует собственно емкости с СО2 для выталкивания пива, и для вытягивания пива из бочки также используются механические насосы. Приверженцы традиций презирают любые способы применения СО2 для сохранения качества пива, пытаясь убедить, что все они в результате приводят к получению какого-то "газированного" пива. Кампания за Риэл Эль (CAMRA) является наиболее непреклонной в том, чтобы риэл эль разливался только при помощи beer engine без применения "покровного" давления. С этой целью, CAMRA отказывается указывать в ежегодно издаваемой бесценной книге "Гид хорошего пива" (Good Beer Guide) те пабы, где используются СО2 системы. В то время как CAMRA так замечательно придерживается традиций, возможно, нереально будет ждать от самых маленьких пабов в самых удаленных местах, чтобы они могли заботиться о бочковых элях точно также, как это делается в более оживленных пабах. Если применяется бочковое охмеление,используют небольшое устройство типа сита, чтобы хмель оставался в бочке и не попадал в чей-нибудь стакан. На конце раздаточного крана используется распыляющее устройство, заставляющее пиво проходить через несколько небольших отверстий, что приводит к выбросу газа в пиво и стакан и к образованию густой пены. Бочковые эли также могут разливаться непосредственно из бочки самотеком. При таком способе бочку устанавливают таким образом, чтобы выпускное отверстие смотрело вниз. Открыв кран и воздушный вентиль, пиво будет литься из бочки самотеком. Если при данном методе используется сода-кег, погружную трубку для жидкости необходимо удалить или значительно обрезать. Это отличный способ розлива качественного пива, если пиво будет выпито за один вечер.
СОВЕТЫ ПИВОВАРАМ
Производство бочковых элей не особенно отличается от производства обычного домашнего пива. В обоих случаях, пиво газируется естественным способом в закрытом сосуде. Главное отличие в количестве образующейся карбонизации. Обычные эли и лагеры газируются приблизительно до 2.5 объемов СО2, в то время как для бочковых элей эта цифра приближается к 1.5–1.75 от объема. При изготовлении бочковых элей существует 2 способа, которым можно следовать: (1) Дайте брожению закончиться, затем добавьте небольшое количество сбраживаемого сырья. (2) Тщательно контролируйте брожение, и когда конечная плотность будет в пределах 1°, закупорьте бочку. Этот способ предпочтительней, но осуществить его может быть не так легко, потому что пивовар довольно точно должен знать, какой будет конечная плотность. Эта метод можно упростить, если использовать для розлива сода-кег из нержавеющей стали, и внимательно стравливать избыток давления, когда бочонок будет насыщаться газом. Использование осветлителей может стать еще одним дополнительным усилием, и многие домашние пивовары могут не захотеть с этим связываться. В этом случае, убедитесь, что Вы используете дрожжи, которые известны своими превосходными флоккуляционными свойствами. Если осуществляется бочковое охмеление, следует использовать только свежий шишковой хмель или цельный прессованный хмель (plugs). Поместите хмель в проницаемый мешок, используйте продезинфицированный груз, чтобы мешочек опустился на дно бочки. Если Вы собираетесь подавать пиво как настоящий бочковой эль, можно использовать подачу самотеком, но в идеале бочка должна быть опустошена за один вечер. Исторически, индийский пэйл эль имел значительно более высокую первоначальную плотность, но по своему опыту я знаю, что сегодня она снизилась до 1.035 — 1.045, и это просто более сильно охмеленная версия биттера. Крепкие эли можно найти даже со столь "низкой" первоначальной плотностью, как 1.051, но часто они имеют плотность в районе 1.062. Обратите внимание, что, несмотря на относительно низкое содержание спирта по объему у всех типов пива, кроме крепкого эля, это пиво имеет много несброженных веществ, которые придают бочковым элям богатое тело и полноту вкуса. Несброженные вещества состоят главным образом из декстринов, которые нормальные дрожжи не могут усвоить. Эти декстрины переходят в неизменном виде в готовое пиво. Использование карамельных солодов (обычно называемых кристаллическим солодом) всегда увеличивает в пиве количество несбраживаемых веществ. Карамельные солода обычно используются при производстве бочкового эля. Многие пивовары делают сусло с первоначальной плотностью 1.055 или выше и растворяют его в ферментере, добавляя кипяченную охлажденную воду, чтобы получить первоначальную плотность 1.035. Используя этот способ, определенная степень карамелизации может быть достигнута при варке в котле, что может положительным образом сказаться на вкусовых признаках.
СЛОВАРЬ ТЕРМИНОВ
Burton Unions — Классическая техника брожения, при которой дрожжевой оттек из ферментера собирается в сборном танке, а затем повторно возвращается назад в ферментер. Isinglass (рыбий клей) — оклеивающее вещество, изготавливаемое из переработанных плавательных пузырей рыбы, в основном осетра. При использовании твердого осветлителя isinglass, добавьте от 30 до 60 миллиграммов на литр. Перемешивайте одну минуту с обеспложенной водой, дайте постоять, снова перемешайте, и дайте отстояться, затем добавляйте. Нормы внесения меняются в зависимости от плотности, состава солодовой засыпи и технологического процесса. Проведите небольшой тест, чтобы определить производительность клея в вашей системе. Isinglass также используется в баках для созревания в количестве двух унций на один галлон или, грубо, в 2 процентной концентрации. Осветлитель (оклеивающие вещества, клей) — материалы, которые помогают осветлить эли бочкового созревания. Включает isinglass, желатин, а при необходимости удаления белка, алгинол (alginol) (отрицательно заряженные полисахариды). Остаточный экстракт — не следует путать с конечной плотностью, остаточный имеет отношение к количеству сбраживаемых веществ, оставшихся на определенный момент. Эти сбраживаемые вещества будут переработаны дрожжами во взвешенном состоянии по прошествии определенного времени. Стеллаж — деревянное устройство, используемое для установки на нем бочки, приготовленной для розлива. Конечная плотность — окончательная плотность пива, измеренная после того, как все сбраживаемые вещества были использованы дрожжами. Пиво верхового брожения — пиво, сброженное путем применения дрожжевых штаммов верхового брожения, Saccharomyces cerevesae. Дрожжи этого класса обычно сбраживают сусло, действуя у поверхности ферментера, и поднимаются на поверхность в конце брожения. Несбраживаемые вещества — это в основном декстрины, которые нормальные пивоваренные дрожжи не могут переработать. В результате, эти "бодибилдеры" (строители тела) переходят в готовое пиво и привносят сладость, тело и полноту вкуса. Несбраживаемые вещества можно грубо подсчитать по конечной плотности. Пиво, которое имеет первоначальную плотность 1.040 и конечную плотность 1.010, имеет 10/40 несброженных веществ или 25 процентов. Yorkshire Squares — Классические квадратные ферментеры, изготовленные из сланца, на пивзаводе Samuel Smith brewery. Yorkshire Squares состоят из чашевидных сланцевых устройств, которые расположены выше ферментеров, что позволяет собирать дрожжевой оттек для его удаления или, если его оставляют, для повторного внесения в ферментер.
Уксус. Что это такое и как его делают
Н. Кустова
Множество блюд в кухнях разных народов мира невозможно приготовить без уксуса. Не обойтись без него в заготовках, да и в качестве простой приправы уксус подают ко многим кушаньям. Изготовление уксуса, как и виноделие, один из древнейших технологических процессов, освоенных человеком. Но если в производстве вина за последние несколько тысячелетий не произошло принципиальных изменений (использование современного оборудования не в счет), то в производстве уксуса в 70-х годах XX века случилась настоящая революция. Начнем с того, что главным компонентом пищевого уксуса является уксусная кислота. Получать ее можно двумя способами: химическим — из продуктов сухой перегонки древесины и микробиологическим — в результате уксуснокислого брожения спиртосодержащих жидкостей, таких, как виноградное вино, сидр, пивное сусло, забродившие мед и соки различных фруктов или водный раствор этилового спирта (С2Н5ОН). В таких жидкостях окисление этанола до уксусной кислоты проводят в большинстве случаев уксусные бактерии Acetobacter aceti. В результате в готовом продукте оказывается не только кислота, но и небольшое количество сложных эфиров, альдегидов и других органических соединений. Именно благодаря этим веществам пищевой уксус обретает присущий ему особый вкус и приятный аромат. Разведенная же водой уксусная кислота, полученная химическим путем, лишена таких качеств. Считается, что в пищевой промышленности и в быту лучше использовать уксус, изготовленный биохимическим способом. Технология производства уксуса имеет интересную и сложную историю. Еще в первом тысячелетии до новой эры виноделы заметили, что, если вино оставить в открытом сосуде, оно через некоторое время прокисает и превращается в уксус. Этим наблюдением и пользовались долгое время, не вдаваясь особенно в суть того, что происходит при этом с продуктом. Один из самых "древних" способов производства уксуса принято называть орлеанским. В деревянные бочки особой формы, расположенные в утепленном помещении в несколько рядов одна над другой, в начале процесса заливают 10–12 л готового нефильтрованного уксуса. Эта порция — своего рода закваска, ведь в нефильтрованном уксусе содержится достаточно большое количество бактерий. К уксусу приливают примерно 10 л профильтрованного вина. Через восемь дней, если процесс идет нормально, доливают еще 10 л, и так до тех пор, пока бочка не заполнится до половины объема. После этого около 40 л готового продукта сливают, а к оставшемуся — вновь добавляют фильтрованное вино, и цикл повторяется[93]. Весь цикл занимает от недели до месяца, зато продукт обладает таким высоким качеством, что этот неэффективный способ до сих пор применяется в винодельческих районах Франции. Наряду с орлеанским способом существовал метод, описанный немецким ученым Бургавом (Boerhave) в 1732 году. Сейчас эта технология известна под названием "метод Шуценбаха". Суть его в том, что спиртосодержащую жидкость (в описании Бургава упоминается раствор хлебного спирта) пропускали сверху вниз через объем, заполненный тщательно вымоченными в уксусе крупными буковыми стружками. Эта технология оказалась значительно более производительной, чем орлеанский способ, и во всем мире она используется до сих пор[94]. И все же до работ Пастера в середине XVIII века не было понятно, за счет чего вино превращается в уксус. Пастер в большой статье "Исследование свойств уксуса" ("Etude sur le vinaegre") показал, что стерильный раствор спирта в воде на открытом воздухе практически не окисляется, а образование уксусной кислоты происходит благодаря работе уксусных бактерий. И для того, чтобы спирт окислялся эффективно, в жидкости необходимо создать оптимальные условия для их развития. Оказалось, что лучше всего эти микроорганизмы чувствуют себя при температуре около 30 °C и при концентрации спирта, не превышающей 12–14 %. Дальнейшие (уже современные) исследования показали, что максимальная скорость роста A.aceti достигается при более низкой концентрации спирта. Характерной особенностью этих бактерий является и высокая потребность в кислороде. Долгое время считалось, что из-за сравнительно низкой растворимости кислорода в воде (и в растворе этилового спирта тоже) бактерии могут развиваться только на поверхности жидкости или в ее тонкой пленке. Это не противоречило и имевшемуся к тому времени промышленному опыту. При орлеанском методе бактерии развиваются в основном в верхнем слое жидкости в виде слизистой пленки, а при методе Шуценбаха жидкость стекает тонким слоем по поверхности стружек (рис. 1). Производительность аппаратуры, что по одному, что по другому способу, обычно составляет от 2 до 8 кг 100 %-ной уксусной кислоты с 1 м3 объема аппарата в сутки.
Рис. 1. Аппарат Шуценбаха: 1 — деревянная коническая емкость; 2 — слой буковых стружек.
Основным аппаратом, в котором получают уксусную кислоту по методу Шуценбаха, является деревянный чан конической формы. На расстоянии 200–300 мм от основного днища в нем устанавливают горизонтальную перфорированную перегородку. Верхняя часть аппарата на 2/3 заполняется стружками, которые орошаются питательной для бактерий средой, содержащей некоторое количество уксусной кислоты (чаще всего это 6 %-ный раствор), этиловый спирт (3–4 %) и небольшое количество аммонийных и фосфатных солей. По мере протекания раствора бактерии, закрепившиеся, или, как теперь принято говорить, иммобилизованные на стружках, окисляют спирт в уксусную кислоту. В нижней части аппарата скапливается готовая продукция — 9 %-ный уксус. В процессе окисления выделяется тепло, которое повышает температуру внутри аппарата до 30–35 °C. В результате разницы температур создается естественная и довольно интенсивная конвекция. Воздух поступает в патрубки под ложным днищем, проходит через аппарат и выходит в верхней его части. Так сама собой осуществляется аэрация, необходимая для работающих бактерий. Несколько слов стоит сказать о стружках. Это не просто отходы от обработки древесины. Для загрузки в аппараты подходят только буковые стружки, закрученные в рулон диаметром от 2 до 5 см и высотой от 3 до 6 см. Серьезные требования предъявляются и к древесине. Она должна быть совершенно лишена любых видов гнили. Словом, стружки для уксусного производства — вещь совсем не дешевая. В аппарат Шуценбаха загружается 1–1,5 м3 стружек. На одном предприятии работают десятки таких аппаратов. Производительность аппаратуры при работе по данному способу низка и составляет не более 1,5 кг уксусной кислоты на 1 м3 стружек в сутки (в пересчете на 100 %-ную уксусную кислоту). При этом выход уксуса (от теоретически возможного при использовании исходного количества этилового спирта) не превышает 75 %. Процесс ведется непрерывно, десятилетиями, без смены бактерий и стружки. Высокая кислотность заливаемого в аппарат раствора необходима для того, чтобы другие бактерии не могли "заселить" аппарат и испортить таким образом продукт. Это дает возможность вести производство уксуса без соблюдения стерильности. Единственный спутник уксусных бактерий в этом процессе — мелкие нематоды — угрицы. Они питаются бактериями и тоже легко переносят высокие концентрации уксусной кислоты. Уксус очищают от них фильтрованием после пастеризации, в результате которой они погибают и выпадают в осадок. В настоящее время на подавляющем большинстве предприятий производство уксуса ведут циркуляционным способом Фрингса. Эта технология имеет немало общего с методом Шуценбаха. Здесь также используются аппараты, наполненные стружками, также на стружках иммобилизованы уксуснокислые бактерии, и также масса стружек орошается питательным раствором, содержащим спирт, уксусную кислоту и минеральные соли. Однако есть и существенные различия между этими методами. Прежде всего, это касается размера аппаратов. На некоторых предприятиях объем их заполненной стружками рабочей камеры достигает 60 м3. В такой аппарат (рис. 2) через специальную распределительную систему подают 10 %-ный раствор спирта со скоростью в несколько раз большей, чем по методу Шуценбаха. При помощи насоса раствор многократно циркулирует через аппарат до тех пор, пока весь спирт не окислится и не образуется 9 %-ный раствор кислоты. Около 10 % исходного чистого спирта в этом процессе теряется. Цикл длится 5–6 дней, после чего повторяется.
