Эмбрионы в глубинах времени [Марсело Санчес] (fb2) читать онлайн
- Эмбрионы в глубинах времени (пер. Павел Иванович Волков) 2.53 Мб, 260с. скачать: (fb2) - (исправленную) читать: (полностью) - (постранично) - Марсело Санчес
[Настройки текста] [Cбросить фильтры]
[Оглавление]
Марсело Санчес ЭМБРИОНЫ В ГЛУБИНАХ ВРЕМЕНИ Каменная летопись биологии развития
Благодарности
Я хотел бы поблагодарить Цюрихский Университет и его Научный Факультет, а также Палеонтологический Институт за создание вдохновляющей, побуждающей к работе и благоприятной обстановки, в которой я смог написать эту книгу. Я благодарю нынешних и бывших сотрудников моей лаборатории и близких коллег за всё, что я узнал от них: Ингмара Вернебурга, Лауру Уилсон, Торстена Шейера, Кристиана Митгатча, Массимо Дельфино, Яси Хьюджи, Кристиана Колба, Даи Койябу, Джеймса Нинана, Фреди Карлини, Сандрин Ладевез, Коринн Уиммер, Яна Прочела, Питера Менке, Мадлен Гейгер, Катю Полачовски, Фиону Стрэл, Патрисию Мейер, Веру Вайсбекер, Фернандо Галлиари, Лайзу Рэйджер, Томаса Шмельцля и Анджали Госвами. Я также благодарю некоторых коллег из Цюриха и из-за границы за обсуждение идей и за любезно предоставленную информацию: Леннарта Олссона, Шигеру Куратани, Майка Ричардсона, Хироши Нагашиму, Хьюго Бюше, Тома Кемпа, Джона Спайса, Норберто Джанини, Йоханнеса Мюллера, Райнера Шоха, Шиге Кураку, Андреаса Вагнера, Инесс Хоровиц, Орэнжел Агилеру, Лайонела Готье, Рено Лебрана, Анджали Госвами, Норму МакЛеод, Рика Мэддена, Мартина Сандера, Кэтлин Смит, Пола Тейлора, Рафаэля Хименеса, Нико Гудмана, Клода Монне, Майкла Хаутманна, Кристиана Клюга, Хайнца Феррера, Ричарда Хоффманна, Кеннета де Бэтса, Томаса Мартина, Александра Нютцеля, Клаудию Хоффманн, Фреди Карлини, Карин Ниффелер и Северина Урди. Я благодарю всех, кто внёс свой вклад в издание «Seminars in Cell and Developmental Biology», которое я редактировал в 2010 году, за помощь, вдохновившую на написание этой книги: Ричарда Клотье, Зерину Джохансон, Мойю Смит и её коллег, Луизу Хэмфри, Кристофа Золликофера, Марсию Понс де Леон, Мартина Сандера, Николь Клейн, Надю Фробиш, Флориана Витцманна, Райнера Шоха, Дженнифер Олори, Торстена Шейера и Массимо Дельфино. Роб Ашер в Кембридже, Англия, Лаура Уилсон и Торстен Шейер в Цюрихе, а также Анна-Кристина Хоннен в Берлине любезно и критически перечитали ранние наброски и сделали очень полезные замечания. Три анонимных рецензента, а особенно Мишель Лорин, были очень щедры на критику и предложения, которые улучшили ясность этой работы и помо¬гли мне избежать ошибок. Торстен Шейер, Кристиан Митгатч, Джеймс Нинан, Лайза Рэйджер, Кевин де-Карли (Цюрих) и Найджел Хьюз (Риверсайд) любезно просмотрели части текста. Торстен Шейер и Яси Хьюджи дали весьма нужные советы, касающиеся главы о палеогистологии, а Кристиан Митгатч — по онтогенетическим и историческим вопросам. Чже-Си Луо поделился информацией, касающейся «ранних млекопитающих» со своей обычнойотзывчивостью и добрым юмором. Северин Урди (Цюрих) любезно представил развёрнутые комментарии и догадки, касающиеся букашек и ракушек. Любые несоответствия или огрехи, которые остались здесь — полностью на моей ответственности. За помощь с графикой и вопросами форматирования я благодарю Морану Михальевич, Кевина Де-Карли, Рози Ротт, Фиону Стрель и Торстена Шейера. Мадлен Гейгер, Катя Полачовски и Клаудия Джоел прекрасно выполнили некоторые из рисунков. Джон А. Лонг (Лос-Анджелес), Мэтт Фридман (Оксфорд), Ясмина Хьюджи (Цюрих), Йоахим Хауг (Нью-Хейвен), Панчо Гоин (Ла-Плата), Чже-Си Луо (Питтсбург), Торстен Шейер (Цюрих), Кристиан Клюг (Цюрих), Кристоф Золликофер и Марсия Понс де Леон (Цюрих), Луис Костер (Базель) и Райнер Шох (Штутгарт) очень великодушно дали право воспользоваться принадлежащими им изображениями. Я благодарю за очень нужную и прекрасно оказанную помощь Хейке Готцманна — за решение административных вопросов, и Хейни Уолтер — за помощь в вопросах, связанных с информационными технологиями. Я также благодарен директору моего института Хьюго Бюше за его поддержку. Линн Мейнхардт, Кейт Уорн и Чак Крамли из издательства Калифорнийского Университета быстро отвечали на мои многочисленные вопросы, возникавшие в процессе редактирования, и любезно дали мне много рекомендаций разного рода, а Шейла Берг сделала в отношении рукописи очень полезные и ёмкие замечания, касающиеся редактирования и стилистики. Вольфганг Майер в Тюбингене поддерживал мой интерес к онтогенезу на протяжении многих лет и был очень ценным наставником. Петер Хольцварт в Цюрихе, Роб Ашер в Кембридже, Энн-Кристин Хоннен в Киле и Берлине, и моя мать Глория Виллагра в Пуэрто-ла-Крусе/Буэнос-Айресе оказали мне моральную поддержку. В последние несколько лет я пользовался поддержкой главным образом Швейцарского Исследовательского Совета (SNF) и Цюрихского Университета.Пролог
Разнообразие жизни обычно представляется в виде эволюционных древ: ветвящееся изображение завершается рисунками животных и растений. Это хорошо, поскольку очертания древа отображают общую историю, которую разделяют друг с другом живые организмы, в том числе и мы сами. Но у этого способа отображения информации есть одно ограничение. Изображённые живые организмы — это статичные объекты, обычно во взрослом состоянии с узнаваемыми особенностями своего вида. В реальной действительности организмы изменяются на протяжении своей жизни. Если мы захотим изобразить биологию биоразнообразия, мы могли бы показать для каждого организма фильм, сделанный в технике замедленной съёмки и демонстрирующий различные стадии истории жизни — для многоклеточного организма от оплодотворения до смерти. Варианты и количество изменений, которые происходят на протяжении жизни индивида, настолько многочисленны и сложны, что каждой стадии этого процесса посвящены различные научные дисциплины. Первые шаги деления клеток и формирования эмбриона ассоциируются с эмбриологией. Многие исследования в этой области относятся, например, к гаструляции — стадии, на которой образуются зародышевые листки и устанавливается план строения тела взрослого организма. Специалисты по биологии развития склонны изучать момент возникновения или происхождение тканей и органов. Специалисты в области сравнительной анатомии и зоологи обычно интересуются изменениями, возникающими после рождения, которые некоторые биологи любят называть словом «рост», противопоставляя его развитию. Для всех этих дисциплин молекулярная биология предложила методы и концепции, позволяющие поставить совершенно новый набор вопросов, касающихся механистических основ эволюции истории жизни, а также заново ответить на старые вопросы, поставленные морфологами. Индивидуальное развитие — это благодарный предмет для исследований в биологии. А как вам тот факт, что большинство видов, представленных на древе жизни, уже вымерло? Некоторые могут подумать что палеонтология нужна биологии развития, как рыбе велосипед. Они также могут утверждать, что летопись окаменелостей полностью умалчивает о многих аспектах эволюции индивидуального развития и генетики. Я не думаю, что дела обстоят именно так, и я написал эту книгу, чтобы объяснить, почему. Конечно, палеонтология всё ещё в значительной степени умалчивает о таких ключевых вопросах, как гаструляция. Потому и важно определить границы, которых может достичь палеонтология. В этом контексте мне вспоминается вклад известного специалиста по анатомии рыб в спорах 1980-х годов: роль окаменелостей против молекул при реконструкции древа жизни. Колин Паттерсон (1933-98), важная фигура в исследованиях окаменелостей, десятилетиями работавший в палеонтологическом отделе Музея Естественной Истории в Лондоне, настоятельно выступал в пользу главенства информации о ныне живущих видах над ископаемыми остатками при исследовании эволюционных отношений. Большинство коллег-палеонтологов жёстко критиковало его. Несколько лет спустя его идеи стали оцениваться намного выше, и в 1996 он был предоставлен к медали Ромера-Симпсона — это высшая степень одобрения Общества Палеонтологии позвоночных. Роль палеонтологии в реконструкции древа жизни в значительной степени признана большинством биологов, несмотря на ограничения, возникающие при работе с фрагментарными данными, что резко контрастирует с большим массивом информации (например, геномной), который известен сейчас для большего и большего количества ныне живущих видов. Интеграция информации из областей палеонтологии и эмбриологии имеет давнюю историю. В викторианскую эпоху, например, Томас Гексли сделал важный вклад в обеих областях знания: например, он был первым, кто предположил, что птицы родственны динозаврам, и он же открыл главные аспекты ранних стадий жизни книдарий, группы, к которой принадлежат кораллы и медузы. Фактически, многие исследователи продолжают вносить большой вклад в эти очень различающиеся области знания. Например, Фил Донахью из Бристоля изучает конодонтов, базальную группу позвоночных животных, которая вымерла примерно 200 миллионов лет назад, но у него также есть исследовательская программа, касающаяся микро-РНК и её роли в морфологическом разнообразии. Взятые вместе, его исследования в области палеонтологии, биологии развития и молекулярной биологии ведут к лучшему пониманию истории жизни. В этой книге я исследую то, что мы можем узнать о развитии организмов непосредственно из летописи окаменелостей. Ископаемые остатки — это не просто статичные объекты, части более не существующих животных. Глядя проницательным взглядом, обладая соответствующей информацией и оставаясь в концептуальных рамках, мы можем многое узнать о размножении и развитии вымерших животных. Может показаться, что «моментальный снимок» какой-то стадии развития, который даёт окаменелость, окружён таким количеством неизвестностей, что интерпретация зачастую неполной анатомии остаётся, в лучшем случае, предположительной. Но существуют методы изучения неполных окаменелостей, и выводы, сделанные большинством палеонтологов, основаны на материальных свидетельствах. Эта книга предназначена для людей, обладающих общим знанием биологии и интересом к ископаемым остаткам и эволюции. Примечания и ссылки в конце книги могут помочь разъяснить и уточнить разнообразные вопросы, к которым я здесь обращаюсь. Я прошу, чтобы мне простили несколько случайный характер упоминаемых ссылок, поскольку некоторые из затронутых здесь тем очень обширны, и им сопутствует долгая история исследований и плодотворных размышлений. Многие примеры взяты из моей собственной работы и из работ моих самых близких коллег, поскольку эти примеры я знаю лучше всего. В конце концов, каждая книга представляет собой личный взгляд на проблему, и эта вовсе не является исключением.Глава первая Окаменелости, онтогенез и филогенез
Я помню, как на меня, тогда ещё ребёнка, произвело очень сильное впечатление утверждение, ошибочно приписанное Томасу Гексли, где говорится, что, если посадить обезьян перед пишущими машинками, а они будут случайно нажимать клавиши, то при наличии достаточного времени они наверняка напечатали бы полную «Британскую энциклопедию». У меня есть сокращённый вариант «Энциклопедии» на испанском — пятнадцать толстых томов; поэтому я представляю себе, о каком массиве текста идёт речь. Впервые я прочитал это утверждение в креационистском буклете, где было указано на абсурдность этого утверждения.[1] Но для меня оно имело смысл: маловероятно, но всё же, при наличии бесконечного времени, это могло бы случиться. При написании этой книги я решил немного исследовать этот вопрос, и выяснил, что этот мысленный эксперимент с обезьянами и пишущими машинками всерьёз рассматривается с философской и статистической точки зрения и используется в различных популярных работах. Фактически, это один из наиболее известных мысленных экспериментов, из датируемого 1913 годом эссе французского математика Эмиля Бореля. С тех пор он стал популярной иллюстрацией в теории вероятностей. Вполне очевидно, что вероятность того, что обезьяны напечатают «Британскую энциклопедию» или произведения Шекспира, бесконечно мала. Насколько это уместно при обсуждении эволюции и индивидуального развития?[2] Здесь есть две точки обсуждения: временной отрезок и вероятность эволюционного изменения, возникающего в течение этого времени. Геологическое время не бесконечно, но оно долгое, очень долгое, или глубокое — хорошее слово для описания, предполагающее, что в глубинах пород скрыто то, благодаря чему мы можем многое узнать об этом отдалённом прошлом. Понятие «глубокое время» возникло вместе с популярной работой по геологии американского автора Джона МакФи под названием «Basin and Range» В ней он изложил, как геологи интеллектуально и эмоционально развивают у себя ощущение необъятности времени.[3] Осознание наряду с этим необъятности биологического разнообразия ископаемых форм открывает исключительные перспективы, и это может быть наиболее фундаментальным вкладом в человеческое понимание вселенной в сравнении с тем, что могут дать геологи и палеонтологи. Благодаря датированным ископаемым остаткам, размещённым на эволюционных древах, и оценкам, основанным на молекулярных данных, мы знаем, что жизнь произошла, по крайней мере, от 3,5 до 3,2 миллиардов лет назад, и что возраст многоклеточной жизни составляет, как минимум, 700 миллионов лет.[4] В двадцатом веке одним из главных достижений в геохимии была разработка нескольких методов датирования горных пород, основанных на изотопах различных химических элементов, что привело к надёжному установлению абсолютного исчисления времени протяжённой истории Земли и жизни. Считалось, что эволюции биологического разнообразия требовались длительные периоды времени. Дарвину было важно собрать информацию о древности Земли и жизни. Он был озабочен тем, чтобы для эволюции действительно сложных структур, вроде глаза, было «достаточно» времени. Дальнейшее открытие мутаций и их «беспорядочности», на первый взгляд, сделало оправданным волнение Дарвина. Эволюция далека от случайного процесса, и обезьяны за пишущими машинками — это не лучшая аналогия для эволюционных процессов. Одно из главных замечаний, сделанных в недавно вышедших книгах по эволюции — указание на нехаотичность и предсказуемость эволюции. Это не теоретическое заключение, а скорее нечто, проявляющееся эмпирически в характере явлений, заметных в фенотипах и генотипах ныне живущих видов, а также в ископаемых остатках живых организмов. Факт того, что мутации в некоторых генах имеют высокую вероятность быть отобранными неоднократно в независимых родословных линиях, сталкивающихся со схожими условиями окружающей среды, получил название «параллельная генотипическая адаптация». Это делает генетические траектории адаптивной эволюции до некоторой степени предсказуемыми, что даёт ключ к реконструкции молекулярных процессов, которые с большой степенью вероятности работали у вымерших видов, несмотря на случайный характер эволюции. Индивидуальное развитие у вымерших видов также можно реконструировать благодаря открытию принципов, которые являются общими для огромных групп видов и даже включают использование одних и тех же генов, отвечающих за индивидуальное развитие. И всё же разнообразие, как в смысле количества видов, так и в смысле многообразия форм, настолько же обширно, насколько протяжённо время, в течение которого оно эволюционировало.Вымирание большей части жизни на Земле
Теория эволюции даёт рациональное объяснение богатому биологическому разнообразию, которое окружает нас. С каждым днём всё больше людей по всему миру живёт в городах, но даже те, кто редко сталкивается с природой, имеют представление, по крайней мере, о части этого разнообразия благодаря телевидению или посещению зоопарка. Уже описано приблизительно 1,5 миллиона видов, и, по разным оценкам, планету населяет от 10 до 100 миллионов видов. Но это огромное количество видов — лишь малая часть от общего количества видов, которые существовали на планете; консервативные оценки предполагают, что не менее 99 процентов от общего разнообразия жизни, которая существовала на Земле, вымерло.[5] Большая часть этого минувшего разнообразия в значительной степени не документирована, несмотря на работу палеонтологов, поскольку ископаемых остатков тех видов больше не существует, или же они ещё ждут своего открытия и изучения. Часть этого огромного разнообразия, которая знакома нам самым непосредственным образом — это позвоночные, группа животных, обладающих позвоночным столбом, к которой принадлежим и мы сами. Существует примерно 59000 ныне живущих видов позвоночных, но было описано ещё больше видов{1}. Некоторые группы позвоночных с палеонтологической точки зрения известны лучше, чем другие. Для динозавров, например, известно приблизительно 550 описанных родов, но по оценкам специалистов, должно было существовать примерно 1850 родов. Не все новые роды и виды ископаемых организмов, которые были описаны, являются валидными, потому что палеонтологи непрерывно проводят ревизии своих выводов, касающихся таксономии, и новые исследования в области видового разнообразия и анатомии помогают уточнить критерии, на которых основываются выводы. Обзор, сделанный в 2003 году, выявил, что валидные на тот момент 4399 родов ископаемых млекопитающих составляли 80 процентов от общего количества когда-либо описанных родов. Это число составляло 67 процентов в 1945 году, как сообщал в своей классической статье американский палеонтолог Джордж Гейлорд Симпсон. Свои названия получают всё больше и больше валидных ископаемых родов млекопитающих и других позвоночных. Случай с эволюцией нашего собственного рода Homo иллюстрирует противоречия и уклоняющиеся мнения, касающиеся таксономии ископаемых форм. Однако по многим вопросам имеет место согласие во мнениях. Например, большинство антропологов подтверждает, что около двух миллионов лет назад по меньшей мере пять различных видов людей населяли планету. Мы — последняя ветвь когда-то гораздо более ветвистого эволюционного древа, существующего с момента отделения наших предков от шимпанзе примерно семь миллионов лет назад. Цифры, которые я привожу здесь, служат просто для иллюстрации того факта, что, если мы хотим понять эволюцию биологического разнообразия, взгляд в прошлое является основополагающим. Роль палеонтологии в описании исчезнувшего биологического разнообразия очевидна, но, помимо простого описания, существует ещё много способов, которыми палеонтология вносит свой вклад в эволюционную биологию.Летопись окаменелостей и что она рассказала нам об индивидуальном развитии
Новые области для исследования, возникшие в ходе подъёма молекулярной биологии в последние десятилетия, вызвали целую волну вполне обоснованного энтузиазма. Это также привело к постановке в определённых кругах вопроса об оправданности дальнейшей работы в таких старых дисциплинах, как палеонтология. В контексте обсуждения этого вопроса многие из моих коллег выделили ряд специфичных аспектов, когда только обращение к палеонтологии способно помочь другим биологическим дисциплинам и создать целостную программу исследований. Они таковы: 1. Ископаемые остатки живых организмов образуют шкалу времени для эволюции. Древнейшие представители групп живых организмов из пород, датировка которых представляется возможной, предоставляют информацию о сроках, в течение которых, по крайней мере, могло произойти разделение между группами живых организмов. Если у нас имеются последовательности ДНК двух ныне живущих видов и благодаря этому знаем, насколько они различаются, и, если мы знаем, когда эти формы стали отличаться друг от друга в летописи окаменелостей, мы сможем определить скорость изменений в ДНК. Тогда эту скорость можно использовать для оценки времени дивергенции у других, родственных групп, для которых ископаемые остатки ещё не известны.
Рисунок 1. Главные геологические эры и периоды, упоминаемые в тексте, и их возраст в миллионах лет.