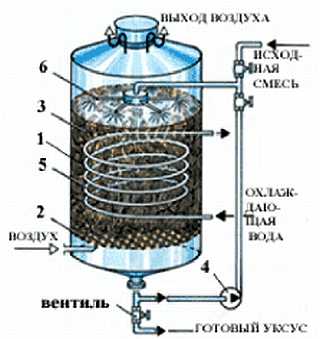
Рис. 2. Аппарат Фрингса: 1 — корпус; 2 — ложное перфорированное днище; 3 — слой буковых стружек; 4 — циркуляционный насос; 5 — змеевик системы термостатирования; 6 — распределительное устройство.
В аппаратах большого объема тепловыделение оказывается настолько значительным, что в них приходится встраивать специальные теплообменники. Чаще всего в рабочей камере располагают змеевики, по которым циркулирует охлаждающая вода, но иногда приходится устраивать еще и дополнительные, так называемые выносные теплообменники, которые устанавливают снаружи аппарата в циркуляционном контуре. При получении уксуса циркуляционным способом удельная производительность достигает 6–8 кг кислоты в сутки на 1 м3 рабочего объема аппарата. Но и у этого метода оказались существенные недостатки, главным из которых был, пожалуй, размер аппаратов. В начале шестидесятых годов XX века появилась технология, при которой уксуснокислые бактерии стали культивировать в специальных аппаратах — ферментерах в жидкости, — так называемый метод периодического глубинного культивирования. Ферментеры для глубинного культивирования уксусных бактерий — это изготовленные из нержавеющей стали емкости, внутри которых размещаются перемешивающие устройства и аэраторы различных конструкций (рис. 3).
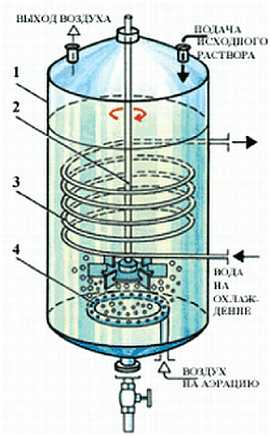
Рис. 3. Схема ферментера для производства уксуса: 1 — корпус из нержавеющей стали; 2 — перемешивающее устройство; 3 — аэратор (его обычно называют барботером); 4 — змеевик системы термостатирования.
Процесс получения уксуса при периодическом глубинном способе заключается в следующем. От предыдущего цикла в аппарате остается жидкость (примерно 1/3 рабочего объема аппарата), которая служит посевным материалом для следующего цикла. В аппарат заливается до рабочего объема питательная смесь, содержащая уксусную кислоту и этанол. Перемешивающее устройство интенсивно перемешивает жидкость, а через аэратор непрерывно подается воздух. В начале цикла условия жизни для бактерий резко меняются, и в результате некоторое время не наблюдается их заметного роста, эта стадия в развитии микроорганизмов называется лаг-фазой. По окончании лаг-фазы концентрация спирта начинает уменьшаться, а кислоты — наоборот, расти. Некоторое время в аппарат приходится порциями добавлять раствор спирта. После того как концентрация уксуса достигает 9-10 %, около 2/3 объема жидкости отбирается как готовый продукт, и цикл повторяется. Производительность глубинных аппаратов в несколько раз выше, а сами они в несколько раз меньше, чем аппараты, заполненные стружками, в них значительно меньше потери этанола. Кроме того, отпадает необходимость применения древесных стружек. Немаловажно и то, что при глубинном способе возрастает культура производства. В начале 70-х годов прошлого столетия у группы сотрудников кафедры "Машины и аппараты микробиологических производств" в Московском институте химического машиностроения (теперь это Московский государственный университет инженерной экологии), возглавляемой профессором Петром Ивановичем Николаевым, возникла идея совместить в промышленном масштабе микробиологические методы с приемами постановки и ведения процессов, хорошо отработанными в химической технологии. Для этого пришлось провести целый комплекс серьезных исследований. Вот ведь парадокс: процесс был известен уже как минимум два с половиной тысячелетия, но до середины XX века оставался в основном эмпирическим. До этого момента усовершенствования технологий касались, прежде всего, устройства аппаратов, а микробиологические аспекты разрабатывались весьма слабо. В 60-е годы стали появляться работы, посвященные физиологии и биохимии уксусных бактерий. Они были направлены на изучение влияния концентрации кислорода и состава питательной среды, включая как минеральный фон, так и влияние этанола и самой уксусной кислоты. В это же время на кафедре микробиологии Ленинградского университета под руководством профессора М. С. Лойцянской были проведены исследования систематики, морфологии и физиологии этих бактерий. Были выделены штаммы бактерий, растущих в очень простой по составу среде, обладающей большой окислительной активностью, что оказалось необычайно полезно для промышленного производства уксуса. Оптимальная температура для роста Acetobacter aceti — 25–30 °C. В качестве источника азота уксуснокислые бактерии используют минеральные соли, предпочтительно аммонийные. Ацетобактеры сами синтезируют все необходимые витамины и поэтому растут в питательных средах без их добавления. Лучшим соединением углерода для бактерий рода Acetobacter является уксусная кислота. Хорошо растут они также в средах, содержащих этиловый спирт или молочную кислоту, превращая их в уксусную. Исследованиями Ю. Л. Игнатова было показано, что накапливаемая в процессе уксусная кислота снижает окислительную активность бактерий и уменьшает удельную скорость роста клеток. Этот факт позволил П. И. Николаеву с сотрудниками организовать процесс получения уксусной кислоты в батарее из нескольких аппаратов глубинным способом в непрерывном режиме. В результате получилась оригинальная технологическая схема, в которой процесс получения 9 %-ной уксусной кислоты ведут в четырех-пяти последовательно соединенных ферментерах (рис. 4). В такой батарее, в первых двух, по ходу жидкости, аппаратах при сравнительно низкой концентрации уксусной кислоты бактерии размножаются с большой скоростью при высокой окислительной активности, что обеспечивает высокую продуктивность процесса. В последних по ходу жидкости аппаратах, работающих, напротив, при высоких концентрациях уксусной кислоты, продуктивность снижается, в них происходит в основном доокисление оставшегося в растворе спирта. Общая производительность всех аппаратов батареи значительно выше, чем одного, выпускающего уксус 9 %-ной концентрации. Ю.Л. Игнатов показал, что производительность единицы рабочего объема аппарата, работающего по батарейному способу, может достигать 4 9,4 кг уксусной кислоты с 1 м3 в сутки. Разработанный способ был на удивление быстро внедрен на нескольких заводах. Сейчас по этой технологии работают Экспериментальный пищекомбинат в Балашихе, уксусные цеха в городах Горловка и Днепродзержинск на Украине, завод в Словакии.

Рис. 4. Схема установки для получения уксуса в непрерывном режиме. Переток жидкости из аппарата в аппарат происходит из-за разницы давлений в "воздушной подушке", возникающей за счет разного заглубления переточных труб h: h2 > h3 > h4 > h5.
Краткие сведения о химизме окисления этанола в уксусную кислоту Acetobacter aceti Итоговая реакция окисления этилового спирта в уксусную кислоту выглядит следующим образом: Acetobacter aceti С2Н5ОН —> СН3СООН + Н2О + Q По современным представлениям, окисление этилового спирта уксуснокислыми бактериями вида Acetobacter aceti — двухфазный процесс. Этанол окисляется алкоголь- и альдегиддегидрогеназами с образованием уксусной кислоты и двух молекул НАДН2. (Этот фермент отвечает за перенос водорода в дыхательной цепи.) Алкогольдегидрогеназа Acetobacter aceti содержит недавно открытую простетическую группу метоксантин, или пирролохинолинхинон. Этот фермент находится на внешней стороне плазматической мембраны и катализирует окисление этанола в уксусную кислоту. Метоксантин частично попадает в питательную среду и в пищевой уксус, придавая ему слегка желтоватую окраску.
ХИМИЧКА
Опыты в домашней лаборатории

ТЕХНИКА БЕЗОПАСНОСТИ
Этот раздел был создан исключительно для вашей безопасности, поэтому прочтите его, пожалуйста. Все опыты полностью безопасны, если у вас есть достаточные химические знания. Так что перед работой обязательно проанализируйте описание эксперимента, просмотрите примечания про технику исполнения и правила безопасности. В случае необходимости обратитесь за консультацией к учителю или почитайте дополнительную литературу. Можете быть уверены, что при точном выполнении приведённых ниже советов вы не получите травм или отравлений при работе с химическими веществами. Итак, вот основные и самые важные правила: 1) НИКОГДА не пробуйте на вкус и не нюхайте химические вещества! 2) При работе с открытым пламенем соблюдайте правила противопожарной безопасности. 3) Если вы хотите добавить спирт в спиртовку, то её обязательно необходимо потушить перед доливанием. 4) Если вам необходимо сделать раствор кислоты, то необходимо лить кислоту в воду, а не наоборот. Для того чтоб запомнить это, существует простенький стишок:
Сначала вода, потом кислота,
иначе случится большая беда!
Несколько полезных советов. Вот несколько советов, которые могут вам понадобиться для проведения многих опытов: 1) Во время работы с различными солями желательно надеть одноразовые резиновые перчатки, т. к. кожа имеет свойство впитывать вещества, попавшие на неё. 2) Наиболее токсичными являются соли следующих металлов: Hg, Pb, Cd, Со, Ni, Zn, Ва, Sb, Sr, Сr; менее токсичны соли Си, Fe, Аl, Аg; и наименее токсичны соли Са, К, Na и Мg. Но работать необходимо осторожно со всеми веществами. 3) Я советую завести специальное полотенце (предпочтительно вафельное) для вытирания вымытой химической посуды. 4) Если после кипячения раствора на стенках химической посуды остался несмываемый налёт, то его можно удалить, ополоснув ёмкость раствором соляной кислоты. 5) Если для проведения эксперимента вам необходим пробиркодержатель, а у вас его нет, то его можно сделать из обычной деревянной (не пластмассовой!) прищепки.
ГДЕ ВЗЯТЬ ХИМИЧЕСКИЕ РЕАКТИВЫ ДЛЯ ОПЫТОВ?