2. Ископаемые остатки предоставляют информацию об историческом распределении организмов в пространстве. Сам молодой Чарльз Дарвин обнаружил это во время своего кругосветного путешествия. Он описал ископаемых морских моллюсков из Анд, что явно указывало на драматические изменения, происходящие на протяжении геологического времени. Открытие ископаемых родственников утконоса в Южной Америке несколько лет назад, наряду с другими открытиями, даёт несомненное свидетельство в пользу ранее предполагавшихся фаунистических связей с Австралией. Распространение по всем южным континентам родственника ранних млекопитающих Lystrosaurus, существа размером с собаку, в триасе, а также ископаемых морских рептилий, называемых мезозаврами, в пермский период, даёт независимые свидетельства в пользу дрейфа континентов. Ископаемые остатки документально свидетельствуют о существовании в прошлом на островах многих видов, на которые повлияло присутствие человека, во многих случаях приведшее их к вымиранию в доисторические времена, как это случилось со многими гигантскими лемурами, найденными в пещерах на Мадагаскаре. Список примеров, действительно, очень длинный, и значение ископаемых остатков в решении (и создании) биогеографических загадок, а также их роль в проверке гипотез, основанных на ныне живущих видах, бесспорны. 3. Ископаемые остатки могут документировать порядок, в котором появился комплекс особенностей, в настоящее время являющийся диагностическим для современной группы организмов или для вида. Современные млекопитающие являются, в дополнение ко многим другим признакам, уникальными обладателями волос, молочных желёз и двух смен зубов. Однако, мы знаем, что эти черты появились не все сразу, что отражено в богатой летописи окаменелостей, уходящей в прошлое примерно на 315 миллионов лет — это минимальное время, обозначенное ископаемыми остатками для расхождения родословных линий млекопитающих и рептилий от их общего предка. 4. Ископаемые остатки, документирующие происхождение групп ныне живущих видов, критически важны для проверки родственных отношений между живыми организмами. Без вымерших динозавров-теропод, иллюстрирующих происхождение птиц, например, гораздо труднее понять, как курица может быть более близким родственником черепахе, чем млекопитающему. Окаменелости улучшают точность реконструкции эволюционного древа путём установления того факта, что признаки могут эволюционировать параллельно, как, например, постоянная температура тела у птиц и млекопитающих. Они также могут давать прямую информациюотносительно того, как выглядели первые перья и волосы этих животных на ранних этапах эволюции. Ископаемые остатки часто документируют организмы с уникальной мозаикой особенностей, которая является ключевой для нашего понимания происхождения и функции тех или иных признаков у ныне живущих организмов. Тероподы демонстрируют уникальную комбинацию особенностей, о которых мы бы не узнали, если бы смотрели только на ныне живущие виды. Кто бы мог предположить, что эти существа, жившие в меловой период, такие, как наземный Tyrannosaurus rex, обладали разноцветными перьями?[6] К летописи окаменелостей, пусть и такой неполной, как она есть, нужно относиться очень бережно как к наиболее важному источнику свидетельств того, что в действительности происходило на протяжении больших отрезков времени, охватывающих крупномасштабные эволюционные изменения. С помощью палеонтологических исследований могут быть разрешены фундаментальные вопросы, касающиеся эволюции. Например: • Протекает ли эволюция с относительно постоянной скоростью путём экспоненциального увеличения числа родословных линий, прерываемых только событиями вымираний? Или же она протекает путём взрывного видообразования, включая «адаптивную радиацию», в остальное время оставаясь относительно стабильным процессом? • Как может коренным образом меняться очень большое количество фенотипических признаков, не теряя своей структурной и функциональной интеграции, которой должен обладать любой жизнеспособный организм? Чтобы проиллюстрировать важность ископаемых остатков, я упомянул о происхождении перьев и волос. Они рассматриваются как «эволюционные новшества»: особенности, которые очень отличаются от прежнего состояния и инновационные, и потому они так или иначе вели к новой главе в эволюционной истории, или же коррелировали с ней. Другой пример — эволюция рук и ног, поскольку она документирована для самых ранних представителей группы, к которой принадлежат наземные позвоночные. С чем биологи согласны, и на чём они делают упор вновь и вновь, так это с тем, что для того, чтобы понять эти новшества, вы должны знать, как они развивались в ходе индивидуальной истории организма, который обладает ими. Мы знаем человеческую руку как оформленное и гибкое сочетание скелета, мускулатуры, кожи, нервов, кровеносных сосудов и других тканей. Чтобы рассмотреть происхождение руки, вы должны вернуться назад во времени к нашему эмбриональному периоду, в течение которого формируются различные её компоненты. Какова их связь с другими частями тела? Какие гены задействованы в их развитии? Как объединяются мускулы, сухожилия и кости? Как утверждал эволюционный биолог Гюнтер Вагнер, «объяснение эволюционных новшеств равнозначно идентификации изменений в ходе индивидуального развития, которые делают новый признак возможным». Нам следует предпринять ещё одно путешествие назад во времени, чтобы понять происхождение руки, и это будет путешествие по геологическому времени, а не по времени жизни индивида. Первые ископаемые позвоночные с мускулистыми руками и ногами жили почти 400 миллионов лет назад. Изучая те ископаемые остатки, мы можем узнать тот экологический контекст, в котором эволюционировали руки, и комбинацию анатомических особенностей, которыми обладали те животные. Мы знаем что у самых ранних позвоночных с руками и ногами было больше пяти пальцев на каждой конечности, и что они были водными животными. Теперь представьте себе, что мы смогли вернуться назад в геологическом времени, а затем вернуться назад во времени индивидуального развития ныне вымерших животных, чтобы понять, как возникли их руки. Это гипотетическое путешествие воплощает объединение двух дисциплин, которые в первом приближении могут показаться весьма различными: палеонтологии и биологии развития. Но их отношения чрезвычайно близки. Мы можем вернуться назад во времени в истории этих дисциплин и их отношений, и увидеть, насколько давно признаны параллели между индивидуальными изменениями и трансформациями, которые документируют ископаемые остатки.Отношения между палеонтологией и биологическими дисциплинами, изучающими индивидуальное развитие
Целые дисциплины или идеи возникают или утверждаются в канонах человеческого знания благодаря усилиям и успехам отдельных индивидуумов, у которых есть интерес, стимул и возможности заниматься тем или иным вопросом. Так, несомненно, произошло в случае с Жоржем Кювье, которого часто называют «отцом палеонтологии». Кювье получал образование главным образом в Штутгарте и жил в Париже в конце 1700-х — начале 1800-х годов.[7] В течение нескольких десятилетий он описал анатомию сотен ископаемых организмов, установив реальность процессов вымирания и изменения по мере течения геологического времени. Он жил до Дарвина и никогда не сочувствовал эволюционным идеям своего прежнего преподавателя Жана-Батиста Ламарка или же Этьена Жоффруа Сент-Илера, человека, очень интересовавшегося индивидуальным развитием организмов и столь же выдающегося, как и сам Кювье. Статуями или бюстами представителей всего этого ушедшего цвета французской науки может полюбоваться любой турист в Париже, посетив замечательный Музей Естествознания и Ботанический сад (Jardin des Plantes). Анатомические труды Кювье были широко известны, а его видный политический статус во время и после правления Наполеона также способствовал его заметности и распространению его энциклопедических работ. Главный труд Кювье, «Leçons d’anatomie», был переведен на немецкий язык известным анатомом, работавшим в Галле, Иоганном Фридрихом Меккелем-младшим. Собственные труды Меккеля также касалась анатомических сравнений, но они относились к эмбрионам, и его особенно интересовали уродства в процессе индивидуального развития. То же самое, что Кювье делал с ископаемыми организмами в Париже, Меккель делал с эмбрионами в Галле, когда собрал огромную коллекцию зародышей многих видов позвоночных животных. Его наследие можно увидеть даже в базовой анатомической терминологии: меккелев хрящ, значительная часть скелета нижней челюсти у эмбриона человека, и меккелев дивертикул, часть подвздошной кишки у некоторых людей. Меккель видел свою задачу как дополнение к работе Кювье в области анатомии, но с позиций эмбриологии. С него началась длинная череда биологов, изучающих эмбрионы. Меккель видел параллели между различиями среди взрослых особей ныне живущих и ископаемых видов с одной стороны, и различиями между эмбрионами одного и того же вида на разных временных отрезках в ходе индивидуального развития с другой стороны. Выяснение пределов, верности и значения этого потрясающего факта — это как раз то, с чем было связано многое в биологии на протяжении пары веков. Жаль, что Кювье и Меккель не дожили до наших дней, чтобы увидеть окаменелость, которая дала бы им шанс поразмыслить и над вымиранием, и над уродствами одновременно. Команда французских и китайских палеонтологов, работающая в северном Китае, опубликовала в 2007 году сообщение об открытии весьма необычного экземпляра рептилии, принадлежащей к хористодерам, которые представляют собой группу полуводных рептилий с неясными родственными связями. Найденный в породах мелового периода, знаменитых скрытыми в них замечательными окаменелостями крылатых динозавров и ранних млекопитающих, скелет прекрасной сохранности имеет две головы, каждая из которых связана со своим собственным набором шейных позвонков, и остальная часть скелета также полностью сохранилась. Свёрнутая поза этого и других сходных, но обладающих одной головой экземпляров предполагает, что эта особь представляет собой уродливый эмбрион. Принадлежность этих ископаемых остатков к группе Choristodera была определена главным образом по длинным шейным позвонкам и коротким конечностям — это две характерных особенности этой группы, известной по достаточно полным и многочисленным экземплярам взрослых особей из этой формации. Уродство, представляющее собой раздвоение головы/шеи, обычно для ныне живущих рептилий и называется аксиальной бифуркацией. Это состояние не обязательно фатально, поскольку многие черепахи и змеи по много лет выживают с ним в неволе. Очевидно, хористодеры обладают общей с ныне живущими рептилиями системой индивидуального развития, в которой мог возникнуть этот вид аномалии развития организма.
Рисунок 2. Реконструкция позднего эмбриона или новорождённой рептилии-хористодеры из меловых отложений Китая. Двухголовое состояние называется антериорной аксиальной бифуркацией. Общая длина животного, не считая хвоста, была приблизительно 40 мм. Рисунок Мадлен Гейгер, основанный на статье Buffetaut и др. 2007.
Возможность изучать уродства в развитии организма по ископаемым остаткам в большей степени возможна в царстве беспозвоночных, где палеонтолог собирает и изучает сотни, если не тысячи образцов. Позвоночные встречаются реже и в большинстве случаев требуют больше времени для препарирования. Параллели между эмбриологией и палеонтологией чаще всего ищутся не среди необычных окаменелостей, а скорее путём сравнения двух свидетельств или морфологических преобразований. Так происходило даже в случае некоторых учёных с антиэволюционным мировоззрением, как до, так и после Дарвина. Например, натуралист Луи Агассиц, родом из Швейцарии, ученик Кювье, занимался поиском отношений между «градациями» таксонов, преобразованиями в развитии организмов и геологической последовательностю окаменелостей. Под «градацией» он понимал то, что между таксонами существуют различия в той или иной степени, поэтому можно мысленно преобразовать один таксон в другой, если исследовать природу этих различий. Агассиц понял, что исследование индивидуального развития, а также изучение ископаемых остатков в контексте геологического времени представляют собой параллели, проясняющие понимание этих преобразований, даже если он не думал, что механизмом, в конечном счёте ответственным за это, была эволюция. Пример можно прочитать в его крупном труде «Recherches sur les poissons fossils», изданном несколькими томами в период между 1833 и 1843 гг. Агассиц утверждал, что хвост взрослых ископаемых лучепёрых рыб, группы, к которой принадлежат почти все современные виды рыб, кроме целакантов и двоякодышащих, демонстрирует параллели с развитием хвоста современных видов этой группы от эмбрионов к взрослым особям. Молодые особи имеют простой хвост, у более поздних стадий имеется гетероцеркальный хвост, в котором верхняя часть крупнее, чем нижняя, а взрослая особь обладает хвостом, у которого обе части сходного размера. Эти изменения подобны последовательности, которую Агассиц описал для ископаемых форм. То, что черты различия, а также и сходства между видами были результатом общего происхождения с изменениями, произошедшими эволюционным путём, было признано многими другими биологами после Дарвина, в особенности явно немецким интеллектуалом Эрнстом Геккелем, которому было двадцать пять лет в 1859 году, когда книга «Происхождение видов…» была впервые издана. Как утверждает его самый новый и осведомлённый биограф, в Европе, в том числе в Англии, больше людей прочитало об эволюции в книгах Геккеля, чем в книгах самого Дарвина или любого другого автора. Геккель был первым человеком после 1859 года, кто открыто опубликовал эволюционное древо с существующими в природе группами организмов. Геккель был также склонен к популяризации фактов и в конечном счёте главным образом его считают ответственным за случившуюся позже историю с утверждением «онтогенез рекапитулирует филогенез».[8] Этот принцип заключает в себе идею о том, что индивидуальное развитие так или иначе отражает в себе различия между видами. Это утверждение имеет историю, восходящую даже к главному учителю Кювье, Карлу Фридриху Кильмейеру из Тюбингена, чьи лекции в университете, как говорили, привлекали целые толпы людей. Параллель между развитием и эволюцией была классическим предметом исследований и обсуждения в немецкой биологии, что отразилось и в немецком языке: одно и то же слово «Entwicklung» может использоваться для описания как эволюции, так и индивидуального развития.[9]Параллели между трансформациями в палеонтологии и в индивидуальном развитии
Рука раннего человеческого зародыша выглядит как весло, поверхностно напоминая конечность наших водных предков. Существует много других примеров общего сходства между палеонтологическими изменениями и изменениями, связанными с индивидуальным развитием. Наиболее знаменитый из них — это тот, который относится к эволюции и развитию наших нижней челюсти и среднего уха, особенностей, которые мы разделяем со всеми остальными млекопитающими.