Если вы по каким-то причинам не можете заказать специальные химические реактивы, попробуйте найти их среди товаров народного потребления (на примере Москвы): Азотная кислота — на Митинском радиорынке. Алюминий — на строительном рынке, в пунктах приема металлолома Аммиак водный (нашатырный спирт) — в аптеке. Ацетат свинца — в аптеке. Ацетон (диметилкетон) — в хозмаге среди растворителей. Бензин — в хозмаге среди растворителей, на автозаправочной станции. Борная кислота — в аптеке. Висмут — в припоях, особенно легкоплавких. Гексаметилентетрамин (уротропин, сухой спирт) — в туристическом магазине. Гидрокарбонат натрия (сода) — в продовольственном магазине. Гидроксид калия (КОН) — на Митинском радиорынке. Гидроксид кальция (гашеная известь[95]) — на строительном рынке, в крупном хозмаге. Гидроксид натрия (едкий натр) — в хозмаге продается как средство для прочистки труб "Крот". Гидропирит — в аптеке. Глицерин[96] — на радиорынке, в магазинах радиодеталей. Глюкоза — в аптеке. Графит — в простых карандашах, на радиорынке. Диоксид кремния (кварцевый песок) — на строительном рынке. Дистиллированная вода — в аптеке. Дихлорэтан — в хозмаге (клей для оргстекла). Дихромат калия К2Сr2O7 — используется как оранжевая краска, продаётся в виде порошка под названием "хромпик". Йод — в аптеке. Желатин — в продовольственном магазине. Золото — в ювелирных ларьках[97], в магазинах сырья для ювелиров. Карбамид (мочевина) — в хозмаге, в отделе удобрений. Карбонат кальция (мел) — в канцтоварах, на строительном рынке. Карбонат натрия (стиральная сода, кальцинированная сода) — в хозмаге. Кевлар (нить) — в рыболовном магазине. Крахмал — в продовольственном магазине. Интерферон — в аптеке. Изопропанол (баллончик для очистки дисплеев и контактов) — в крупных компьютерных и радиомагазинах. Лимонная кислота — в продовольственном магазине. Марганцевокислый калий KMnO4 — марганцовка, продаётся в аптеках. Медь — в пунктах приема металлолома, в электротехнических изделиях Нафталин — в хозмаге (средство от моли). Нитрат аммония (аммиачная селитра) — в хозмаге, среди удобрений. Нитрат калия KNO3 (калиевая селитра) и нитрат натрия NaNO3 (натриевая селитра) — удобрения, продающиеся в с/х магазинах. Нитрат кальция (кальциевая селитра) — в хозмаге, среди удобрений. Нитрат серебра (ляписный карандаш) — в аптеке. Оксид кальция (негашеная известь) — в хозмаге, на строительном рынке. Оксид магния (жженая магнезия) — в аптеке. Оксид хрома(III) Сr2O3 — очень устойчивая зелёная краска, называется "хромовый зелёный". Продаётся в хоз. магазинах. Олово — в припоях. Ортофосфорная кислота — на Митинском радиорынке. Парафин — в хозмаге, в свечках. Перманганат калия (марганцовка) — в аптеке. Пероксид водорода (перекись водорода) — в аптеке. Персульфат аммония — на радиорынке, в магазинах для любителей традиционной (нецифровой) фотографии. Порох — в охотничьих магазинах, петардах и фейерверках. Резорцин — в аптеке. Сахароза (сахар) — в продовольственном магазине. Сера S — продаётся в виде порошка и серных шашек. Порошок применяется для опрыскивания растений, а шашки для выкуривания насекомых-вредителей. Серебро — в ювелирных ларьках, в магазинах сырья для ювелиров. Свинец[98] — в пунктах приема металлолома, в автомобильных аккумуляторах Серная кислота — в автомагазине. Силикат натрия Na2SiO3 (метасиликат) — в хозмаге, на строительном рынке — это силикатный клей, продающийся в магазинах канцелярских принадлежностей. Часто его продают на разлив под названием "жидкое стекло". Слюда — на Митинском радиорынке. Соляная кислота — на Митинском радиорынке. Стеарат натрия (хозяйственное мыло) — в хозмаге. Сульфат железа FeSO4*7Н2О (железный купорос) — в хозмаге, с/х магазинах. Сульфат меди CuSO4*5Н2O (медный купорос) — в хозмаге, среди средств по борьбе с плесенью. Сульфит натрия — в магазинах для любителей традиционной (нецифровой) фотографии — проявитель. Тетраборнокислый натрий (бура) Nа2В4O7*10Н2O — это вещество используется для пайки. Купить его можно на радиорынке. Тимол — в аптеке. Ферроцианид калия (желтая кровяная соль) — в магазинах для любителей традиционной (нецифровой) фотографии. Фоторезист — на радиорынке, в магазинах радиодеталей. Фторопласт — на Митинском радиорынке. Уайт-спирит — в хозмаге. Уголь активированный — в аптеке. Уголь древесный — в хозмаге. Уголь каменный — на строительном рынке. Уксусная кислота — в продовольственном магазине. Фенолфталеин — входит в состав некоторых слабительных препаратов, таких, как пурген и фенолфталеин. Продаётся в аптеках. Формиат натрия — в хозмаге или на строительном рынке — это жидкость для отопительных систем. Хлорид аммония — на радиорынке. Хлорид-гипохлорит кальция (хлорка) — в хозмаге. Хлорид железа (хлорное железо) — на радиорынке, в радиомагазине. Хлорид кальция — в аптеке. Хлорид натрия (поваренная соль) — в продовольственном магазине. Целлюлоза (вата) — в аптеке. Цинк — в батарейках (особенно советских). Этиленгликоль — в автомагазине — это антифриз[99]. Этиловый спирт С2Н5OН — лучше всего покупать его в аптеках.
Лабораторная посуда 1.
 Стакан с носиком служит для хранения жидких и твердых веществ, а также для проведения простейших химических операций (растворение, нагревание).
3.
Стакан с носиком служит для хранения жидких и твердых веществ, а также для проведения простейших химических операций (растворение, нагревание).
3.  Колба коническая используется для хранения жидких и твердых веществ, а также для проведения различных химических операций.
4. Колба круглодонная служит для проведения разнообразных химических операций при нагревании.
5. Колба с отростком предназначена для перегонки и разделения жидкостей при нагревании.
6. Колба плоскодонная служит для проведения химических операций, а также для хранения жидких и твердых веществ.
7.
Колба коническая используется для хранения жидких и твердых веществ, а также для проведения различных химических операций.
4. Колба круглодонная служит для проведения разнообразных химических операций при нагревании.
5. Колба с отростком предназначена для перегонки и разделения жидкостей при нагревании.
6. Колба плоскодонная служит для проведения химических операций, а также для хранения жидких и твердых веществ.
7.  Мерная посуда: цилиндр (рис), стакан и пробирка — используют для измерения объема жидкостей.
8.
Мерная посуда: цилиндр (рис), стакан и пробирка — используют для измерения объема жидкостей.
8. Воронка конусообразная служит для переливания жидкостей и фильтрования.
9.
Воронка конусообразная служит для переливания жидкостей и фильтрования.
9. Стеклянная палочка предназначена для размешивания веществ в химической посуде. Для предохранения посуды от случайного растрескивания при размешивании веществ на конец стеклянной палочки надевают кусочек резиновой трубки.
10.
Стеклянная палочка предназначена для размешивания веществ в химической посуде. Для предохранения посуды от случайного растрескивания при размешивании веществ на конец стеклянной палочки надевают кусочек резиновой трубки.
10.  Ложка фарфоровая (1), шпатель (2) служат для взятия твердых и сыпучих веществ. Ложка-дозатор (3) предназначена для взятия определенной порции вещества.
11.
Ложка фарфоровая (1), шпатель (2) служат для взятия твердых и сыпучих веществ. Ложка-дозатор (3) предназначена для взятия определенной порции вещества.
11.  Чашка фарфоровая применяется для выпаривания жидкостей.
12.
Чашка фарфоровая применяется для выпаривания жидкостей.
12.  Тигель фарфоровый предназначен для нагревания и прокаливания твердых веществ при высокой температуре.
13.
Тигель фарфоровый предназначен для нагревания и прокаливания твердых веществ при высокой температуре.
13.  Треугольник фарфоровый используется для размещения в нем тигля. Треугольник помещают на кольцо штатива.
14.
Треугольник фарфоровый используется для размещения в нем тигля. Треугольник помещают на кольцо штатива.
14.  Ступка с пестиком служат для размельчения и растирания твердых веществ.
15.
Ступка с пестиком служат для размельчения и растирания твердых веществ.
15.  Штатив для пробирок, служит для размещения в нем пробирок.
16.
Штатив для пробирок, служит для размещения в нем пробирок.
16.  Зажим пробирочный служит для закрепления пробирки, если вещество в пробирке требуется нагреть в пламени.
17.
Зажим пробирочный служит для закрепления пробирки, если вещество в пробирке требуется нагреть в пламени.
17.  Зажим пружинный используют для зажимания резиновых трубок при монтаже различных приборов.
18.
Зажим пружинный используют для зажимания резиновых трубок при монтаже различных приборов.
18.  Промывалка служит для промывания осадков дистиллированной водой, для смывания осадков с фильтров и стенок сосудов. Ее используют также для хранения небольших количеств дистиллированной воды.
19.
Промывалка служит для промывания осадков дистиллированной водой, для смывания осадков с фильтров и стенок сосудов. Ее используют также для хранения небольших количеств дистиллированной воды.
19.  Воронка делительная предназначена для разделения несмешивающихся жидкостей, имеющих разную плотность.
20.
Воронка делительная предназначена для разделения несмешивающихся жидкостей, имеющих разную плотность.
20.  Склянка с пипеткой служит для хранения растворов реактивов при работе с малыми количествами веществ.
21. Пластина для капельного анализа (керамическая или стеклянная) используется при выполнении опытов с очень малыми количествами веществ в объеме 1–2 капель.
22.
Склянка с пипеткой служит для хранения растворов реактивов при работе с малыми количествами веществ.
21. Пластина для капельного анализа (керамическая или стеклянная) используется при выполнении опытов с очень малыми количествами веществ в объеме 1–2 капель.
22.  Прокладка керамическая огнезащитная используется при нагревании веществ в стеклянной посуде. Прокладку размещают на кольце металлического штатива.
Прокладка керамическая огнезащитная используется при нагревании веществ в стеклянной посуде. Прокладку размещают на кольце металлического штатива.
ПРИБОРЫ ДЛЯ ПОЛУЧЕНИЯ ГАЗОВ
(В.Н.Верховский "Техника Химического Эксперимента")Приборы периодического действия Вот простейший из вышеуказанных приборов, состоящий из банки с жидкостью и пробирки с веществом. Пробирка держится в пробке, вставленной в горло склянки, и может быть погружена в жидкость. На конце пробирки сделано отверстие таких размеров, чтобы через него не могли проскочить помещенные в пробирку кусочки твердого вещества. На дно пробирки кладут несколько кусков стекла. Через отверстие в пробирку входит налитая в банку жидкость. Газ выходит через присоединенную к пробирке газоотводную трубку. Чем большее сопротивление должен преодолеть газ, тем глубже должна быть погружена в жидкость пробирка. Отверстие в дне пробирки можно сделать при помощи напильника, точильного камня или карборундового бруска, спилив самую нижнюю часть дна (держать напильник перпендикулярно к оси пробирки). Но лучше «продуть» отверстие, нагрев дно пробирки на остром пламени паяльной горелки или оттянуть конец пробирки и обрезать его. В последнем случае вместо пробирки можно воспользоваться трубкой подходящего диаметра.

Пробка, вставленная в пробирку, должна хорошо держать. Пробка же, в которой укреплена пробирка, наоборот, не должна прилегать слишком плотно к горлышку банки, чтобы жидкость в банке имела сообщение с атмосферой. Можно прорезать в пробке сбоку продольную щель. Если пробирка плохо держится в отверстии пробки и съезжает вниз, ее можно укреплять при помощи небольшого клина из спички и т. п. Вместо пробирки можно воспользоваться сеткой из такого материала, на который не действует жидкость, служащая для получения газа. Например, при получении оксида углерода СO2 взаимодействием мрамора с разбавленной соляной кислотой берут медную сетку, делают из нее мешочек или корзиночку (рис. 2), которую подвешивают на крючке из проволоки или стеклянной палочки. Крючок можно передвигать в пробке, вставленной в банку с жидкостью. Газ выходит через газоотводную трубку, находящуюся в той же пробке.
Такие приборы позволяют регулировать ток газа. Это имеет большое преимущество, особенно если требуется ровная струя газа или во время опыта необходимо изменять силу тока газа, а также если газ бывает нужен в определенные промежутки времени. Пока газ не нужен, реакция не идет и вещества не расходуются без надобности. Простейший небольшой прибор такого рода отличается от данного на рисунке 1 прибора тем, что снабжен краном или зажимом на каучуке. При закрывании крана образующийся газ вытесняет жидкость из пробирки, и реакция прекращается. Таким образом, для прекращения реакции нет необходимости поднимать пробирку. (В пробке должен быть вырез 1.) Чтобы прибор хорошо действовал, конец пробирки с твердым веществом лучше оттянуть подлиннее. Кусочки вещества не должны быть слишком мелкие, чтобы они не проваливались глубоко, в оттянутый конец пробирки, иначе после закрывания крана жидкость будет касаться твердого вещества и газ будет непрерывно выходить из пробирки. Если пробирка хорошо оттянута и куски твердого вещества достаточно крупные, то между твердым веществом и жидкостью будет оставаться слой газа, который и будет препятствовать их соприкосновению, пока прибор не работает. В нижнюю часть пробирки можно положить куски стекла для лучшего разделения твердого вещества от жидкости в то время, когда кран закрыт. Обычно после закрывания крана некоторое количество газа выходит наружу (пробулькивает), пока идет реакция между твердым веществом и не успевшей стечь с него жидкостью. Если прибор не используют долгое время, пробирку с твердым веществом можно поднять, как в приборе, изображенном на рисунке 4.
Нижнее отверстие пробирки с твердым веществом должно быть достаточно велико (2–3 мм), так как через него во время действия прибора проходят два тока жидкости: вверх — реагирующая жидкость, вниз — более тяжелый раствор, содержащий продукты реакции между жидкостью и твердым веществом (обычно раствор соли). На рисунке 4 изображен приборчик для получения газов. В одно из колен U-образной трубки в корзиночке из медной сетки помещены кусочки твердого вещества (цинк, мрамор и др.). В трубку наливают разбавленную кислоту. Когда зажим закрыт, прибор не действует. В качестве автоматического прибора можно использовать склянку Тищенко. Кусочки твердого вещества должны быть в этом случае достаточно мелки, чтобы они проходили в узкое горлышко склянки. Для наполнения кислотой склянку наклоняют почти горизонтально и кислоту наливают через воронку с узким, оттянутым и изогнутым концом, который входил бы в боковой отросток склянки. Зажим при этом должен быть закрыт. Можно также сначала налить в склянку через горлышко кислоту, затем присоединить к отростку склянки каучук, через который ртом высасывать воздух, пока кислота не перейдет в левую половину склянки, после чего зажать каучук (пальцами или зажимом), всыпать через горлышко цинк, вставить в склянку пробку и снять каучук с отростка. Склянку Тищенко следует выбрать с возможно малым отверстием в перегородке. Большое преимущество такого прибора — устойчивость и простота обращения при работе. Для лабораторных опытов учащихся очень удобны небольшие автоматические приборы, имеющиеся в продаже, а также самодельные — из большой пробирки или широкой стеклянной трубки (рис. 5).

Твердое вещество (цинк, мрамор и пр.) помещают в этих приборах на резиновый кружок с вырезами (рис. 5), надетый на трубку воронки и свободно входящий в пробирку (кусочки твердого вещества не должны проваливаться в вырезы и в зазор между кружком и стенками реакционного сосуда). Жидкость наливают через воронку при открытом (винтовом) зажиме в таком количестве, чтобы слой ее над твердым веществом был высотой 2–3 см. При перекрытии резиновой газоотводной трубки зажимом жидкость давлением газа вытесняется в воронку, стекает с твердого вещества через вырезы в резиновом кружке, и реакция прекращается. Для возобновления ее достаточно открыть зажим. Все эти приборы употребляют главным образом для получения небольших количеств газов при лабораторных опытах учащихся. Чтобы получать большие количества газа, который выходил бы из прибора сильной струей, пользуются общеизвестными приборами Сен-Клер-Девиля и Киппа. Прибор Сен-Клер-Девиля изображен на рисунке 6.
Он состоит из двух склянок с тубулусами (в 1–4 и более литров), соединенных каучуковой трубкой. В одной склянке находится твердое вещество, в другую наливают жидкость. Склянку с жидкостью помещают выше склянки с твердым веществом, но жидкость попадает к твердому веществу только в том случае, если открыт кран или зажим на газоотводной трубке и жидкость может вытеснить изсклянки с твердым веществом находящийся в ней газ (при начале работы — воздух). После закрывания крана или зажима образующийся газ вытесняет жидкость обратно в стоящую выше склянку, и реакция прекращается. Чем выше поставлена склянка с жидкостью относительно склянки с твердым веществом, тем сильнее может быть струя газа и тем большее сопротивление она может преодолеть. Возможность получать значительное давление газа составляет большое преимущество прибора Сен-Клер-Девиля по сравнению с большинством других приборов для получения газов при взаимодействии жидкости твердым веществом. При устройстве прибора Сен-Клер-Девиля на дно склянки, предназначаемой для твердого вещества, насыпают сначала слой битого стекла, фарфора, стеклянных бус, крупного гравия, черепков от глиняной посуды и т. п., а затем уже насыпают твердое вещество, участвующее в реакции получения газа. Насыпание индиферентного вещества имеет еще то преимущество, что после окончания работы, когда главная масса жидкости вытеснена в верхнюю склянку, часть нижней склянки, заполненная стеклом и т. п., представляет собой резервуар, наполняющийся газом, который продолжает выделяться вследствие действия на твердое вещество, еще не успевшей стечь с него, части жидкости. Чем больше слой индиферентного вещества, тем меньше газа может выйти из прибора через соединительную трубку и через слой жидкости в верхней склянке. Газ выходит обыкновенно большими пузырями, что сопровождается сильным бульканьем, отвлекающим внимание учащихся от опыта. Чтобы _ избежать этого, следует не сразу закрывать кран, а несколько раз приоткрывать его, чтобы выпустить избыток газа, если позволяют условия опыта. При прохождении пузырей газа через слой жидкости в верхней склянке из нее могут вылетать брызги жидкости, поэтому в склянку с жидкостью вставляют небольшую воронку, которая служит также и для наливания свежей жидкости. Пробки к нижним тубулусам склянок для аппарата Сен-Клер-Девиля должны быть обязательно резиновые, их необходимо привязывать. Длина каучуковой трубки, соединяющей склянки большого объема, 50–60 см, наружный диаметр около 1 см, а толщина стенок 1,5–2 мм. Стеклянные трубки, вставленные в нижние пробки и служащие для надевания соединяющего склянки каучука, должны быть хорошо оплавлены, концы их могут выставляться из пробок не более чем на 3–4 см. Соединительную резиновую трубку следует к ним привязать, для чего на концах стеклянных трубок желательно сделать небольшие перетяжки, как на соединительных и переходных трубках. Для регулирования выхода газа применяют стеклянный кран, согнув одну из трубок его под прямым углом, но можно удовольствоваться и винтовым зажимом. Пока прибор не нужен, склянку с кислотой ставят ниже склянки с твердым веществом: прибор в таком положении может храниться долгое время.
Для удобства пользования и хранения полезно сделать для прибора Сен-Клер-Девиля деревянный ящик, открытый с одной стороны. При хранении прибора можно склянку с жидкостью ставить внутрь ящика, а с твердым веществом — наверх. В таком виде прибор занимает мало места. При снаряжении прибора твердого вещества обыкновенно накладывают побольше, чтобы не приходилось очень часто переснаряжать его (но следует иметь в виду, что, чем больше твердого вещества, тем больше будет выделяться газа после закрывания крана). Жидкости наливают не более половины склянки. Если образующийся газ горюч, то каждый раз после добавления в прибор твердого вещества вытесняют из прибора весь воздух, чтобы не образовалась взрывчатая смесь. При отсутствии склянок с тубулусами можно сделать прибор Сен-Клер-Девиля из обычных склянок (бутылок) достаточного объема (рис. 8).
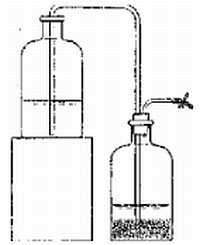
В таком приборе жидкость переливается в реакционную склянку с помощью сифонной трубки и вытесняется в верхний сосуд давлением газа при перекрывании! газоотводной трубки зажимом.
Аппарат Киппа Широко используется аппарат Киппа(рис. 10 и 11)


Твердое вещество помещают в средней части сосуда, отделенной от нижней перетяжкой, благодаря чему оно не соприкасается с остающейся на дне прибора жидкостью, когда прибор отключен. Нижняя часть сосуда в приборе Киппа играет ту же роль, что и пространство между кусками индиферентного вещества в приборе Сен-Клер-Девиля. Воронка вставлена в прибор на шлифе, газоотводная трубка с краном или зажимом — на пробке, лучше каучуковой. Нижний тубулус служит для выливания отработавшей жидкости. Твердое вещество вносят через тубулус. Чтобы твердое вещество не проваливалось вниз, отверстие между средним шаром и нижней частью прибора закрывают кружком из твердой резиновой пластины. В этом кружке прорезают широкое отверстие посередине для трубки воронки прибора и несколько небольших отверстий, через которые свободно могла бы проходить жидкость. Иногда приборы Киппа делают с соответствующим приспособлением из стекла в виде вставленной в перетяжку сосуда широкой трубки с отверстиями (рис. 11). Эта трубка, вставленная в перетяжку сосуда, должна иметь достаточно большие отверстия, расположенные не слишком высоко. В противном случае кислота не успевает стекать в нижнюю часть прибора или не имеет возможности стечь, и реакция продолжается еще долгое время после закрытия крана. Выделяющийся газ при этом проходит в нижнюю часть прибора и через трубку воронки пробулькивает в верхний шар большими пузырями, вызывая напрасный расход реактивов и разбрызгивание кислоты из отверстия (рекомендуется поэтому всегда держать в верхнем отверстии шара небольшую воронку). Если вставная трубка имеет очень маленькие или слишком низко расположенные отверстия, то лучше удалить ее совсем, заменив резиновым или пластмассовым кружком, или вставить в отверстие перетяжки свернутую в трубку пластмассовую сетку, из которой обычно делают сейчас прокладки для пластинок свинцовых аккумуляторов. Прежде чем снаряжать прибор, следует смазать вазелиновой мазью все пришлифованные части. Нижнюю и верхнюю (с краном) пробки необходимо привязать медной проволокой. Очень удобны для этого тонкие электрические многожильные провода в пластмассовой изоляции. Если пробки притертые, то сначала обматывают в 3–4 витка провод у толстого ранта тубулуса, наложив провод так, чтобы остались два одинаковых длинных конца. Эти концы скручивают, перекидывают скрученную часть на головку пробки, охватывая ее проводом с двух сторон, снова скручивают концы и перекидывают провод к ранту тубулуса, охватывая горло его с двух сторон, и опять скручивают концы. Во избежание потери времени на развязывание и привязывание пробки, что приходится делать при каждой перезарядке, удобнее сделать для нее запор, изображенный на рисунке 12.

Для устройства этого запора горло тубулуса обвивают проволокой, на которой делают две петли — одна против другой. В одну петлю вставляют конец длинной медной или лучше алюминиевой пластинки и огибают ее вокруг проволоки так, чтобы полоска могла откидываться на петле, как на шарнире. Пластинка должна быть вырезана и выгнута так, как показано на рисунке, чтобы она охватывала пробку с обеих сторон двумя боковыми отростками, оставленными посередине пластинки. Свободный конец пластинки выгибают в виде крючка, который может быть притянут ко второму сделанному на проволоке кольцу при помощи колечка, отрезанного от каучуковой трубки. Проволоку и пластинку лучше всего взять алюминиевые. Тогда приспособление будет служить много лет. Придется только иногда менять резиновое колечко.
ПРИЕМЫ ОБРАБОТКИ СТЕКЛА И ПРОБОК
Резание стеклянных трубок Чтобы отрезать кусок трубки определенной длины, нужно сначала отметить место надреза, взять трубку в левую руку, зажать ее между указательным и большим пальцами и ножом для резки стекла или трехгранным напильником сделать царапину. После этого, держа трубку обеими руками так, чтобы царапина приходилась между ними, слегка согнуть трубку со стороны, противоположной царапине. На месте царапины трубка обычно ломается. Если при небольшом усилии этого не произойдет, царапину несколько расширить и растягивание повторить. После того как трубка отрезана, концы ее следует оплавить. Для этого внести конец трубки наклонно в несветящееся пламя горелки, все время поворачивая трубку вокруг оси до тех пор, пока конец не накалится. Затем вынуть трубку из пламени и положить для остывания на сетку с асбестом (но не на стол!)
Сгибание стеклянных трубок Надеть на горелку насадку («ласточкин хвост»), зажечь горелку и равномерно нагреть место сгиба (см. рисунок), спокойно вращая трубку вокруг оси. Когда трубка размягчается, вынуть ее из пламени и плавно согнуть. Правильность сгиба зависит от равномерности нагревания и плавности сгибания. При быстром сгибании трубка будет иметь на сгибе впадину сверху или выемку снизу, что недопустимо.
Правильное и неправильное пользование пробками и трубками: 1 — резание трубки; 2 — правильно согнутая трубка; 3 и 4 — неправильно согнутые трубки; 5 — нагревание трубки в пламени "ласточкиного хвоста"; 6 — трубка, согнутая под прямым углом; 7 — вытягивание трубки.
Вытягивание стеклянных трубок Довольно часто требуется оттянуть трубку или же вытянуть капилляр. Для этого трубку надо нагреть на протяжении 4–5 см в пламени горелки, все время вращая трубку вокруг оси. После размягчения оттянуть ее до нужного диаметра. Если нужно получить капилляр (очень тонкую трубку с маленьким диаметром), то нагретую до размягчения трубку вынимают из пламени и быстро, но плавно растягивают обеими руками в оба конца.
Подбор пробок В лаборатории употребляют три сорта пробок: корковые, резиновые и стеклянные. Выбирать корковую пробку следует такого размера, чтобы диаметр ее был немного больше внутреннего диаметра горла сосуда. Затем пробку надо вложить в пробочный пресс и поворачивая, обжимать до тех пор, пока она не станет эластичной и будет плотно входить в горло сосуда. Держа левой рукой горло сосуда у отверстия правой осторожно вставляют пробку в горло, вращая и как бы ввинчивая ее. Пробка должна плотно входить в отверстие.
Сверление пробок Чтобы просверлить в пробке отверстие, надо сделать на суженной части обжатой пробки отметку той стеклянной трубкой, для которой делается отверстие. Затем подобрать пробочное сверло (рис.) несколько меньшего диаметра, чем наружный диаметр отметки. Сверлить пробку в горизонтальном положении, держа ее в левой руке, а сверло — в правой. Сверлить лучше всего следующим образом: слегка надавливая правой рукой на сверло, делать полуобороты сверлом вправо и влево. Когда сверло войдет в пробку почти до самого края, положить ее на дощечку или на другую пробку и тогда уже осторожно сделать окончательный прорез. Если его делать на весу, то, как правило, края прореза будут рваные. По окончании сверления вытолкнуть шомполом пробковый цилиндрик из сверла. Диаметр сверла при сверлении каучуковых пробок подбирать равным или немного большим вставляемой трубки. В качестве смазывающих веществ, облегчающих сверление резиновых пробок, обычно употребляют вазелиновое масло, глицерин или раствор щелочи. Для подгонки трубок к отверстию корковой пробки иногда делают расточку отверстия круглым напильником. Когда отверстие в пробке готово, смочить кончик отрезанной стеклянной трубки водой и вставить ее в пробку, все время держа трубку правой рукой на расстоянии 2–3 см от пробки и как бы ввинчивая ее. Держать пробку и стеклянную трубку так, чтобы руки были близко одна к другой. От частого употребления сверла тупятся. Тупое сверло рвет пробку, а не режет ее. Чтобы наточить сверло, применяют специальные ножи для точки сверл. Сверло надевают на коническую часть точила, нож прижимают к сверлу. При поворачивании ножа сверло, неподвижно зажатое в левой руке, точится довольно хорошо и быстро.

Сверление пробок: 1 — набор сверл для пробок; 2 — точка сверла; 3 — машинка для сверления пробок; 4 — сверление пробки; 5 — пробкомялка (жом) для пробок
ОПЫТЫ С МЕТАЛЛАМИ
• Никелирование металлических предметов Металлические изделия и предметы очищают от грязи, обезжиривают раствором соды, промывают в воде, на несколько секунд опускают в 50 %-ный раствор азот ной кислоты и повторно промывают дистиллированной водой. Подготовленное изделие выдерживают 30–50 минут в горячем растворе, содержащем на 1 л воды 280 г гептагидрата сульфата никеля и 100 мл концентрированной соляной кислоты. После получения никелевого покрытия (оно получается плотным и блестящим) изделие промывают водой и полируют суконкой.
• Кристаллы меди Все умеют выращивать кристаллы различных солей. А вот кристаллы меди не всякий сумеет вырастить. Для этого необычного опыта понадобиться: CuSO4, поваренная соль, кусок жести и стакан (можно обыкновенный). Из куска жести вы режьте круг, чтобы он свободно входил в стакан. В стакан насыпьте порошок сульфата меди(медного купороса) слоем в 5 мм и засыпьте этот слой солью.
ВНИМАНИЕ! Слои не перемешивать. Закройте слои кружком из фильтровальной бумаги и накройте кружком из жести. В стакан налейте раствор соли.
Через две недели вырастут довольно крупные кристаллы меди. Чтобы они хорошо сохранились поместите их в пробирку с раствором серной кислоты.
• Горение металлов Горение металлов в кислороде, хлоре широко известно. Менее знакомо горение металлов в парах серы. В штатив укрепляют вертикально большую пробирку, наполненную на одну треть серой, и нагревают до кипения серы. Затем в пробирку опускают пучок тонкой медной проволоки (можно предварительно нагреть) и наблюдают бурную реакцию.