Рисунок 3. Зародыши человека различного возраста на протяжении первых трёх месяцев развития, какими они изображены в классической работе Ф. Кейбеля (1906), частично по работе Вильгельма Гиса. Не в одном масштабе.
Этот случай часто приводится как выдающееся достижение сравнительной биологии, и потому его стоит рассмотреть более подробно. У млекопитающих имеется единственная кость, образующая нижнюю челюсть; она называется зубной костью — подходящее имя, поскольку это как раз то место, где расположены зубы. У всех прочих позвоночных в челюстях имеется много костей. Млекопитающие обладают тремя слуховыми косточками, тогда как у других позвоночных со сходным типом уха есть только одна. Хотя у нас может даже мысли не возникнуть о том, что наши челюсти и уши как-то связаны, фактически это именно так. Млекопитающие — большие оригиналы с точки зрения анатомии, и мы, основываясь на дарвиновском понимании эволюционной биологии, можем сделать вывод о том, что наши ранние предки, должно быть, в какой-то точке нашей истории были подобны прочим позвоночным. В действительности окаменелости показывают, что наш общий с рептилиями предок, который жил, по крайней мере, около 315 миллионов лет назад, обладал несколькими челюстными костями и простым средним ухом с единственной слуховой косточкой. Первые окаменелости, документирующие преобразования, которые привели к анатомии современных млекопитающих, были найдены в Южной Африке и в России в 1840-е годы, но с тех пор ископаемые остатки самых ранних предков млекопитающих были обнаружены на всех континентах. На протяжении истории млекопитающих челюстные кости, кроме зубной кости, подверглись редукции, и в процессе эволюции образовался сустав между зубной костью и черепом. Прежний челюстной сустав становился всё меньше и меньшие и постепенно отделился от нижней челюсти. Сочленение между костями сохранилось, но с другой функцией, а именно для передачи звуковых волн, и со временем оно приобрело специализацию к передаче высокочастотных звуков. Исходные сочленовная и квадратная челюстные кости называются у млекопитающих malleus и incus как элементы уха, или, говоря более простым языком, это молоточек и наковальня. Вместе со stapes (стремечком) эти кости образуют три косточки среднего уха млекопитающих. Связи между косточками образуют систему рычагов, с помощью которой высокочастотные звуковые волны передаются к клеткам внутреннего уха, и в конце концов к мозгу для заключительной обработки. Эти косточки уха млекопитающих, некогда бывшие частью жевательного аппарата у наших вымерших предков и у наших современных кузенов рыб, по размеру не намного больше, чем мелкие камешки. По крайней мере, у некоторых млекопитающих было зарегистрировано существование двух челюстных суставов на ранних стадиях индивидуального развития: исходный для челюстноротых позвоночных и всё ещё характерный для рептилий, и новый, имеющийся лишь у млекопитающих — между зубной костью, нашей единственной костью нижней челюсти, и чешуйчатой костью черепа. Кости среднего уха прикреплены с помощью меккелева хряща к внутренней стороне зубной кости. Некоторые виды из раннего триаса демонстрируют точно такие же особенности строения: два одновременно существующих челюстных сустава. Первая из найденных окаменелостей, показывающая эту замечательную особенность, происходит из Южной Африки и Альфред Кромптон из Гарвардского Университета[10] в 1958 году дал ей название Diarthrognathus. Другое «раннее млекопитающее», хорошо известное палеонтологам — Morganucodon из позднего триаса/ранней юры, который обладал типичным для млекопитающих челюстным суставом только между зубной и чешуйчатой костью, но имел видимые остатки «старой челюстной анатомии». Morganucodon был мелким, размером с землеройку, и, хотя теперь он известен из Евразии, Африки и Северной Америки, первоначально он был описан из местонахождений в Англии и Уэльсе.
Рисунок 4. Развитие нижней челюсти и косточек среднего уха у сумчатого млекопитающего Monodelphis domestica. Серия образцов показывает постепенное уменьшение относительного размера элементов, которые станут ушными косточками у взрослой особи. Медиальный вид молодой особи из сумки демонстрирует длинный и заметный меккелев хрящ, который становится относительно меньше, когда животное растёт. Не в одинаковом масштабе. По Rowe 1996; и Luo 2007, 2011, с изменениями.

Рисунок 5. Челюстной сустав у «раннего млекопитающего» Morganucodon. Череп показан с вентральной стороны, а нижняя челюсть — с медиальной стороны. Длина черепа — около 2,5 см. Рисунок любезно предоставлен Ч.-С. Луо, с изменениями (см. Luo 2007).
Этот замечательный пример параллелей между онтогенезом и филогенезом из летописи окаменелостей не подразумевает того, что одно может использоваться для заполнения пробелов в другом. Общее подобие обнаруживается в изобилии, но отклонения также обычны. Когда черты сходства присутствуют, они иногда кажутся поразительными, но при более пристальном рассмотрении оказываются лишь поверхностными. Молодая особь сумчатого, всё ещё прикрепляющаяся к своей матери, обладает двойным челюстным сочленением на стадии, когда череп ещё не окостенел и расположение мускулатуры и мягких тканей ещё плохо различимо. Это довольно сильно отличается от анатомии взрослого Diarthrognathus (с двумя челюстными суставами), который бродил примерно 200 миллионов лет назад по тем местам, которые сегодня являются южноафриканской пустыней Карру.[11]Онтогенез обычно не является рекапитуляцией филогенеза
Тот факт, что онтогенез не является простой рекапитуляцией филогенеза, достаточно общепризнан даже при том, что общие параллели находятся в изобилии. Чтобы понять, что означает явление рекапитуляции в том смысле, в каком его определил аналитическим путём Геккель, мы используем простую схему. Чтобы произошла истинная рекапитуляция, дополнение должно было бы возникнуть в конце исходной, или предковой последовательности появления признаков или на возможном продолжении траектории. На рисунке 6 у нас изображены четыре вида, каждый из которых характеризуется обычным путём индивидуального развития, состоящим из первого шага, M1-M1. У каждого вида новая стадия добавляется в конце последовательности. В этом идеальном случае вид с наиболее специализированным состоянием, вид «D», сохраняет в своём онтогенезе последовательность эволюционных преобразований, которые совершились путём добавления новых стадий в конечные этапы развития. В ходеэволюции случаются отклонения от гипотетического хода рекапитуляции, что признавал и сам Геккель. Эти изменения означают, что последовательность появления признаков в цепочке может сдвигаться, один или больше признаков могут исчезать, или же может возникнуть совершенно новый признак. Эти изменения иногда настолько драматичны или считаются настолько важными, что их рассматривают как эволюционные новшества; так произошло в случаях происхождения волос у млекопитающих или перьев у динозавров. Существуют различные виды отклонений от рекапитуляции, среди них гетерохрония (изменения во времени появления) и гетеротопия (изменения в пространственном расположении в структуре).
Рисунок 6. Последовательный характер добавления новых признаков на конечных стадиях в эволюции онтогенетических последовательностей приводит к онтогенетической рекапитуляции, но имеются отклонения от этого способа преобразований. По Wägele 2005, с изменениями.
Некоторые палеонтологи исследовали эволюционные преобразования, документированные ископаемыми остатками, и сравнили их непосредственно с онтогенетическими преобразованиями, отыскивая параллели или их отсутствие. Например, изменения в развивающемся черепе эмбрионов аллигаторов очень схожи с теми, которые наблюдаются в ряду ископаемых остатков ранних крокодилов. Это сходство не просто поверхностное, но соотносится со статистически существенной корреляцией в порядке, в котором в обеих последовательностях происходят дискретные изменения тех же самых признаков. Итак, несомненно, что существуют параллели между онтогенезом и летописью окаменелостей, показывающих морфологические изменения в некоторых структурах у некоторых животных. Но это исключения.Эволюция онтогенеза
Каждый этап развития в жизни организма может подвергнуться изменениям в процессе эволюции. Этого и следует ожидать, потому что живые организмы подвергаются давлению со стороны окружающей среды на каждой стадии и в каждый момент своей жизни. Головастик противостоит таким обстоятельствам, как взаимоотношения с хищниками или поиск пищи, и на этой стадии в процессе эволюции возможно появление специализаций. Фактически, при изучении лягушек и других амфибий,[12] таких, как саламандры, было выявлено ошеломляющее разнообразие эмбриональной морфологии и видоспецифичных особенностей индивидуального развития. Кристиан Митгатч, ранее работавший в моей лаборатории в Цюрихе, и научный руководитель его работы на соискание степени доктора философии Леннарт Олссон, в Йене (Германия), в том же месте, где Геккель сделал свою долгую и знаменитую академическую карьеру, продемонстрировали большое разнообразие у ранних эмбрионов лягушек, проявляющееся в разном времени развития клеточных предшественников многих черепных структур. Они осуществили это, прослеживая движение клеток из нервного гребня, который у позвоночных является зародышевым листком, ответственным за развитие многих тканей, специфичных для позвоночных. Эти ткани включают, например, элементы зубов и много черепных костей. Работа Митгатча и Олссона дала нам прекрасные изображения эмбрионов, показывающие потоки клеток, мигрирующих в разное время и в различных пропорциях у разных видов (рисунок 7).
Рисунок 7. Различная степень развития сегментов, или повторяющихся блоков эмбриональных тканей тела (сомитов) в начале миграции клеток нервного гребня у эмбрионов лягушек Bombina orientalis и Dendrobates azureus. Фото любезно предоставили Кристиан Митгатч и Леннарт Олссон.