ОБМЕННЫЕ РЕАКЦИИ
• Золотой дождь

Существует множество веществ с сильной температурной зависимостью растворимости. Именно на этом явлении и основан данный опыт. Для проведения этого опыта необходимо взвесить равные количества ацетата свинца(II) и йодида калия. Далее готовят два раствора. В два химических стакана наливают по 50 мл дистиллированной воды. В один добавляют 1 мл столового уксуса (или ~0,2 мл концентрированной уксусной кислоты) и растворяют ацетат свинца. Кислота добавляется для того, чтобы подавить гидролиз ионов РЬ2+. Во втором растворяют Kl. Затем оба раствора сливают в колбу из огнеупорного стекла объёмом 150 мл. При этом происходит реакция двойного обмена между ацетатом свинца и йодидом калия: РЬ(СН3СОО)2 + 2KI => 2КСН3СОО + РbI2 После смешивания растворов выпадает жёлтый осадок йодида свинца(II). Смеси растворов в колбе необходимо дать отстояться, чтобы осадок осел полностью. После этого с осадка осторожно сливают жидкость и вместо неё доливают 100 мл дистиллированной воды. Теперь раствор необходимо нагреть до кипения и кипятить в течении 2–3 мин. Осадок должен раствориться полностью. Если всё было сделано правильно, то после охлаждения раствора выпадет множество золотистых кристалликов, которые при встряхивании колбы будут парить в толще воды. Размер кристалликов очень сильно зависит от скорости охлаждения: чем медленнее охлаждать, тем более крупными и красивыми будут кристаллики. Для большей их прочности перед кипячением в раствор добавляют немного глицерина (~0,5 мл на 100 мл раствора).
• Затвердевание жидкости В химический стакан наливают 20–50 мл силикатного клея (силикат натрия) и добавляют 2–3 мл раствора соляной кислоты (1:1) и тут же перемешивают стеклянной палочкой. Через 30–40 секунд жидкость загустеет и ее уже нельзя вылить из стакана.
• Неорганический сад Другой, но не менее интересный опыт с силикатом натрия. Готовят горячий (около 60 °C) насыщенный раствор силиката натрия в 100 г воды. Полученный раствор медленно охлаждают до комнатной температуры, при этом образуется вязкая масса, напоминающая по консистенции силикатный клей. Затем берут несколько кристаллов различных окрашенных солей, например, меди, кобальта и никеля. Кристаллы осторожно помещают на дно стакана и оставляют на ночь. Утром можно полюбоваться на "неорганический сад", который вырастет из "семян" — кристаллов.

В методическом отношении, опыт интересен тем, что показывает один из вариантов (хотя и не самый распространенный) образования в природе силикатов, показывает явление осмоса (хотя в школе его не изучают) и еще раз доказывает, что химия не скучная наука. Уравнение реакции в случае с медным купоросом: CuSO4 + Na2SiO3 = CuSiO3 + Na2SO4
• "Заживление раны" Этот опыт демонстрируется в различных вариантах. Перед вами один из них. Кожу руки обильно смачивают "йодом" (слабый раствор хлорного железа), якобы для дезинфекции. После этого набирают в тонкую трубочку раствор роданида калия и проводят концом этой трубочки по смоченному месту (для усиления эффекта опыта можно использовать тупой нож). На руке появляется "кровоточащая рана", которую легко смывают затем обычной водой, а руку вытирают насухо полотенцем.
Примечание: Реакцию образования раствора роданида железа (III) можно использовать для получения хорошей имитации крови.
ОПЫТЫ ПО ОРГАНИЧЕСКОЙ ХИМИИ
• Холодное пламя Ингредиенты:
— сухая борная кислота, чайная ложка — этиловый спирт, чайная ложка — концентрированная серная или соляная кислота, одна капляВсе это помещается в блюдце, перемешивается и слегка подогревается на теплой (чтобы можно было держать руку) водяной бане. Выделяющийся эфир борной кислоты можно поджечь (не поднося спичку близко к блюдцу, чтобы не зажечь спирт). Эфир борной кислоты горит очень объемным, слегка зеленоватым пламенем, которое не только не обжигает, но даже не греет ощутимо, в нем спокойно можно держать руку, однако после того, как эфир борной кислоты весь выгорит, может начать гореть спирт, а его пламя действительно обжигает.
• Выделение кофеина из чая Эксперимент основан на способности кофеина, подобно иоду, подвергаться возгонке (сублимации — свойство вещества при нагревании переходить в газ минуя жидкое состояние). Для проведения опыта необходимы две столовые ложки листового чая. Чай перетирают в ступке до мелкого порошка, затем переносят в сухую фарфоровую выпаривательную чашку, которая сверху накрывается большой стеклянной воронкой, причем края воронки должны выдаваться за края чашки примерно на 1 см. После этого нагревают чашку над пламенем горелки. Кофеин сублимируется и вновь конденсируется на воронке в виде белых кристаллов.
• Получение фенолформальдегидной смолы В пробирку помещают 10 капель жидкого фенола и 8 капель 40 % формальдегида. Смесь нагревают на водяной бане до растворения фенола. Через 3 минуты в пробирку добавляют 5 капель концентрированной соляной кислоты и помещают ее в стакан с холодной водой. После образования в сосуде двух четких фаз следует слить воду и вылить полимер из пробирки. В течение нескольких минут образовавшаяся новолачная смола затвердевает.
• Получение искусственного шелка Около 20 г гидрокарбоната меди растворяют в 25 %-ном растворе аммиака путем его постепенного добавления к соли и при постоянном помешивании. В полученный медно-аммиачный раствор вносят медицинскую вату небольшими порциями и перемешивают до полного растворения клетчатки и получения вязкой массы. Растворение ваты идет медленно, поэтому опыт можно отложить на 1–3 дня. Для получения нитей искусственного волокна медно-аммиачный раствор клетчатки набирают в медицинский шприц и выдавливают через иглу, погруженную в осадительный раствор, представляющий собой 10–15 %-ный раствор серной или соляной кислоты.
УГЛЕВОДОРОДЫ
• Получение метана Реактивы: Ацетат натрия (безводный!), гидроксид натрия. Оборудование: Круглодонная колба, пробирка, лабораторный штатив (2), газоотводная трубка и пробка. Методика: В сухую пробирку или круглодонную колбу вносят тщательно прокалённый ацетат натрия CH3COONa и гидроксид натрия NaOH в соотношении 1:3 (по объему). Затем в пробирку вставляют пробку с газоотводной трубкой и, как обычно, проверяют прибор на герметичность. После чего, начинают нагревать смесь, сначала умеренно, затем более сильно. Образующийся метан СН4 собирают в пробирку методом вытеснения воды или воздуха, а затем поджигают. Газ сгорает подобно водороду — спокойно или со свистом — это зависит от чистоты полученного метана СН4. Реакция: CH3COONa + NaOH = Na2CO3 + СН4СН4 + 2O2 = 2Н2O + СO2
• Получение этилена и его свойства Реактивы: Этанол (96 %), серная кислота концентрированная, перманганат калия, бромная вода. Оборудование: Колба Вюрца (пробирка), спиртовка, прибор для собирания газов, ванночки с растворами (стаканчики). Методика: В пробирку или, что лучше в колбу Вюрца наливают 5 мл этанола и 2 мл серной кислоты. Смесь нагревают до 180°, при этой температуре начинает выделяться этилен (при более низкой температуре образуется эфир). Сначала этилен собирают в пробирку методом вытеснения воды и поджигают (газ легко сгорает). Затем газоотводную трубку прибора последовательно опускают в стаканчики или ванночки с раствором перманганата калия и с бромной водой. Наблюдают исчезновение (или значительное ослабление окраски). Реакция: С2Н5ОН +(H2SO4)= С2Н4 + Н2O
• Получение ацетилена Реактивы: Карбид кальция, перманганат калия, вода. Оборудование: Коническая колба, пробка с трубкой Методика: В коническую колбу объемом 100 мл кладут кусочек карбида кальция, наливают воду и затыкают пробкой с короткой трубкой. Сначала газ пропускают в подкисленный раствор перманганата калия, затем резиновую часть трубки снимают и поджигают ацетилен. Газ горит коптящим пламенем. Реакция: СаС2 + 2Н2O = Са(ОН)2 + С2Н2
• Свойства бензола Реактивы: Бензол, перманганат калия, бром, азотная кислота (65 %), серная кислота конц., железные опилки. Оборудование: Пробирки, химические стаканчики, делительная воронка Методика: 1. Запах бензола. Учащимся дают понюхать в пробирке бензол, и объясняют, что название "ароматические углеводороды" на прямую не связано с запахом бензола. 2. Отношение к воде и перманганату калия. В химический стакан наливают раствор перманганата калия (не слишком концентрированный), а затем бензол. Смесь расслаивается, причем граница раздела двух фаз становится зеркальной, но перманганат не обесцвечивается. 3. Горение бензола (под тягой!). В фарфоровую чашку наливают 1 мл бензола и поджигают горящей лучинкой. Бензол сгорает ярким коптящим пламенем. Примечание: категорически советую проводить опыт под тягой — нашей школе пришлось отмывать потолок от копоти! 4. Бромирование бензола. В колбу с длинной вертикальной трубкой ("обратный холодильник") наливают 4 мл бензол и 2 мл чистого брома. Реакция не протекает. Затем в смесь на кончике ножа добавляют железных опилок и закрывают пробкой с холодильником. Начинается довольно бурная реакция, по окончании которой на дно оседает тяжелый слой бромбензола. 5. Нитрование бензола. В колбу с обратным холодильником наливают 8 мл концентрированной серной кислоты и 5 мл азотной (65 %). Полученную нитрующую смесь охлаждаю под сильной струёй воды. Затем добавляют 5 мл бензола. Смесь встряхивают. Если реакция пойдет медленно. Всплывающий желтый слой и есть нитробензол. Его можно отделить на делительной воронке и использовать для дальнейших опытов (получение анилина и др.).
Внимание! Нитробензол ядовит! (ТЯГА!)Реакции: 2С6Н6 + 15О2 = 12СО2 + 6Н2О С6Н6 + Вr2 = С6Н5Вr + НВr С6Н6 + HNO3 = C6H5NO2 + Н2O
• Определение углерода и водорода в парафине Реактивы: Парафин, оксид меди (2), сульфат меди (безводный), известковая вода. Оборудование: Пробирки (2), штатив (штатив-держатель), пробка с газоотводной трубкой, спиртовка. Методика: В пробирку помещают оксид меди и парафин в объемном отношении 1:5. На её стенку аккуратно кладут обезвоженный сульфат меди, после чего, закрывают прибор пробкой с газоотводной трубкой и проверяют на герметичность. Смесь начинают осторожно нагревать. Когда парафин расплавится, трубку прибора опускают в известковую воду. Нагревание продолжают 5-10 минут, при этом: известковая вода мутнеет, сульфат меди синеет, на дне пробирки образуются зернышки металлической меди. Реакция: CnH2n+2 + (3n + 1)CuO = nСO2 + H2n+1O
СПИРТЫ
• Горение этанола Реактивы: серная кислота конц., перманганат калия, спирт. Оборудование: Спиртовка, стеклянные пластинки, стеклянные палочки Методика: Горение спирта в принципе, не нуждается в демонстрации, однако если этот опыт поставить эффектно, то можно произвести благоприятное впечатление на учащихся, а заодно повторить курс неорганической химии. На стеклянную или фарфоровую пластинку наносят немного кристалликов перманганата калия и прикасаются к ним стеклянной палочкой, смоченной в серной кислоте. При этом образуется сильный, и к тому же взрывоопасный (!) окислитель — оксид марганца (VII). Далее, стеклянную палочку подносят к фитилю спиртовки (фитиль должен быть мокрым), при этом спирт загорается. Реакции: 2КМnO4 + H2SO4 = Мn2O7 + Н2O + K2SO4 2С2Н5ОН + 2O2 = 4СO2 + 6Н2O + Q
• Качественная реакция на многоатомные спирты Реактивы: Глицерин (при отсутствии можно заменить сахорозой), раствор сульфата меди, гидроксида натрия. Оборудование: Демонстрационные цилиндры Методика: В демонстрационную пробирку или цилиндр наливают немного глицерина (обратить внимание учащихся на вязкость глицерина — причины: обилие водородных связей, "маслообразность" — влияние углеводородной цепи) и разбавляют водой. В другом цилиндре готовят осадок гидроксида меди, смешивая растворы гидроксида натрия и сульфата меди. Содержимое цилиндров сливают вместе и перемешивают. Наблюдают образование соединения с красивой, "васильковой" окраской. При подготовке учащихся к практическим работам опыт показывают в другой последовательности. В пробирку наливают раствор глицерина, добавляют гидроксид натрия (должен быть избыток), а затем сульфат меди. В этом случае, также наблюдается "васильковая" окраска, однако образование гидроксида меди не происходит, проведение опыта сильно упрощается, что важно для проведения практических работ. Реакция: СН2СНСН2(ОН)3 + Сu(ОН)2 = СН2СНСН2(OН)O2Сu (уравнение дано в упрощённом виде)
• Химические и физические свойства фенола Реактивы: Фенол, лакмус или метилоранж, гидроксид натрия, соляная кислота, бромная вода, азотная кислота, хлорид железа (III) Оборудование: Химические стаканы, демонстрационные пробирки, пробиркодержатель, спиртовка Методика: 1. К воде, налитой в химический стакан, прибавляют с помощью шпателя фенол и перемешивают смесь. Констатируют образование эмульсии, которая вскоре расслаивается. Объясняют, что нижний слой — раствор воды в феноле, верхний — раствор фенола в воде. 2. К части эмульсии полученной в предыдущем опыте прибавляют какой-нибудь индикатор. Среда — кислая. Значит фенол — кислота. Чтобы это подтвердить, к эмульсии прибавляют концентрированный раствор гидроксида натрия. Раствор становится прозрачным. На доске пишут соответствующую реакцию. Теперь, к полученному раствору фенолята натрия приливают раствор соляной кислоты. Снова образуется эмульсия, пишут уравнение, делают вывод о том, что фенол — кислота слабая. 3. Эмульсию фенола смешивают в демонстрационной пробирке с бромной водой. Выпадает белый осадок трибромфенола. Пишут реакцию, рассматривают взаимное влияние фенил-радикала и ОН-группы друг на друга. 4. Нитрование фенола. Опыт также иллюстрирует о, п-замещение в феноле. В пробирку насыпают немного фенола и расплавляют его (при этом демонстрируется его легкоплавкость — т. пл. 40 °C). После плавления смесь выливают в химический стакан с раствором азотной кислоты (2:3). При этом образуются различные продукты, в частности — пикриновая кислота (2,4,6-тринитрофенол — пикриновая кислота). 5. Качественная реакция на фенолы. Все фенолы с хлоридом железа (III) дают фиолетовое окрашивание.