Самые ранние сухопутные позвоночные, которые населяли Землю, представляют собой превосходный палеонтологический пример изменений на разнообразных стадиях онтогенеза. Среди этих животных были некоторые из предков современных лягушек, саламандр и группы безногих амфибий. Наиболее разнообразными среди этих первых «амфибий» были темноспондилы (Temnospondyli), группа, существовавшая с нижнего карбона, примерно 340 миллионов лет назад, до нижнего мела, около 120 миллионов лет назад. Большая часть времени их существования перекрывается со временем существования наземных динозавров. Самые мелкие из них были менее 30 сантиметров в длину, но некоторые формы напоминали крокодилов и достигали длины около 7 метров. Ювенильные и личиночные стадии описаны для многих темноспондилов и они показывают большое разнообразие экологии личинок и характера роста. Все темноспондилы выводили своё потомство в воде, но типы водных местообитаний были различными. Некоторые группы предпочитали или выдерживали солоноватую, или даже морскую воду; другие ограничили своё распространение озёрами.[13] Эти личинки были исключительно плотоядными, на что указывают характер их зубов и, в некоторых случаях, мощные челюсти. Другая ископаемая фауна, зарегистрированная на участках, где были обнаружены эти окаменелости прекрасной сохранности, а также подробное изучение анатомии жевательного аппарата или случайно сохранившегося содержимого желудка темноспондилов показывает, что некоторые из них охотились на рыбу, тогда как другие поедали различных мелких «амфибий». Замечательная сохранность разнообразных жизненных стадий у темноспондилов отчасти объясняется их большим размером, что повышает вероятность их сохранения, а также делает их легче заметными для палеонтологов во время работы в поле.[14] A родственная группа, лепоспондилы, была, как и самые ранние «амфибии», представлена мелкими формами: в размерном диапазоне от 5 до 20 сантиметров. Об онтогенезе лепоспондилов почти ничего известно, и это бросает вызов палеонтологам, пытающимся точно установить их родственные связи и происхождение современных земноводных, используя данные об индивидуальном развитии. Значительное разнообразие путей онтогенеза демонстрирует группа одних из самых ранних позвоночных с ногами, или четвероногих, похожих на тритонов — бранхиозавриды. Почти все двадцать видов этой группы сохраняли личиночные особенности, такие, как наружные жабры или зубчики на жаберных дугах во взрослом состоянии; это неотенические признаки. Некоторые другие виды претерпевали метаморфоз и утрачивали свои личиночные признаки во взрослом состоянии. Все представители группы были мелкими, в противоположность прочим темноспондилам. Тысячи экземпляров некоторых бранхиозаврид известны из местонахождений в Германии, включая одно в бассейне Тюрингенского Леса. Там Ральф Вернебург много лет изучал ископаемые остатки темноспондилов, таких, как бранхиозавры. Некоторые виды бранхиозаврид обладали особенностями, которые позволяли считать их ручьевыми формами; другие, с видимыми наружными жабрами, описаны как озёрные формы. Один из спорных вопросов для экспертов — могут ли в некоторых случаях два типа возникать как различные экоморфотипы одного и того же вида, продвигая онтогенетическую изменчивость ещё на один шаг вперёд, демонстрируя случай «фенотипической пластичности», тему, которую я обсуждают в главе 2.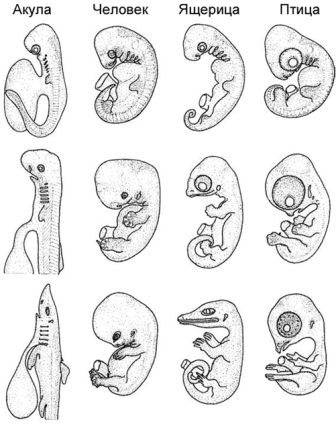
Рисунок 8. Три стадии развития у акулы, человека, ящерицы и птицы. По Westheide и Rieger 2009, с изменениями.
Как и в вышеприведённых примерах, любая стадия на протяжении развития индивидуума может изменяться. Это демонстрировалось опытным путём для многих видов позвоночных и стадий процесса. Эволюционируют все стадии онтогенеза, а не только взрослые, объект приложения усилий многих исследователей. Многие авторы представляют данные и идеи, призванные подвергнуть критике «соломенное чучело», защищающее наивную рекапитуляционную идею, согласно которой онтогенетические и эволюционные преобразования точно отражают друг друга. И важнее всего тут то, что желание исследовать фундаментальный аспект того, как эволюционировало индивидуальное развитие, привело к пристальному изучению организмов из многих групп, и итогом этого стало несколько значительных открытий. Например, Майк Ричардсон и немного позже Ингмар Вернебург документировали внешнюю морфологию эмбрионов, которая уже в первом приближении показывает, что в более или менее сопоставимые периоды индивидуального развития имеются существенные различия в пропорциях и формах, а также в размере желтка у рыб, змей, лягушек, птиц, ящериц и млекопитающих, в том числе людей. Эта особенность — лишь одна из многих, иллюстрирующих различные физиологические и морфологические особенности на протяжении времени индивидуального развития и у различных видов.[15]Онтогенез и условия для поддержания жизни
Если индивидуальное развитие и рассказывает нам историю, то она похожа на те страшные истории, которые рассказывают дети, сидящие вокруг походного костра, когда каждый мальчишка добавляет свой отрывок к тому, что уже было до него. Нет ни одного единого рассказчика, и нет внутреннего диалога. Есть только то, что уже произошло, что происходит сейчас, и что произойдёт потом.Изменения в тканях, которые происходят в ходе индивидуального развития, протекают для живого эмбриона в его настоящем времени. С этой точки зрения может показаться, что взрослые особи — это почти случайность, или же один из нескольких моментальных снимков жизни. Это не означает, что особенности какой-либо стадии развития случайны; они являются результатом процессов, которые мы можем во многих случаях количественно определить и предсказать — продуктом чётко скоординированных и детерминированных механизмов, где пластичность и случайность играют главную роль в формировании результата. Есть генетическая система, отвечающая за индивидуальное развитие, которая работает в контексте окружающей среды. Действие этой системы завершается там, где начинается фенотипическое проявление процессов индивидуального развития, которое управляется физическими и химическими законами, которые работают в природе. Объяснение существования того или иного признака на любой из стадий развития, отличающейся от взрослого состояния, обычно делается в рамках понятий будущего состояния, к которому движется животное. Как утверждал Джон Реисс в своей книге «Not by Design», объяснение существования признака должно делаться в рамках его необходимости на момент его существования, но не в терминах будущего состояния, движению к которому он способствует. Чтобы понять этот основной принцип, полезнее всего обратиться к идее «условий для поддержания жизни», выдвинутой Кювье. «Условия», о которых говорил Кювье — это особенности живого существа, без которых оно не могло бы выжить. Организм не может существовать, пока не будут удовлетворены его условия для поддержания жизни. Это может показаться тавтологическим или тривиальным, но, подобно многим великим истинам, идея становится фундаментальной лишь после того, как она была полностью признана. Эта идея имеет важные значения для концептуальных рамок, которыми мы пользуемся, чтобы исследовать эволюцию индивидуального развития. Например, при рассмотрении экологических аспектов эволюционного разнообразия понятие «ниша» часто используется так, словно она должна была существовать в окружающем мире, и словно это цель, к которой стремится группа организмов. Как обсуждалось ранее, ниша не существует независимо от организмов, которые фактически создают её. Среди любых форм жизни организм воздействует на окружающую среду, но также подвергается воздействию с её стороны.[16] Когда образуется новый океанический остров, со временем некоторые организмы, случайно оказавшиеся там, будут вести себя так же. Они не будут поступать так в соответствии с неким планом или целью; они просто будут пытаться выжить. Большинство биологов не настолько наивно, чтобы думать, что у живых организмов есть план, но изложение часть строится на основе разработанного плана. Естественный отбор ограничивает эволюционирующую совокупность, «выбирая» без плана среди множества экспериментов, которые индивидуальное развитие проделывает в новом контексте окружающей среды. Поскольку живые организмы — это мозаика признаков, которые все объединены, но некоторые — теснее прочих, индивидуальное развитие под действием естественного отбора будет эволюционировать сложным образом. Эволюционные изменения у островных видов иногда объясняются как результат остановки или замедления развития, при которых островная форма просто напоминает ювенильную стадию своего предка. Но живые организмы мозаичны. Каждый орган или часть тела, хотя и составляет единое целое с остальными, управляется различными правилами относительного роста. Островной вид может быть в общих чертах подобен ювенильной особи предкового вида, но это может быть лишь поверхностным сходством, или же быть верным лишь для некоторых особенностей. «Мозаика» образуется в каждый из моментов эволюционного времени из-за того, что условия для подержания жизни должны удовлетворяться на каждой из стадий индивидуального развития. Я обсудил несколько параллелей между биологией развития и палеонтологией по предмету и по подходу. Ещё одна — это телеологическая позиция, которая утверждает, что в основе изменений лежит замысел, и он направлен в сторону улучшения. Это та позиция, которая преобладала в этих двух дисциплинах. Идея наличия замысла и цели связана с телеологией, термином, доставшимся нам от греческих философов, которые первыми обсуждали в различных контекстах вопросы наличия предназначения и цели у жизни. Если даже кто-то и принимает принципы множества религий или иных форм веры в сверхъестественное, телеология совершенно неуместна в любой научной дискуссии, касающейся эволюции. И всё же поиск объяснений и незнание принципов самоорганизации жизни привели в прошлом к телеологической концепции жизни. Изучение ископаемых остатков не поддерживает телеологическое представление о жизни. Летопись окаменелостей демонстрирует многочисленные образцы отличающихся друг от друга и параллельных морфологических изменений. Классические примеры — это ветвистое эволюционное дерево лошадей, берущее своё начало в раннем эоцене, примерно 55 миллионов лет назад, или родословная людей, когда примерно два миллиона лет назад одновременно жило пять видов гоминид, но лишь один из них существует сегодня. Формы телеологического мышления, которые по современным стандартам выглядят весьма причудливыми, были весьма обычными в истории биологии. Во многих случаях утверждается о существовании некоторого рода жизненной силы, которая осуществляет эволюционное изменение в предопределённом направлении, будучи вызванной свойствами, присущими организмам. Примеры такого рода — принцип врождённого стремления к совершенствованию Ламарка и идея «ортогенеза»; к последней обращался на страницах своих книг влиятельный биолог Теодор Эймер. Эти идеи витализма и внутренних движущих сил жизни очень повлияли на палеонтологию. Идею эволюции, идущей посредством «аристогенеза», вида ортогенеза, подразумевающего постоянное совершенствование, отстаивал, например, Генри Ф. Осборн, ведущая фигура в американской палеонтологии, много лет занимавший должность президента Американского Музея Естественной Истории в Нью-Йорке в первые десятилетия двадцатого века.[17]Марк С. Блумберг «Freaks of Nature»
Онтогенез и филогенез
Некоторые иллюстративные материалы весьма поучительны и помогают глубже понять отношения между онтогенезом и филогенезом. Я приведу здесь два своих любимых примера. Первый принадлежит немецкому ботанику Вальтеру Циммерману (1892–1980), который, помимо своей работы в области морфологии растений, был оригинальным и влиятельным писателем на темы теории эволюции и методов биологической классификации. На варианте «схемы Циммермана», воспроизведённом здесь (рисунок 9), есть спирали онтогенеза, включающие в себя различные стадии жизни, изменяющиеся в ходе геологического времени. Филогенез является эволюцией онтогенеза, в данном случае он проиллюстрирован происхождением птиц, с символичным археоптериксом, «недостающим звеном между птицами и рептилиями» из Баварии, реконструкция которого помещена на несколько ступеней ниже текущего состояния онтогенеза птиц. Полный спектр изменений во времени, включающий и филогенез, и онтогенез, Циммерман называет «гологенезом», термином, который никогда не был общепринятым в стандартной терминологии на моём поле деятельности.
Рисунок 9. Отношения между онтогенезом и филогенезом, как их изобразил Вальтер Циммерман. По Циммерману, 1968, с изменениями.
Другую схему, которая прекрасно иллюстрирует эволюцию, создал Вольфганг Майер, мой бывший наставник в Тюбингенском Университете. Среди его самого значительного вклада в науку — исследования анатомии млекопитающих ближе к моменту рождения, когда новорождённые сталкиваются с новыми испытаниями со стороны окружающей среды. Как любит замечать Майер, разные виды приобрели в процессе эволюции различную анатомию на этой стадии развития, и это же справедливо для всех стадий, от зиготы до смерти — на протяжении истории всей жизни. Естественный отбор действует на всех стадиях, и в процессе эволюции меняются пакеты генов, структуры и функции, работающие на разнообразных уровнях биологической организации. В некоторых случаях это изменение происходит постепенно, в сумме приводя к крупным эволюционным преобразованиям по ходу течения геологического времени. Генетические изменения передаются следующим поколениям посредством полового размножения.
Рисунок 10. Отношения между онтогенезом и филогенезом, изображённые Вольфгангом Майером. По Maier 1999, с изменениями.
Можно использовать много переменных величин, чтобы охарактеризовать процессы онтогенеза, представленные в абстрактной форме на этих рисунках. Среди них продолжительность эмбриональной и ювенильной стадий, возраст полового созревания, состояние в момент выхода из яйца или при рождении, а также репродуктивная долговечность. Все они входят в историю жизни животного, и каждая из них может эволюционировать. Аналогичным образом эволюционирует также история жизни у растений. В случае с ныне живущими видами способность связывать эти переменные величинами с исследованиями в области популяционной динамики важна для вопросов охраны природы, поскольку они могут выделить стратегии управления для тех организмов, которые рассматриваются как находящиеся в зоне риска. С эволюционной точки зрения знание этих переменных величин может помочь нам понять, почему определённые группы вымирают, тогда как другие становятся многочисленными и разнообразными, или же просто выжили. Некоторые из этих переменных также связаны с крупными анатомическими новшествами, как крупный мозг людей отчасти стал возможным благодаря долгому периоду беременности. Ископаемые остатки являются ключом к нашему знанию о том, когда возникли эти особенности, даже если они не могут предоставить прямую информацию, касающуюся некоторых переменных величин, вроде продолжительности беременности. Это становится возможным, потому что большое количество эмпирических данных показало тесную ассоциацию с некоторыми анатомическими особенностями, которые сохранились в ископаемом виде вместе с деталями истории жизни. Как в случае с ранними наземными позвоночными, которые обсуждались выше, окаменелости, сохранившиеся наилучшим образом, являются первыми и наиболее очевидными объектами изучения, позволяющими документировать эволюцию истории жизни индивида в геологическом прошлом.Онтогенез и реконструкция древа жизни
Со времён новаторской работы Эрнста Геккеля и с теорией эволюции в качестве точки отсчёта основной задачей была реконструкция древа жизни. История подходов и проблем длинна и сложна. Вопрос, который неоднократно оказывается на поверхности в ходе дискуссий, состоит в том, как дополнить это древо информацией об онтогенезе. При использовании морфологии вывод об отношениях между видами обычно делается путём сравнения наличия и специфичности деталей различных особенностей строения. В случае специалистов по позвоночным мы главным образом исследуем кости, мышцы и нервы взрослого животного. Но наследуемые и отличающиеся от вида к виду особенности, которые необходимы для анализа, позволяющего реконструировать родословное древо, присутствуют на всех стадиях жизненного цикла животного, поэтому нет никаких причин ограничиваться только взрослыми особями в качестве источника информации об анатомии. Онтогенез, таким образом, важен, но как же его правильно использовать? Большинство практикующих систематиков, биологов, чья деятельность непосредственно связана с реконструкцией родословного древа и классификацией, согласилось бы со следующим рядом утверждений, который претендуют на обобщение вопросов, в отношении которых специалисты большей частью достигли консенсуса. 1. Подход, основанный на изучении исключительно взрослых стадий с большей степенью вероятности приведёт к заблуждениям при осуществлении филогенетического анализа, нежели подход, охватывающий большую часть истории жизни особи у изучаемого вида. Это связано с повышением уровня комплексности анализа особенностей организмов (признаков), который преодолевает значительную атомизацию черт поверхностного морфологического сходства.[18] Пример уместности ювенильных стадий для определения родственных отношений между организмами демонстрируют нам животные, которых изучал сам Чарльз Дарвин. Морских желудей долгое время считали моллюсками из-за их известковой раковины у взрослых особей. Но личинки морских желудей похожи на личинок ракообразных, и эти две группы в настоящее время считаются близкими родственниками{2}. 2. Онтогенез в отдельности не может использоваться в качестве критерия для установления направления эволюционного изменения той или иной особенности. Систематики стремятся определить, какие особенности являются предковыми, а какие — специализированными, то есть «полярность» изменений. Если бы эволюционные преобразования всегда осуществлялись путём добавления новых шагов в конце исходной онтогенетической последовательности, или же путём продления онтогенетической траектории (идеальный случай, изображённый выше), то сравнение хода онтогенеза указало бы направление преобразований. Но рекапитуляции — это скорее исключение, чем норма. Сравнения с группами, близкородственными рассматриваемой, служат для реконструкции системы родственных отношений. Как только обнаружено начало системы, становится возможным построение древа родственных отношений, по которому определяется полярность состояний признака. 3. Не все группы организмов обладают схожими процессами роста, поэтому природа независимости или морфометрических отношений между частями тела не универсальны. Различия, о которых я упоминаю здесь — главный предмет исследований в лаборатории моего цюрихского коллеги Хьюго Бюше, который совместно с Клодом Монне и другими коллегами разрабатывает новые математические модели, чтобы понять рост аммонитов, которые являются вымершими моллюсками. У моллюсков, как, например, у брахиопод, или плеченогих{3}, очертания раковины формируются посредством добавочного роста по краям отверстия увеличивающегося диаметра, с серией связанных с ним кривых. В противоположность этому, у трилобитов и других членистоногих происходят линька и добавление сегментов. У иглокожих, к которым относятся морские звёзды и морские ежи, исходная двусторонняя симметрия преобразовалась в пятилучевую, что привело к модификациям процесса роста. Как я говорю в главе 9, вполне очевидно, что образование формы тела у аммонитов, трилобитов и иглокожих происходит различными путями, что должно приниматься во внимание при реконструкции эволюционных историй этих групп. Когда проводятся количественные исследования филогенеза или реконструкция эволюционного древа, сложная форма организмов обычно подразделяется на отдельные особенности (атомизация).[19] Однако в этих исследованиях должен рассматриваться рост, и установление того, как наилучшим образом объединить эту линию свидетельств — серьёзный вызов для специалистов по систематике.Глава вторая Эво-дево, пластичность и модули
Дисциплину, которая объединяет области ведения генетики развития и эволюции, окрестили «эво-дево»{4}. Очень немногим новым аспектам эволюционной биологии было уделено столько же внимания от практиков и философов от биологии, если такое внимание вообще оказывалось. Предполагалось, что «эво-дево» создаст новый вид синтеза знания, который поможет понять происхождение биологического многообразия. По этой причине важно подумать над тем, как палеонтология вносит свой вклад в эту область знания. Главный объект внимания «эво-дево» ещё является объектом споров. Некоторые видят его в объяснении способности эволюционировать, или эволюабельности. Многие видят его в объяснении эволюционных новшеств или инноваций в смысле действительно новых, крупных шагов в морфологических изменениях — например, эволюцию глаз, зубов, конечностей или черепашьего панциря.[20] Другие видят в «эво-дево» лишь «мимолётную комету», дисциплину, которая будет вытеснена более новым слиянием областей знания, или новым порядком групп исследователей и тем, возможно, продиктованным подъёмом геномики.[21] Но, как бы то ни было, никто не отрицает того, что эти дискуссии стали источником многих размышлений и импульса к изучению индивидуального развития с эволюционных позиций. Начала «эво-дево» были во многом связаны с выяснением того, что общего имеют друг с другом многочисленные отдалённо родственные группы, как, например, мухи и люди, или несопоставимые элементы анатомии, как зубы и конечности. То, что глаз мухи и глаз позвоночного образуются благодаря действию сходных молекулярных механизмов — это крупное открытие. Выяснение того факта, что некоторые молекулы принимают участие в развитии и зубов, и конечностей — это тоже важное достижение. Но эти образования, начиная с момента своего появления, имеют различные формы и цвета, и именно это является захватывающим моментом. «Эво-дево» изменила акценты с попыток понять, почему организмы столь различны, на вопрос о том, как могли возникнуть эти несходство и разнообразие, если помнить о широко распространённом явлении генетического консерватизма, характеризующего эти организмы. Понимание отношения между генотипом и фенотипом — основное в этом предприятии. В прошлом многие биологи предполагали, что отношения между генотипом и фенотипом прямолинейны и просты, когда жизнеспособная мутация непосредственно приводит к новому фенотипу. На всех уровнях организации, даже на молекулярном, это оказалось неверным. Уже сто лет назад некоторые биологи писали о норме реакции, явлении, при котором один и тот же генотип способен образовывать различные фенотипы в зависимости от внешнего воздействия со стороны окружающей среды.[22] Эта идея была во многом забыта ведущими эволюционными биологами, но теперь она является центральной для исследований индивидуального развития с эволюционных позиций. Область ведения недавно окрещённой экологической биологии развития, прекрасно описанная в учебнике Скотта Гилберта и Дэвида Эпела, суммирует множество данных, касающихся взаимодействия между окружающей средой и индивидуальным развитием. Два аспекта разнообразия, которые стали центральными темами исследований «эво-дево» — это фенотипическая пластичность и модульность. О том, что означают эти важные концепции и какое отношение имеет к ним палеонтология, говорится далее.Фенотипическая пластичность и изменчивость
Развитие организма — это не река генетической информации, неуклонно текущая по своему руслу в сторону создания биологической формы, а скорее множество рек, притоков и водоворотов — бурный, циклический процесс, задействующий генную регуляцию и синтез белка.Марк С. Блумберг «Freaks of Nature»
Ничто в изменчивости не имеет смысла вне контекста развития организма.У изменчивости есть негенетический компонент, и фенотипическая пластичность относится к явлениям изменчивости, вызванным воздействием окружающей среды. Такое воздействие могут оказывать физические факторы, такие, как температура, или биотические факторы вроде взаимодействия с другими видами, или стимулы типа рациона. Конечно, это очень важно, поскольку изменчивость — это как раз то, на что действует естественный отбор. Чем больше изменчивость, тем активнее происходит эволюция и тем больше разнообразие.[23] Фенотипическая пластичность в характере роста, связанная с факторами окружающей среды, была отмечена для нескольких видов динозавров, в том числе для прозауропода Plateosaurus. В периоды изобилия пищи или благоприятного климата животные росли быстрее. Такой вид пластичности характерен не для всех прозауропод; он отсутствует, например, у близкородственного Massospondylus. Этот вид исследований можно предпринять только для того вида, для которого известно большое количество ископаемых остатков, достаточное для построения графиков роста с использованием палеогистологии для оценки возраста отдельных экземпляров.Юкка Йернвалл

Рисунок 11. Старое и новое представление отношений между генотипом и фенотипом. На основе сходного рисунка из Sholtis и Weiss (2005).
Фенотипическая пластичность предполагалась для нескольких групп ранних наземных позвоночных палеозоя; это мнение основано на тщательных анатомических и палеоэкологических исследованиях сотен экземпляров. Райнер Шох в Штутгарте продемонстрировал на прекрасных по своей сохранности ископаемых остатках темноспондилов, что один и тот же вид был способен изменять свою онтогенетическую стратегию в зависимости от условий окружающей среды — это же известно для многих современных саламандр. Данное животное, Sclerocephalus, достигало максимального размера в 2 метра и на первый взгляд напоминало странного крокодила. В больших мелководных озёрах, которые были бедны пищей, Sclerocephalus затормаживался в развитии, что приводило к появлению мелких взрослых особей, поедавших мелкую добычу. В озере с более сложной экосистемой, где и численность, и количество видов пищи были больше, Sclerocephalus имел более продолжительную стадию роста, становился крупнее и мог бы даже рисковать выходить на сушу, хотя содержимое желудка показывает, что это было преимущественно водное существо. В другом доисторическом озере, отложения которого сохранили очень крупных и прожорливых акул, были найдены только личинки, и это заставляет предполагать, что взрослые особи жили где-то ещё, возможно, чтобы избежать хищников. Связь между экологическими условиями и изменениями в онтогенезе была установлена для этих палеозойских позвоночных на основании надёжных свидетельств.