• Качественное определение спиртов и фенолов Реактивы: Парафин, оксид меди (2), сульфат меди (безводный), известковая вода. Оборудование: Пробирки, штатив (штатив-держатель), пробка с газоотводной трубкой, спиртовка Методика: Задание: В трех предложенных вам пробирках находятся следующие вещества: а) фенол б) этанол в) пропантриол-1,2,3 Используя предложенные реактивы, определите содержимое каждой пробирки Реактивы: Растворы гидроксида натрия, сульфата меди (II), хлорида железа (III)
АЛЬДЕГИДЫ
• Получение альдегида Реактивы: спирт этиловый, медная проволока. Оборудование: спиртовка, химический стакан Методика: Из медной проволоки делают небольшую спиральку, которую раскаляют в пламени спиртовки и опускают в спирт. Эту операцию повторяют несколько раз. Полученный раствор обладает всеми свойствами альдегидов. Реакция: СН3СН2ОН + CuO = СН3СНО + Н2O
• Реакция полимеризации формальдегида Реактивы: формальдегид Оборудование: демонстрационные пробирки, пробка с газоотводной трубкой, спиртовка Методика: В демонстрационную пробирку наливают 10 мл 40 %-ного раствора формальдегида. Пробирку начинают нагревать в пламени спиртовки. Раствор начинает мутнеть (образуется параформ — полимер формальдегида). В другую пробирку насыпают немного сухого параформа, закрывают пробирку пробкой с газоотводной трубкой и начинают нагревать. В этом случае происходит мономеризация (деполимеризация) и образуется газообразный формальдегид (порошок улетучивается). Реакция: nСН2O = (СН2O)n
• Получение фенолформальдегидной смолы Реактивы: Фенол, формалин, соляная кислота (1:2). Оборудование: пробирки, спиртовка, фильтровальная бумага Методика: В пробирку помещают смесь 2,5 г фенола и 5 мл формалина. Ничего не происходит. Затем добавляют 7-10 капель соляной кислоты (катализатор). При этом начинается катализатор. Смесь охлаждают и дают ей отстояться. При этом в зависимости от условий образуется либо желтая смола, либо кусочек уже твердой белой смолы.
• Реакция серебряного зеркала Реактивы: Нитрат серебра (раствор 2 %), аммиак (раствор 25 %, разбавить в 8 раз), формалин (или другой альдегид), бихромат калия, серная кислота концентрированная (или готовая хромовая смесь). Оборудование: ершик, пробирки, химический стакан, спиртовка, пробиркодержатель Методика: Для проведения реакции берут пробирку или колбу, два раза моют ее ершиком, промывают хромовой смесью и ополаскивают дистиллированной водой. Эта стадия подготовки очень важна, т. к. от чистоты сосуда зависит исход опыты. В сосуд наливают на 1/4 раствор нитрата серебра и по каплям начинают добавлять раствор аммиака до полного растворения образовавшегося в начале осадка. К полученному раствору добавляют раствор альдегида. Полученную смесь аккуратно нагревают до потемнения растворы. Затем раствор выливают и показывают учащимся серебряное зеркало. Если зеркало не получилось, то не стоит браковать опыт: черный осадок это тоже серебро, только в виде мелкого порошка.
ПРИМЕЧАНИЕ: Аммиачную смесь не рекомендуется готовить заранее, т. к. при стоянии раствора в нем могут образоваться взрывчатые вещества — соли гремучей кислоты.Реакция: НСНО + Ag2O = НСООН + 2Аg
• Реакция с гидроксидом меди Реактивы: гидроксид натрия, сульфат меди, раствор альдегида. Оборудование: пробирки, химический стакан, спиртовка, пробиркодержатель Методика: В демонстрационную пробирку наливают раствор сульфата меди и добавляют к нему по каплям раствор гидроксида меди. Констатируют образование синего осадка гидроксида меди. Затем с осадка сливают часть жидкости и приливают к полученному осадку раствор альдегида. Смесь нагревают в пламени спиртовки, при этом происходит постепенное изменение окраски по схеме: синее-черное-зеленое-красное-желтое. Реакция: CuSО4 + 2NaOH = Cu(OH)2 + Na2SО4 НСНО + CuO = Cu2О + HCOOH
ЭЛЕКТРОНИКА
Автоматическое бесконтактное переключающее устройство[100]
В. НовожиловАвтоматическое бесконтактное переключающее устройство (АБПУ) предназначено для последовательной коммутации до десяти электрических цепей. Оно может оказаться хорошим помощником в световой организации демонстрационных и выставочных залов, клубов, дискотек, а также может быть с успехом использовано радиолюбителями для коммутации различных цепей. Частоту переключения цепей в АБПУ можно изменять в широких пределах. Предусмотрена также возможность в процессе переключения зафиксировать любой канал на длительное время, продолжить коммутацию либо вернуться в начальное положение с любого положения и с этого положения продолжать коммутацию в прежней последовательности. Номер каждого включенного канала высвечивается светодиодным индикатором, что позволяет непосредственно наблюдать за частотой переключения каналов. Отличительной особенностью устройства является отсутствие каких бы то ни было механических контактных групп, склонных к обгоранию при коммутации цепей с повышенным током нагрузки, что обеспечивает повышенную надежность, а также отсутствие в процессе коммутации радиопомех. Питание АБПУ производится от встроенного стабилизированного выпрямителя с выходным напряжением 5 В±10 %. Потребляемый устройством ток от источника питания при токовой нагрузке на каждый канал до 3 А не превышает 100 мА. Поэтому его можно питать также и от набора сухих гальванических элементов, имеющихся в широкой продаже. Принципиальная схема АБПУ приведена на рис. 1.
Рис. 1. Принципиальная схема АБПУ
Устройство собрано на пяти микросхемах серии К155 и одной — К514. На элементах DD1.1-DD1.3 выполнен генератор прямоугольных импульсов. С помощью переменного резистора R1 можно вручную изменять частоту генерации. Импульсы с выхода генератора поступают на вход делителя частоты, который состоит из двух последовательно включенных делителей на 2 (D-триггеры микросхемы DD2) и делителя на 10 (DD3). Таким образом, коэффициент деления равен 40. Импульсы с частотой повторения в 40 раз меньшей частоты задающего генератора поступают далее на двоично-десятичный 4-разрядный счетчик DD4, работающий в коде 1–2 — 4–8, выходы которого подключены к дешифраторам DD5 и DD6. Дешифратор DD5 управляет контрольным семисегментным светодиодным индикатором HG1, а к выходам дешифратора DD6 подключены блоки десяти транзисторных ключей. В процессе счета импульсов транзисторные ключи поочередно открываются, управляя включением тех или иных цепей. При подаче уровня логического 0 на вход S элемента DD2.1 нажатием кнопки с фиксацией SB2 счет прекращается, и счетчик хранит ранее полученную информацию. Таким образом, на неограниченное время остается включенным тот транзисторный ключ, который был включен в момент нажатия кнопки. При повторном нажатии кнопки SB2 на вход S элемента DD2.1 поступает уровень логической 1, счет импульсов возобновляется и происходит дальнейшее переключение транзисторных ключей. При нажатии на кнопку без фиксации SB1 делители DD2, DD3 и счетчик DD4 устанавливаются в нулевое состояние и после отпускания этой кнопки, если кнопка SB2 разомкнута, счет начинается с 1. Блоки транзисторных ключей могут быть реализованы по самым различным схемам в зависимости от характера коммутируемой нагрузки — номинальных значений напряжения и тока. На рис. 2 показаны некоторые возможные варианты схемного выполнения транзисторных ключей.
Рис. 2. Схемы транзисторного ключа: а — первый вариант ключа; б — схема мощного ключа; в — схема ключа для коммутации головных телефонов
Транзисторный ключ, схема которого показана на рис. 2,а, рассчитан на ток нагрузки до 1 А, однако в том случае, если выходной транзистор будет снабжен теплоотводом, ток нагрузки может быть увеличен до 3 А. Если вместо транзистора КТ315А использовать КТ315И, номинальное напряжение нагрузки и напряжение питания ключа может быть увеличено до 60 В. Допустим, что вход этого ключа присоединен к выходу 3 дешифратора D6. Когда на входы дешифратора будет подан код, соответствующий десятичному числу 2, на выходе 3 дешифратора окажется низкий потенциал, соответствующий уровню логического 0. Транзистор VT1.1 окажется заперт, а транзистор VT2.1 открыт до насыщения, и нагрузка окажется под током. При смене кода на входе дешифратора на его выходе 3 появится высокий потенциал, соответствующий уровню логической 1. Транзистор VT1.1 откроется, а транзистор VT2.1 закроется. Нагрузка будет выключена. Сопротивление резистора R2.1 подбирается таким, чтобы насытить выходной транзистор в открытом его состоянии. В качестве нагрузки на схеме показана лампа накаливания, но можно использовать и другие потребители. На рис 2,б показана схема мощного ключа, выходной каскад которого собран на транзисторе П210А. Для использования выходного р-n-р транзистора здесь понадобилось добавить к предыдущей схеме один инвертирующий каскад. При указанных на схеме сопротивлениях резисторов ключ обеспечивает в нагрузке ток 0,6 А. При уменьшении сопротивления резистора R2.2 увеличивается ток базы транзистора VT2.2 при запертом транзисторе VT1.2, уменьшается сопротивление коллектор — эмиттер VT2.2, что приводит к увеличению тока базы выходного транзистора и соответствующему увеличению тока нагрузки. При сопротивлении резистора R2.2, равном 1 кОм, ток нагрузки достигает 1,8 А. Для дальнейшего увеличения тока нагрузки нужно уменьшать сопротивления всех трех резисторов с соответствующим увеличением их мощности рассеяния. На рис. 2,в показана схема одного из вариантов транзисторного ключа для коммутации головных телефонов. На транзисторе VT2.3 собран однокаскадный усилитель звуковой частоты. Транзистор VT1.3 используется в ключевом режиме. Вход 1 блока подключается к источнику звукового сигнала, а вход 2 — к одному из выходов дешифратора DD6. Когда на выходе дешифратора, к которому подключен блок, потенциал соответствует уровню логического 0, транзистор VT1.3 заперт, на работу схемы не влияет и входной сигнал усиливается усилителем. Когда же на вход 2 поступает потенциал, соответствующий уровню логической 1, транзистор VT1.3 отпирается. При этом выходной транзистор запирается и сигнал на выход не проходит. Если головные телефоны подключены к выходам нескольких блоков транзисторных ключей, входы 2 которых соединены с выходами дешифратора, можно будет поочередно прослушивать звуковые сигналы от разных источников, подключенных ко входам 1 ключей, выбирая тот сигнал, который необходим. Конструктивно АБПУ может быть выполнено либо в виде отдельного устройства, либо в виде составного модуля какого-либо аппарата, в зависимости от выбранного назначения. Расположение элементов схемы произвольное. Конденсаторы С2-С4 должны быть подключены непосредственно к выводам питания микросхем DD2, DD3 и DD4 для обеспечения помехоустойчивости. Индикатор, кнопки управления и переменный резистор регулировки скорости коммутации целесообразно вывести на переднюю панель. Блоки транзисторных ключей при необходимости могут быть выполнены на отдельных платах и располагаться непосредственно около нагрузок. При этом они соединяются с цифровой частью АБПУ жгутом проводов, длина которого во избежание сбоев не должна превышать 10 м. Если выходные транзисторы ключей необходимо установить на теплоотводах, необходимо либо изолировать корпус транзистора от теплоотвода слюдяной прокладкой (при этом тепло-отвод может быть электрически соединен с общей шиной), либо изолировать теплоотвод от общей шины (при этом транзистор может быть электрически соединен своим корпусом с теплоотводом). В последнем случае обеспечивается лучший отвод тепла. Налаживание АБПУ начинают с проверки правильности монтажа согласно принципиальной схеме, после чего подключают питание с соблюдением полярности. Блоки транзисторных ключей пока не подключают. Правильно собранная цифровая часть устройства обычно начинает работать сразу, необходимо лишь с помощью осциллографа убедиться в наличии на выходе 8элемента DD1 прямоугольных импульсов. В противном случае необходимо подобрать емкость конденсатора С1. Индикатор должен высвечивать периодически сменяющиеся цифры от 0 до 9, скорость переключения которых должна плавно регулироваться переменным резистором R1. При нажатии кнопки «Запись» счет должен прекратиться, а индикатор должен высветить какую-либо цифру. Допустим, что высвечивается цифра 8. Тогда с помощью осциллографа или вольтметра нужно проверить наличие уровня логического 0 на выходе 9 дешифратора DD6 и уровня логической 1 на остальных выходах этого дешифратора. При повторном нажатии на кнопку «Запись» вновь должна начаться поочередная смена цифр на индикаторе. Остановив счет на другой цифре, опять проверяют уровни на выходах дешифратора DD6. Так проверяются все 10 выходов этого дешифратора, причем уровень логического 0 должен быть только на том выходе, который соответствует цифре, высвеченной индикатором. При нажатии кнопки «Сброс» индикатор должен высвечивать цифру 0. После этого несколько раз выключают и вновь включают питание. Генератор должен надежно запускаться, в противном случае следует заново подобрать емкость конденсатора С1. После налаживания цифровой части АБПУ можно к выходам дешифратора подключить блоки транзисторных ключей с их нагрузками. При налаживании ключей в зависимости от напряжения и тока нагрузки может понадобиться подбор сопротивлений резисторов для получения насыщенного состояния выходного транзистора в открытом состоянии. Налаживание транзисторного ключа, схема которого показана на рис. 2,в, производится следующим образом. Не подавая на вход 1 сигнал, устанавливают кнопкой «Запись» на входе 2 ключа уровень логического 0 и подбором сопротивления резистора R2.3 добиваются получения напряжения на коллекторе транзистора VT2.3, равного +3 В. Затем подают на вход 1 сигнал от какого-либо источника и убеждаются в его неискаженности при помощи осциллографа, подключенного к головным телефонам. Уровень входного сигнала подбирается соответствующим регулятором уровня источника. Затем кнопкой «Запись» устанавливают на входе 2 ключа уровень логической 1. Звук в телефонах должен полностью исчезнуть. При этом может понадобиться подбор сопротивления резистора R1.3. На этом процесс налаживания заканчивается. Чертеж печатной платы АБПУ приведен на рис. 3, 4.