Рисунок 12. Скелет Sclerocephalus haeuseri из бассейна Саар-Нахе, юго-западная Германия, примерно 297 миллионов лет. Изображение любезно предоставил Райнер Шох (Штутгарт).
Sclerocephalus явно проявлял пластичность в своей стратегии развития; у других ранних наземных позвоночных, также известных по сотням экземпляров из различных местностей, она проявляется гораздо слабее. К сохранению таких ископаемых остатков и их открытию привели исключительные обстоятельства. Болота и озёра зачастую являются хорошими местами для захоронения ископаемых остатков, и в некоторых случаях бескислородные условия илистого дна этих водоёмов, со слабой биологической переработкой органики привели к превосходному сохранению в ископаемом виде костей и даже мягких тканей мёртвых тел этих животных. Поскольку онтогенез отдельно взятой особи у этих ископаемых животных нельзя проследить непосредственно, как это можно сделать сегодня у лягушки или саламандры, палеонтологам приходится опираться на тщательное анатомическое сравнение многочисленных экземпляров, и, хотя определение различных форм с точностью до вида может быть проблематичным, обычно мы можем идентифицировать этих животных с высокой степенью достоверности. Палеонтологи традиционно интересовались изменчивостью из-за потребностей таксономического характера. Чтобы определить границы вида по ископаемым остаткам, требуется установить максимальную степень морфологической изменчивости, которую может проявлять вид. Для этой цели подходит метод оценки или измерения некоторых черт строения у как можно большего числа индивидуумов, а сравнение с соответствующими ныне живущими видами и рассмотрение онтогенетической изменчивости являются основополагающими. Другой необходимый аспект — это знание эволюционного древа, в которое включался бы новый вид, и вместе с этим установление продвинутых признаков, которые характеризуют этот вид. Форма и размеры костей изменяются в пределах вида, и даже в пределах популяции. Но не все элементы скелета изменяются в одинаковой степени. Один из факторов, который может оказывать влияние на степень изменчивости — это происхождение кости в процессе индивидуального развития. Некоторые кости образуются из хрящевых предшественников и являются частью основы скелета, во многих случаях глубоко интегрированными в составе скелетного модуля, или группы костей, которая развивается в большей или меньшей степени в унисон. У других костей нет никаких хрящевых предшественников. Для тех костей, которые образуются из соединительной ткани, как, например, коленная чашечка в человеческом колене, присутствующая также у многих других видов млекопитающих, ожидается проявление большей изменчивости по размерам и по форме. Коленная чашечка и другие кости, известные как сесамовидные, неплотно сочленяются с другими костями, и подвергаются большему воздействию со стороны мускулов и сухожилий, работающих вокруг области, в которой они формируются. Потому для сесамовидных костей ожидалось, что они будут более изменчивыми, чем другие кости. Эти предсказания были проверены и подтверждены в ходе изучения плейстоценовых млекопитающих, проведённого Кристиной Раймонд и Доном Протеро, которые измерили почти две тысячи костей из двух местонахождений ископаемых остатков: Ранчо Ла Бреа в Лос-Анджелесе и пещеры Сан Хосито в Нуэво Леон, Мексика. Кости принадлежали шести видам млекопитающих: саблезубой кошке Smilodon fatalis, льву ледникового периода Panthera atrox, бизону Bison antiquus, лошади Equus occidentalis, верблюду Camelops hesternus и наземным ленивцам Paramylodon harlani и Nothrotheriops shastensis. Раймонд и Протеро сравнили в рамках статистического подхода изменчивость для нескольких промеров 989 мембранных костей, включая коленную чашечку и другие сесамовидные кости, с промерами для 811 энхондральных костей, прежде всего для таранных костей сопоставимого размера. Для двадцати одного из двадцати семи промеров у мембранных костей изменчивость оказалась значительно выше, чем у энхондральных.Модульность и интеграция
Концепции модульности и интеграции находятся в центре внимания при разговоре об эволюции и индивидуальном развитии. Модули — это подмножество признаков данного организма, которые больше объединены друг с другом, чем с остальными признаками этого организма. Например, бугры коренного зуба гораздо больше влияют друг на друга, чем любой из них влияет на форму клыка, или, естественно, на какие-то другие аспекты анатомии скелета. Сам Дарвин признавал интеграцию между частями организма, и он привёл много примеров корреляций роста в первой главе «Происхождения видов…». Все особенности строения организма интегрированы, но некоторые — в большей степени, чем другие; поэтому интеграция не гомогенна. То, как мы подразделяем анатомию организма, несомненно, связано с этим. Коренной зуб можно было бы осмыслять как единое образование, без его разделения на бугры и другие части, такие, как корень зуба. Если отмежеваться от этого субъективного аспекта, то интуитивно очевидно, что мускулы, части сустава и кости в области колена будет объединены друг с другом более тесно, чем любая из этих частей с любой частью шеи. Эта интеграция не наблюдается непосредственно и должна выводиться из статистического характера изменчивости исследуемых особенностей. Цель состоит в том, чтобы разграничить модули путём количественной оценки характера взаимодействия между чертами строения — так называемого характера ковариации. Подробности того, как это делается, разнообразны и являются предметом споров, поскольку это расширяющаяся область для исследований. Морфологические модули существуют в различных контекстах: развития организма, генетическом, функциональном и эволюционном. Методы исследований, изучающие их, различны в каждом из случаев. Первое всестороннее исследование модульности с использованием количественного подхода было представлено Эвереттом Олсоном и Робертом Миллером в их книге «Morphological Integration», вышедшей в свет в 1958 году. Они определили программу исследований и представили первые попытки статистического подхода к количественному определению корреляции признаков. Олсон и Миллер весьма часто обращались к ископаемым видам, например, упоминая о явно модульной эволюции зубных бугров в родословной линии эоценового млекопитающего Hyopsodus. Они обратили внимание на «прогрессивную утрату у последовательно сменяющих друг друга видов» параконида, одного из небольших бугров в нижних зубах млекопитающих, которая выглядит независимой от других изменений в форме зубов. Более современная работа, проведённая в лаборатории Юкки Йернвалла в Хельсинки, показала, что несколько признаков бугров на коренных зубах млекопитающих в значительной степени интегрировано из-за связи, возникающей между ними в процессе развития. Эти наблюдения имеют большое значение для реконструкции эволюционных древ, которая в случае ископаемых млекопитающих основывается в значительной степени на признаках строения зубов, которые считаются независимыми друг от друга. Некоторые люди активно реализуют исследовательские программы по изучению модульности, и среди них Крис Клингенберг, работающий с очень разнообразными группами позвоночных и беспозвоночных, и Габриэль Марро — с млекопитающими. Марро и его коллеги изучили тысячи черепов некоторых групп млекопитающих, чтобы провести сложные анализы, которые требуют больших выборок материала, чтобы дать статистически существенные результаты. Объединяя свои данные с известными генеалогиями индивидов, которых они изучали, и эволюционными древами в случае целых групп, Марро получил возможность исследовать отношение между генетической структурой популяций и морфологической изменчивостью черепа в них. Главнейшее открытие, сделанное в ходе этого исследования — то, что важнейшим определяющим фактором способности группы к эволюции является её диапазон изменчивости в размерах. Характер интеграции и модулей, обнаруженный у взрослых особей, отражает ассоциации, которые возникают в ходе развития, имеет генетические основы и наследуется. Это означает, что, изучая взрослых особей, мы можем косвенно изучить индивидуальное развитие. Это открывает область исследования для палеонтологии. Анджали Госвами также изучала модульность в черепе млекопитающих, но старалась также охватить исследованием ископаемые формы, чтобы изучить значение модульности для крупномасштабных картин эволюционных процессов. В своём исследовании, на которое часто ссылаются специалисты, она изучила 105 видов, восемь из которых — ископаемые. Она зафиксировала сведения об опознавательных точках многих участков черепа, сопоставимых (гомологичных) для разных видов, таких, как границы между двумя костями. Далее этот трёхмерный массив данных был статистически проанализирован с проведением тестов, которые показали, какие опознавательные точки в значительной степени коррелируют друг с другом и изменяются в унисон. Анджали идентифицировала в черепе шесть модулей: в передней части лица, вокруг коренных зубов, вокруг глазницы, вокруг боковой части черепа, в своде черепа и в основании черепа. Почти все ископаемые формы, которые исследовала Анджали, соответствуют этому стандартному набору модулей, в том числе один саблезубый вид с очень специализированным черепом. Ископаемая саблезубая кошка Smilodon fatalis демонстрирует характер интеграции черепа, отличающийся от такового у любого из остальных исследованных видов: у него лицевой скелет и увеличенные клыки образуют два отдельных модуля вместо одной передне-ротовой — назальной группы. В чём причина такого отклонения? Некоторые аспекты картины роста Smilodon, вероятно, связаны с новым распределением модульности. Для Smilodon характерно уникальное распределение времени смены зубов. Молочные сабельные зубы прорезаются в молодом возрасте, вероятнее всего, позволяя охотиться в то время — и влияя на характер корреляции между частями черепа на ранних этапах его формирования. У других ископаемых саблезубых форм молочные сабельные зубы появлялись поздно, что в итоге вело к более длительному сохранению предкового характера интеграции черепа в эволюционной линии. Эмпирические свидетельства показывают, что модульность сама по себе, как и изменение пропорций между частями черепа в процессе роста (аллометрия), подвержена процессу эволюции. Ископаемые остатки значительно расширяют диапазон морфологически возможных изменений, или же подтверждают закономерности, выявленные в ходе изучения исключительно нынеживущих видов. Поскольку специалисты по палеонтологии позвоночных вынуждены сосредотачивать своё внимание только на скелете, многие другие аспекты анатомии, а вместе с ней и индивидуального развития, которые могут проявлять себя иным образом, выпадают из поля зрения. Из-за явления модульности анатомические области могут вести себя по-разному и независимо друг от друга. Например, у обезьян, группы млекопитающих, к которой принадлежим и мы, скелет в общих чертах сходен у всех видов, тогда как пищеварительная система чрезвычайно изменчива и включает, в том числе, адаптации к ферментации пищи в передней и задней кишке.Выживание здесь и сейчас: утраченные и замещённые модули