Рис. 3. Печатная плата АБПУ (I часть)
Рис. 4. Печатная плата АБПУ (II часть)
Конденсаторное реле сверхдлительных выдержек времени
А. АристовОдин из распространенных автоматов выдержки времени — конденсаторное реле, поскольку оно, как правило, просто по конструкции и легко налаживается. Но выдержки времени этих реле обычно не превышают нескольких минут. Для получения же больших выдержек приходится использовать конденсаторы значительной емкости, обладающие пониженными сопротивлениями утечки, что снижает стабильность и надежность работы реле. Используя предлагаемый принцип конструирования конденсаторных реле времени, нетрудно построить автомат с выдержкой времени до… нескольких месяцев, причем времязадающий, конденсатор потребуется емкостью всего в несколько микрофарад. Принцип работы подобного реле поясняет рис. 1.
Рис. 1. Функциональная схема конденсаторного реле
Времязадающий конденсатор С1 после заряда его через выключатель S1 до напряжения источника питания подключается к разрядному резистору R1 и электронному ключу через выключатель S2, контакты которого замыкаются на непродолжительное время через определенные интервалы вспомогательным автоматом. Иначе говоря, конденсатор разряжается короткими порциями, что равносильно увеличению емкости конденсатора, а значит, выдержки реле времени во столько раз, во сколько продолжительность между замыканиями контактов выключателя S2 больше продолжительности их замкнутого состояния (можно сказать, что это значение соответствует скважности импульсов, управляющих контактами выключателя). Когда конденсатор разрядится до определенного напряжения, сработает пороговое устройство — электронный ключ — и реле К1 включит (или выключит) нагрузку. А теперь познакомимся со схемой реле времени (рис. 2), построенного на основе рассмотренного принципа. Времязадающий конденсатор здесь — С1. Он подключен через контакты К2.2 герконового реле К2 к пороговому устройству на однопереходном транзисторе V1. Контакты замыкаются периодически через определенные интервалы времени, задаваемые генератором, который управляет работой реле К2.
Рис. 2. Принципиальная схема конденсаторного реле.
Предположим, что конденсатор С1 уже заряжен до напряжения источника питания и отсчет выдержки времени начался. В те моменты, когда контакты замкнуты, конденсатор С1 разряжается через резистор R2 (параллельно ему можно подключить выключателем S1 резистор R3 и ускорить разряд конденсатора в 20 раз, уменьшив тем самым во столько же установленную выдержку времени). Реле подключено к генератору импульсов большой скважности, поэтому контакты замкнуты в течение сравнительно короткого времени, а в течение более длительного периода разомкнуты и конденсатор не разряжается. Управляющие работой реле импульсы дифференцируются цепочкой C3R7R4, в результате чего в момент окончания управляющего импульса (то есть во время заднего фронта его) в цепь базы 2 однопереходного транзистора поступает короткий отрицательный импульс амплитудой около 0,8 В, «разрешающий» открывание транзистора. Но напряжение на эмиттере транзистора пока велико, и он остается закрытым. Когда конденсатор разрядится до определенного напряжения и в этот момент на базу 2 поступит очередной отрицательный импульс, транзистор включится. Конденсатор С1 на короткое время (0,1…0,5 мс) окажется подключенным через открытый переход эмиттер-база 1 транзистора VI к резистору R5 и начнет заряжаться через него. В результате на резисторе появится положительный импульс, который через конденсатор С2 и диод V2 поступит на управляющий электрод тринистора V4 и откроет его. Сработает реле К.1 и контактами К.1.2 (на схеме не показаны) замкнет (или разомкнет) цепь нагрузки. Поскольку при этом откроется диод У5, реле К2 окажется подключенным параллельно реле KL. Через замкнувшиеся контакты К1.1, резистор R1 и контакты К2.1 конденсатор С1 зарядится до напряжения источника питания. Автомат готов к последующему отсчету выдержки. Диод V2 защищает управляющий р-n-переход тринистора от опасного для него обратного напряжения. Диоды V3 и V9 нужны для подавления экстратоков, возникающих при отпускании реле. Чтобы начать новый цикл отсчета времени, реле нужно установить в исходное состояние, когда закрыт тринистор, а реле К1 обесточено. Для этого достаточно замкнуть контакты выключателя S2. Конденсатор С4 быстро зарядится через резистор R9 до уровня напряжения на катоде тринистора, а конденсатор С5 будет более медленно заряжаться через резисторы R10, R11 до напряжения включения однопереходного транзистора V6. Через некоторое время этот транзистор включится и создаст положительный импульс напряжения на резисторе R9. Он окажется соединенным последовательно с напряжением на конденсаторе С4 и создаст на катоде тринистора более положительное напряжение по отношению к аноду. Тринистор закроется, реле К1 отпустит и контактами К1.2 переключит нагрузку. Контакты K1Л разомкнутся. Диод V5 закроется, и реле К2 вновь начнет периодически включаться и выключаться. Интервал времени между замыканием контактов выключателя S2 и отпусканием реле К1 можно установить переменным резистором R10 в пределах 0,15…5 с. Для увеличения этого интервала достаточно подключить параллельно конденсатору С5 дополнительный конденсатор соответствующей емкости. Такое, на первый взгляд, сложное устройство переключения реле времени в исходное состояние позволяет при необходимости автоматизировать работу устройства и добиться его периодического включения без постороннего вмешательства. После установки нужных выдержки и паузы реле будет отсчитывать заданные интервалы, включать (или выключать) нагрузку, а после некоторого перерыва вновь переходить в режим отсчета времени. Такой автомат сможет, например, подсыпать корм рыбам или мелким животным, поливать цветы во время отпуска, а совместно с кинокамерой — проводить покадровую съемку медленно протекающих процессов, таких как рост цветка. Генератор импульсов, управляющих реле К2, собран на кремниевых транзисторах V7 и V8, включенных как аналог однопереходного транзистора. Генератор вырабатывает импульсы прямоугольной формы и амплитудой, близкой к напряжению питания. Длительность импульсов около 20 мс, частоту их следования можно плавно изменять переменным резистором R12. В итоге выдержку времени удается плавно устанавливать в пределах 2,5…80 мин или 45 мин…26 ч (в зависимости от положения контактов выключателя S1). Кроме того, возможна установка фиксированных выдержек, если переключатель S3 устанавливать в другие положения. Так при подключении им к генератору резистора R14 выдержка может быть 3 мин или 1 ч (соответственно при замкнутых и разомкнутых контактах выключателя S1). Если же будет подключен резистор R13, выдержка соответственно станет 36 ч или 30 суток. В четвертом положении переключателя к генератору импульсов будет подключен разъем XI, в гнезда которого можно вставить выводы резистора, задающего любую другую выдержку реле. На однопереходном транзисторе V10 собран вспомогательный генератор, импульсы которого выполняют такую же роль для генератора на транзисторах V1, V8, что и импульсы последнего для каскада на транзисторе VI. Детали реле времени можно разместить в любом подходящем корпусе. При монтаже их следует помнить, что соединение выводов конденсатора С1 с контактом группы К2.1 должно быть выполнено только над платой («в воздухе»), чтобы обеспечить лучшую изоляцию этой точки и предотвратить дополнительный разряд конденсатора. Однопереходные транзисторы — любые из серии КТ117, биполярные можно заменить любыми аналогичными — кремниевыми транзисторами соответствующей структуры. Тринистор — любой из серии КУ101. Реле К1 — на напряжение 12 В, при токе срабатывания не более 50 мА. Реле К2 — самодельное. Обмотка его содержит 3000 витков провода ПЭВ-1 0,1, намотанных на каркасе из картона, изготовленного по размерам геркона, выполняющего роль контактов К2.1. Из герконов лучше применить тот, который рассчитан на работу при токах в доли микроампера. Для надежности работы реле внутрь каркаса катушки желательно поместить два геркона и соединить их выводы параллельно. Подойдет и готовое герконовое реле, срабатывающее при напряжении 4..8 В. Для питания реле использован блок БП-9/12, но вполне пригоден и любой другой источник напряжешь ем 12 В, рассчитанный на ток нагрузки не менее 80 мА. Если при расчете или подборе времязадающих резисторов R2, R3, R12-R14 окажется, что сопротивление любого из них не должно превышать 500 кОм, генератор на транзисторе V10 можно исключить. Налаживание реле сводится к подбору времязадающих резисторов и градуировке шкал переменных резисторов R10 и R12. В случае использования автомата для отсчета непродолжительных выдержек времени, например при проявлении фотопленки, его запускают кратковременным включением выключателя S2. Когда же автомат должен работать длительное время, например для подкармливания рыб в отпуске, контакты этого выключателя должны быть замкнуты постоянно.
Фотоэлектронные устройства (обзор)
В. ЛемкеФотоэлектронные (оптоэлектронные) устройства (ФЭУ) широко применяются в устройствах автоматики, телемеханики, оптической связи, фотоизмерительной и другой технике. Исходя из назначения, они должны соответствовать определенным требованиям, например таким: высокая чувствительность, достаточное быстродействие, способность работы в нужной части оптического диапазона и требуемом диапазоне температуры, помехоустойчивость, простота и экономичность устройства и т. д. Выполнение заданных требований достигается соответствующим выбором светочувствительных элементов и фильтров для них, схемы усилительных каскадов исполнительных устройств и т. д. В качестве светочувствительного элемента в ФЭУ могут быть использованы фоторезисторы, фотодиоды, фототранзисторы, фотоэлементы, фотоумножители, оптроны с открытым оптическим каналом и т. д. В зависимости от типа используемого светочувствительного элемента в ФЭУ есть свои особенности, которые должны учитываться и для более полного использования возможностей светочувствительного элемента и для более рационального построения устройства. Обычно в своих конструкциях радиолюбители используют фотодиоды и фоторезисторы. Фотодиод — полупроводниковый прибор, являющийся приемником лучистой энергии, которая, воздействуя на фотодиод, переводит его из закрытого состояния в открытое. В зависимости от типа фотодиода максимум спектральной чувствительности может находиться в видимой области или в области инфракрасного излучения. Фотодиоды могут работать и в вентильном режиме, преобразуя световую энергию в ЭДС значением 0.5…0.7 В. Фоторезисторы представляют собой активные полупроводниковые резисторы, чувствительные к световому излучению в широком интервале длин волн, включая ультрафиолетовый и инфракрасный спектры. Для фоторезисторов характерно то, что они обладают высоким сопротивлением в темноте, которое при освещении резко уменьшается. Существует большое разнообразие фЭУ, отличительную особенность которых определяет применяемый светочувствительный элемент. В сравнении по параметрам с фотодиодами фоторезисторы отличаются высоким отношением светового тока к темновому, однако, значительно уступают фотодиодам в быстродействии. Рабочее напряжение фотодиодов составляет 10…20 В, а фоторезисторов — 1,3…350 В. На практике ФЭУ выполняются, как правило, в следующих основных вариантах: — светочувствительный элемент без усилительных элементов в цепи управляющего устройства; — с использованием усилителя постоянного тока (УПТ); — с использованием порогового элемента; — светочувствительный элемент в цепи генератора импульсов; — с модуляцией управляющего луча. ФЭУ с фоторезистором или фотодиодом в цепи управляющего устройства без использования усилительных элементов отличаются простотой, но имеют низкую чувствительность и поэтому на практике применяются редко, например в устройствах «автостопа», где источник света находится в непосредственной близости от фоторезистора иди в автоматах включения и отключения ночного освещения, если в качестве исполнительного устройства применить тринисторы или электромагнитные реле с малым током срабатывания, а также для измерения интенсивности светового излучения при включении микроамперметра последовательно с фоторезистором. Световой ток фотодиода намного меньше светового тока фоторезистора и составляет десятки микроампер, поэтому нецелесообразно его ставить в цепи последовательно с исполнительными устройствами без усилительных элементов. В ФЭУ с УПТ желательно использовать фоторезисторы, так как при увеличении температуры окружающей среды на 30…40 °C темновой ток фотодиодов увеличивается в несколько раз, что вызывает нестабильность работы ФЭУ. Из применяемых в настоящее время ФЭУ с УПТ можно выделить три основные группы: 1. Фоторезистор включен между базой транзистора первого каскада УПТ и минусом (для транзистора р-n-р структуры) или плюсом (для транзистора n-р-n структуры) источника питания. 2. Фоторезистор включен между базой и эмиттером входного транзистора УПТ. 3. Фоторезистор включен в цепь УПТ, выполненного на интегральных микросхемах. Однокаскадный УПТ на транзисторе, а также УПТ на составном транзисторе, являющийся разновидностью однокаскадного УПТ с более высоким коэффициентом усиления, применяют в ФЭУ первой группы, например в автоматах управления освещением, в измерительных приборах. Для ФЭУ тревожной сигнализации, в устройствах для подсчета и сортировки штучных изделий и т. п. обычно применяются двухкаскадные УПТ. Повышения чувствительности ФЭУ первой группы можно добиться не только подбором по соответствующим параметрам транзисторов (указано ниже), но и путем увеличения напряжения источника питания. Лучше для питания ФЭУ использовать два источника: один с напряжением 5…20 В для питания УПТ, а другой, с более высоким напряжением, для питания фоторезистора. Подбирая напряжение второго источника в пределах 20…250 В, добиваются наибольшей чувствительности ФЭУ. Практические конструкции ФЭУ с УПТ первой группы приведены в «Радио», 1975, № 3, с. 37 и № 10, с. 64. ФЭУ, в которых фоторезистор включен между базой и эмиттером транзистора входного каскада УПТ, питаются от источника с меньшим напряжением, чем ФЭУ первой группы. В них не полностью используются возможности фоторезистора, поэтому ФЭУ даже с многокаскадными УПТ по чувствительности уступают ФЭУ с однокаскадными УПТ первой группы. Одно из ФЭУ с УПТ второй группы описано в «Радио», 1980, № 9, с. 38. Чувствительность ФЭУ с УПТ на транзисторах зависит от статического коэффициента передачи тока (h21э) транзисторов (особенно оконечного) и рабочего тока электромагнитного реле (или других исполнительных элементов). Чем больше коэффициент h21э транзистора и меньше ток реле, тем выше чувствительность ФЭУ. Транзистор, ко входу которого подключается фоторезистор, должен обладать как можно меньшим обратным током коллектора (IKBO < 2 мкА). Лучшие результаты дает применение кремниевых транзисторов. В радиолюбительской практике используются чаше всего двухкаскадные УПТ на транзисторах, так как построение многокаскадных УПТ связано с усложнением процесса установки режимов работы транзисторов при настройке ФЭУ. Поэтому для дальнейшего повышения чувствительности можно использовать УПТ, выполненные на интегральных микросхемах (ИМС) с высоким коэффициентом усиления. Однако при этом снижается помехозащищенность ФЭУ и использовать его можно при условии неизменности освещенности помещения, в котором установлен фоторезистор. ФЭУ выполняют на интегральных микросхемах не только для повышения их чувствительности, но и для уменьшения габаритов самих конструкций, повышения экономичности. Кроме того, в автоматических устройствах, выполненных на цифровых ИМС, легче согласовать выход ФЭУ со входом ИМС. Вариант ФЭУ, выполненного па одном элементе 2И-НЕ, описан в «Радио», 1977, № 3, с. 60. По чувствительности оно эквивалентно ФЭУ с однокаскадным УПТ первой группы и ее можно улучшить, если на фоторезистор отдельно подавать большее напряжение источника питания. Следует учесть, что в данном случае логический элемент выполняет функцию не порогового элемента, а УПТ, поэтому если освещенность светочувствительного элемента резко не изменяется, то и напряжение на выходе элемента может быть в пределах 0,4… 2,4 В. Если в ФЭУ на цифровых ИМС фоторезистор необходимо вынести на несколько метров, то для повышения помехоустойчивости и улучшения согласования с проводной линией связи следует применять логические элементы с открытым коллекторным выходом (например, элементы ИМС K155ЛA7, K155ЛA8) или использовать транзисторный согласующий каскад, который будет повышать и чувствительность ФЭУ. ФЭУ, в которых в качестве управляющих элементов используются электромагнитные реле, обладают большим гистерезисом, так как ток срабатывания реле больше тока отпускания. Поэтому включение и отключение исполнительных устройств происходит не при одинаковой освещенности фоторезистора. Кроме того, в таких ФЭУ при включении и выключении наблюдается нечеткое срабатывание, проявляющееся в виде «дребезга», особенно в случаях относительно медленного нарастания и убывания среднего уровня освещенности фоторезистора или когда уровень его освещенности имеет значительные колебания относительно среднего уровня. Чтобы избавиться от этих недостатков, нужно обеспечить не плавное, а резкое (скачкообразное) изменение силы тока, проходящего через обмотку реле, т. е. необходимо использовать для этого УПТ, обладающие пороговым эффектом (например, триггеры с эмиттерной связью). ФЭУ без пороговых элементов чаще всего используют для фотоизмерительных приборов и в тех устройствах, где необходимо плавное изменение значения регулирующего параметра, пропорционально изменению освещенности светочувствительного элемента. ФЭУ с использованием порогового элемента получили более широкое распространение. Иногда для простоты в качестве порогового элемента используют стабилитрон, включая его между выходом УПТ и базой транзистора ключевого каскада, нагрузкой которого служит электромагнитное реле или другое управляющее устройство. При достижении на нагрузочном резисторе УПТ падения напряжения, превышающего напряжение пробоя стабилитрона, он открывается и подает напряжение для управления ключевым каскадом. Однако такие устройства имеют ограниченное применение, так как не для всякого ФЭУ можно подобрать стабилитрон с нужным напряжением пробоя, кроме того, следует учитывать и разброс параметров стабилитронов и невозможность плавного регулирования этого порога без усложнения устройства. Более широкое распространение в радиолюбительской практике получили ФЭУ с использованием в качестве порогового элемента триггера Шмитта, принцип работы которого подробно рассмотрен в статье В. Крылова и В. Лапшина «Триггер с эмиттерной связью», опубликованной в сборнике «В помощь радиолюбителю», вып. 52. Некоторые ФЭУ с использованием триггера Шмитта описаны в «Радио», 1975, № 6, с. 35 и 1977, № 12, с. 55. В последнее время в устройствах автоматики все чаще применяют ИМС. При использовании для ФЭУ в качестве порогового элемента триггера Шмитта на транзисторах очень трудно, даже усложняя конструкцию, получить в исходном состоянии выходное напряжение, близкое к нулю. Т. е. на выходе триггера всегда присутствует потенциал, превышающий по значению логический 0. Для автоматических устройств, выполняемых на ИМС, целесообразно применять ФЭУ с использованием в качестве порогового элемента триггер Шмитта в интегральном исполнении. На рис. 1 приведена схема ФЭУ, обладающего хорошей чувствительностью.
Рис. 1. Принципиальная схема ФЭУ с использованием триггера Шмитта в интегральном исполнении
Для согласования высокого входного сопротивления фоторезистора R1 со входом триггера DD1 и для исключения влияния соединительной линии от вынесенного фоторезистора применен каскад на транзисторе VT1, выполняющий функцию УПТ и улучшающий крутизну фронта сигнала с фоторезистора. При затемненном фоторезисторе R1 на базу транзистора VT1 подается положительное напряжение около 0,7 В. В это время транзистор открыт, напряжение на его коллекторе (не более 0,8 В) меньше порога срабатывания триггера, поэтому на его выходе присутствует потенциал логической 1. При освещении фоторезистора его сопротивление уменьшается, напряжение на базе транзистора уменьшается до минус 0,8…1,6 В, в результате он закрывается, напряжение на коллекторе возрастает до значения, превышающего порог срабатывания триггера и на его выходе появляется логический 0. Подбором резистора RJ при затемненном фоторезисторе устанавливают на входе триггера напряжение 0,8 В, а подстроечным резистором R3 добиваются максимальной чувствительности ФЭУ. Если нужно чтобы при освещенном фоторезисторе на выходе триггера DD1 была логическая 1, коллектор транзистора VT1 следует подключить к выводу «+5 В», его эмиттер — непосредственно к входам триггера, резисторы R5 и R6 исключить совсем, а сопротивления резисторов R4 и R7 уменьшить соответственно до 820 и 910 Ом. В этом случае с уменьшением уровня освещенности напряжение на базе транзистора увеличивается от нуля до плюс 2 В. Транзистор при этом открывается и напряжение на его эмиттере возрастает от 0,9 до 2 В, превышая порог срабатывания триггера, в результате на выходе триггера появляется потенциал логического 0. Порог срабатывания триггера устанавливают подбором резистора R7. Однако сопротивление этого резистора не должно быть меньше 620 Ом, иначе триггер может самовозбудиться. Напряжение источника питания, равное 22 В (17 + 5) для фоторезистора ФСК-2, можно считать оптимальным для получения максимальной чувствительности. Существенным недостатком триггеров с эмиттерной связью является наличие гистерезиса, т. е. напряжение отпускания всегда меньше напряжения срабатывания триггера. Величина гистерезиса триггера составляет обычно десятые доли вольта. Меры по уменьшению гистерезиса рассмотрены в статье «Триггеры с эмиттерной связью», помещенной в сборнике «В помощь радиолюбителю», вып. 52, но они, к сожалению, значительно усложняют устройства. В качестве порогового элемента в ФЭУ лучше всего, пожалуй, применять не триггер Шмитта, а электронное реле В. И. Турченкова (принцип работы описан в «Радио», 1974, № 1, с. 37), обладающее рядом преимуществ, в том числе и меньшим гистерезисом и получением возможности хорошего согласования со входами цифровых ИМС. Примером может служить ФЭУ, описанное в «Радио», 1983, № 11, с. 48. Но в таком варианте ФЭУ применены два источника питания и не полностью использованы возможности электронного реле В. И. Турченкова. Порог срабатывания и отпускания электронного реле В И. Турченкова зависит от входного тока (от внешнего источника), текущего через коллекторный переход первого транзистора, и номинала резистора в коллекторной цепи этого транзистора. Отсюда следует, что простое ФЭУ на базе электронного реле В. И. Турченкова можно создать без источника входного тока, заменив резистор в коллекторной цепи первого транзистора фоторезистором с большим темновым сопротивлением. На рис. 2 приведена схема практически проверенного и сравнительно простого варианта ФЭУ.
БЛАГОСЛОВЕНИЕ
РЕДАКТОРАМ
Поверьте мне: судьбою несть
Даны нам тяжкие вериги.
Скажите, каково прочесть
Весь этот вздор, все эти книги, —
И все зачем? — чтоб вам сказать,
Что их не надобно читать!..
…
Зато какое наслажденье,
Как отдыхает ум и грудь,
Коль попадется как-нибудь
Живое, свежее творенье!
Многое в нашем мире было сделано любителями, не профессионалами. За такую работу денег не платят. Сколько фунтов стерлингов получил сэр Исаак Ньютон за свой первый закон механики, сколько за второй и сколько за третий? Глупый вопрос, нисколько. Средства для жизни он добывал за счет должности смотрителя Королевского Монетного Двора. До начала двадцатого века почти вся наука была создана любителями, людьми творческими, увлекающимися. Даже сам Эйнштейн создавал теорию относительности работая в патентном ведомстве. Тогда как тысячи тысяч профессиональных научных сотрудников двадцатого века, получая зарплату, не создали ничего примечательного. Журнал «Домашняя лаборатория» тоже был создан любителями именно для любителей научно-технического творчества. Естественно, что журнал редакторам не платит, поскольку он не делает деньги и не стремиться к этому. В наше меркантильное время, когда деньги — мерило всего, редактором такого журнала может стать только энтузиаст. Журнал «Домашняя лаборатория» отличается от других изданий, прежде всего тем, что он создан и циркулирует в Интернет. В единой, но распределенной структуре. И его редакция также распределена. Не важно где расположены его редакторы. Это может быть и потомок русских эмигрантов проживающий в ЮАР, и русскоязычный канадец, и знающий русский болгарин, и потомственный москвич, и житель далекого Томска в Сибири. Это не важно. Важно то, что это все люди увлекающиеся, творческие, интересующиеся самыми разнообразными областями научных знаний. Редактор журнала оказывается в гуще научных веяний во всей их широте и разнообразии. Роскошная пища для ума! Новая информация — это расширение вашей эрудиции, осознание ее — ваше самосовершенствование. А если вы узкий специалист, вы можете уже в силу своей должности редактора наткнуться на гипотезу или эксперимент из смежной области, которые дадут новый поворот в решении вашей научной проблемы. Вы попадаете в богатейшую информационную среду — питательную среду для ученого человека, для его интеллекта. Не минуют вас и проблемы этические. Ведь вы выступаете в роли судьи, вы выносите приговор по делу, которое может быть для кого-то жизненно важным. Имеете ли вы право на такой образ действий? Достаточно ли ваших знаний для объективности вашего решения? Наконец, не захватил ли вас «синдром уборщицы», которая, будучи облечена атрибутами власти в виде швабры и тряпки, может наорать на идущего по коридору нобелевского лауреата? Такого рода этические проблемы за вас никто не решит. Решая их самостоятельно, вы очертите место своей личности в этом мире, а значит, достигните духовного усовершенствования. И, наконец, может быть, главное. Современное общество уже называют информационным. Когда-то, в каменном веке, человечество умело обращаться только с веществом, затем оно овладело энергией, и вот теперь вышло на высшую ступень овладения информацией. И редакторы журнала «Домашняя лаборатория» должны владеть, и владеют всеми современными информационно-компьютерными технологиями. Обработка научной информации, копание в навозной куче в поисках жемчужного зерна, отделение плевел от зерен — важнейший для человечества род деятельности. Вы стали редактором? Отсеивайте разумное, доброе, вечное! Успехов вам в этой благородной деятельности! В ДОБРЫЙ ПУТЬ! * * * НА ОБЛОЖКЕ
Тихо, тихо, улитка ползи По склону горы Фудзи.С такой обширной областью науки и практики, как биотехнология, следует знакомиться постепенно, не спеша. В этом номере смотрите о ней разделы "Ликбез" и "Лаборатория".
Последние комментарии
3 часов 23 минут назад
7 часов 37 минут назад
9 часов 56 минут назад
11 часов 45 минут назад
17 часов 31 минут назад
17 часов 36 минут назад