Всё в жизни является вопросом альтернатив. Это не наличие универсального оптимума, а локальное достижение лучшего результата. Хороший пример этого «принципа» — самоампутация. Это действие, посредством которого животное отделяет от себя один или больше своих собственных придатков, когда лучше потерять хвост или конечность, модуль, нежели потерять свою жизнь — особенно, если вы сможете затем регенерировать этот придаток, как это известно для многих групп позвоночных, за исключением млекопитающих. Саламандры могут даже регенерировать конечности. Ящерицы, находящиеся ближе к нам на древе жизни, могут регенерировать хвост, но уже не способны регенерировать свои конечности. Хвост — это модуль организма, обычно характеризующийся большей степенью своей изменчивости по сравнению с другими частями осевой области тела у позвоночных, как уже отмечал в 1913 году Э. С. Гудрич. В зависимости от рассматриваемого вида живых организмов, хвосты могут играть важную роль в передвижении, запасании жира или защите от хищников среди прочего. Чтобы вырастить хвост, нужно затратить энергию, и потому, если он есть в наличии, его роль, вне всяких сомнений, принадлежит к числу условий для поддержания жизни. Но много разных животных может выживать и без него. В случае с автотомией хвоста некоторые ящерицы и саламандры, которые схвачены за хвост, будут отбрасывать его часть и таким образом получат возможность спастись бегством. Отброшенный хвост отвлекает внимание хищника от убегающего животного. У сцинков одного из видов (Scincella lateralis) особи, которые отбросили хвост, возвращаются на место, где они его потеряли, и, если они находят его, то заглатывают, чтобы возместить значительную часть затраченной энергии. Ящерицы могут частично регенерировать свой хвост, но при этом исходная структура с позвонками замещается хрящевым тяжом. Внешне хвост также может выглядеть иначе: тоньше или толще, или же иметь окраску, отличную от исходной. Медицинское сообщество, конечно, очень интересуется этим явлением, сознавая важность возможности регенерации тканей для людей, которым была сделана ампутация. В отличие от полной способности к регенерации у амфибий, частичная способность к ней у ящериц — это лучшая модель, демонстрирующая регенерацию ткани в хвосте в противоположность неспособности к регенерации у конечности. Чтобы понять, как действует автотомия хвоста, полезно понять, как она появилась в процессе эволюции. Эволюционировала ли эта способность только один раз, или же много раз и независимо? У каких групп она есть, и что у них есть общего? Эти вопросы всё ещё остаются без ответа. Если рассматривать вымерших животных, то очень трудно, если вообще возможно сказать, какой вид не обладал способностью к регенерации придатков тела, поскольку в данном случае отрицательное свидетельство — это отсутствие свидетельств вообще. Но летопись окаменелостей представляет свидетельства наличия автотомии хвоста. Об этом можно сделать вывод по ископаемым остаткам, на которых сохранилась особого рода пластинка, служащая для разрыва хвоста. На ранних стадиях развития исходные эмбриональные сегменты, называемые сомитами, подвергаются ресегментации, при которой задняя половина сомита соединяется с передней половиной следующего за ним сегмента, и так далее. Пластинка для разрыва хвоста возникает на границе структур, образующихся из двух эмбриональных предшественников, которые объединяются на ранней стадии развития. Когда эту особенность смогли идентифицировать в окаменелых хвостовых позвонках, она была отмечена в разнообразных группах, в том числе у пермских капторинид и других базальных групп рептилий, возможно, у морских рептилий, называемых мезозаврами, возможно, у представителя пролацертиформ Tanystropheus[24], у клювоголовых, ящериц и морских крокодилов из юрского периода, и у ящерицы из среднего эоцена Германии. Поскольку в ходе регенерации позвонки хвоста замещаются хрящевым тяжом, до недавнего времени мы могли только гадать, происходила ли регенерация у ископаемых таксонов. Хельмут Тишлигер из Баварии, который на протяжении ряда лет специализировался на использовании длинноволнового ультрафиолетового излучения для распознавания образований в окаменелостях, сумел открыть новые детали у окаменелостей, известных на протяжении долгого времени, например, отпечатки мягких тканей у берлинского экземпляра Archaeopteryx. Недавно, используя метод фотографирования в длинноволновом ультрафиолетовом свете, который показал ореол, соответствующий былому наличию хвоста, состоящего исключительно из мягких тканей, он обнаружил, что регенерация происходила у некоторых верхнеюрских рептилий отряда чешуйчатых из Германии.Глава третья Онтогенез позвоночных в ископаемом состоянии
Большинство ископаемых остатков позвоночных — это минерализованные части скелета. Поскольку скелет в большинстве случаев лишь частично сформирован у эмбрионов и на других ювенильных стадиях развития, неудивительно, что большинство ископаемых остатков принадлежит взрослым или почти взрослым особям, которые также крупнее, чем другие стадии жизненного цикла, и потому вероятность их обнаружения выше. Но ископаемые остатки эмбрионов и молодых особей существуют. В недавнем обзоре, посвящённом рептилиям, я и мой коллега Массимо Дельфино нашли сотни научных статей, подтверждающих наличие таких образцов (www.developmental-palaeontology.net); многие из них относятся к динозаврам. Некоторые окаменелости интерпретируются как только что выклюнувшиеся из яйца или новорождённые особи, но, учитывая неясности, окружающие эти интерпретации, их лучше считать особями «возраста, близкого к времени рождения», или «перинатальными» особями. Идентификация ископаемого образца в качестве эмбриона неоспорима, когда он найден внутри ископаемого яйца. Более сложный случай — живородящие виды, то есть такие, которые не откладывают яиц, а рождают живое потомство, как это бывает у людей. Разграничить зародыша и последнюю трапезу может быть сложно, когда имеешь дело с ископаемыми остатками возрастом много миллионов лет. Зародыш — это маленький скелет «внутри» более крупного, и этот маленький скелет не несёт признаков повреждений, связанных с пищеварением. Анатомия и размер меньшего скелета, несомненно, имеют основополагающее значение для вывода о том, что он принадлежит к этому же самому виду. Противоречивые интерпретации не так уж и редки. Установление таксономической идентичности эмбрионов и ювенильных стадий — это также сложная задача. В некоторых случаях ассоциации настолько чёткие, что эмбрионы могут быть уверенно отнесены к конкретному виду. В других случаях отнесение образца к той или иной группе основано на некоторых уникальных особенностях, которые являются диагностическими для этой родословной ветви, но более точное таксономическое определение оказывается невозможным. Во многих случаях образцы, вначале описанные как представители различных видов, оказываются с большой степенью вероятности различными стадиями развития одного и того же вида. Редкие ископаемые остатки невзрослых особей — это словно моментальные снимки процесса индивидуального развития, и мы можем многое узнать о росте организма, если будем располагать их серией, или же если нам повезёт обнаружить критическую стадию, например, близкую ко времени рождения, которая сохранилась.Ископаемые птенцы и новорождённые
При изучении истории индивидуальной жизни, относящейся ко времени рождения, перинатальный период и альтернативные стратегии развития — зрелое состояние (выводковое) против незрелого (птенцового) — имеют ключевое значение. Это состояние развития выклюнувшихся из яйца или новорождённых особей коррелирует со многими экологическими и поведенческими параметрами. В случае млекопитающих некоторые новорождённые детёныши весьма незрелые, у них нет шерсти и закрытые глаза — как у хомяков после периода беременности длиной примерно шестнадцать суток. Другие — гораздо более зрелые, рождаются с открытыми глазами, как у морской свинки после беременности, длящейся от шестидесяти до семидесяти суток. У новорождённого детёныша морской свинки и других близкородственных грызунов зубы уже частично сточены из-за движений челюстей, которые начались в утробе. Аналогичные крайности в изменчивости стадий развития при появлении на свет можно наблюдать у птиц.[25] Птенцовые птицы включают пингвинов, чаек, аистов, хищных птиц, попугаев, колибри и певчих птиц, и их птенцы в большой степени зависят от родительской заботы. Выводковые птицы — утки, различные околоводные птицы, тетеревиные, гоацины и индейки. Можно сравнивать разнообразные аспекты анатомии на момент их появления на свет, например, степень дифференциации мозга, или же измерять проявления физиологических процессов. Когда имеешь дело с ископаемыми остатками, степень дифференциации скелета — вот ориентир для выбора. Среди ящериц только что вылупившиеся гекконы обладают значительно менее окостеневшим скелетом, чем большинство других видов, в том числе безногие формы и тегу. Благодаря редким окаменелостям, сохранившим перинатальные стадии развития, палеонтологи получили шанс узнать многое о репродуктивной стратегии и о поведении, связанном с инкубацией яиц. В этом отношении динозавры изучались интенсивнее, чем любая другая группа позвоночных. Один из примеров — это эмбрионы одного из первых когда-либо описанных динозавров. Massospondylus, динозавр из группы прозауропод из ранней юры, получил своё название в 1854 году от Ричарда Оуэна, который ввёл в употребление термин «динозавры» (Dinosauria). Примерно через 120 лет в Южной Африке были найдены яйца этого животного, а через несколько десятилетий, в 2005 году, были описаны их эмбрионы, самые старые когда-либо найденные эмбрионы динозавров. Massospondylus принадлежит к группе, наиболее близкородственной появившимся позже гигантским динозаврам-зауроподам, таким, как Diplodocus. Изучение характера их роста, этих самых крупных животных среди когда-либо бродивших по земле, представляет большой интерес. Крошечные скелеты эмбрионов Massospondylus превосходно сохранились. Почти готовые к выклеву особи не имели зубов, что предполагает, что у них не было возможности питаться самостоятельно. Четыре ноги этих поздних зародышей имели равную длину, что указывает на то, они были четвероногими, хотя взрослые особи, вероятно, были факультативно бипедальными. Череп и глаза были пропорционально более крупными по сравнению с взрослыми особями, что является общим признаком с другими очень молодыми особями позвоночных, хотя черепа молодняка были более высокими и узкими. Основываясь на отсутствии зубов и на других признаках анатомической «незрелости», Роберт Рейсз и его сотрудники выдвинули предположение о том, что потомству после появления на свет мог требоваться уход — это форма поведения, которая также отмечена для других динозавров. Ещё больше об истории жизни Massospondylus известно на основании многочисленных скелетов, представляющих различные стадии роста. Оценивая изменения размеров тела и абсолютное изменение возраста с использованием методов гистологии, мы знаем, что Massospondylus рос со скоростью примерно 35 килограммов в год и всё ещё продолжал расти по достижении примерно пятнадцатилетнего возраста.[26] Также известны прекрасно сохранившиеся эмбрионы в яйцах нескольких других видов динозавров, документирующие тем самым разнообразие репродуктивных стратегий. Среди них — эмбрионы теропод-терезинозавров из верхнего мела Китая. В отличие от прозауропода Massospondylus, молодые тероподы были явно выраженного выводкового типа, или зрелыми на момент выклева из яйца, поскольку им было присуще очень развитое окостенение позвоночного столба, рёбер, тазового пояса и нижних конечностей, в которых две кости, большая и малая берцовые, были уже явственно слитыми и окостеневшими. Применение сканирования методом компьютерной томографии (КТ) сильно облегчает изучение яиц, содержащих эмбрионы. Эта техника позволяет нам заглянуть внутрь яйца, не вскрывая его, не трогая руками. Я и мои студенты используем КТ для разного рода проектов, и это больше не редкость в палеонтологии. Окаменелость аккуратно укладывается на платформу, которая скользит в КТ-сканер, напоминающий гигантский пончик с узкой лежанкой. Обычно объектом исследования являются люди, но окаменелости — более простые объекты: они не шевелятся, не страдают клаустрофобией и не должны задерживать дыхание, пока их сканируют. Прекрасный пример исследований с применением КТ относится к ископаемым остаткам с острова Мадагаскар, принадлежащим недавно вымершей «слоновой птице» Aepyornis, экстраординарному животному, которое достигало высоты около 4 метров и массы тела приблизительно 400 килограммов. Её яйца превышали 7 литров в объёме — они самые крупные среди всех ископаемых или ныне живущих птиц, и крупнее, чем известные яйца зауропод и теропод. Остатки этих животных, в том числе яичные скорлупки, известны из отложений возрастом всего лишь несколько тысяч лет. «Слоновые птицы», возможно, были современниками европейцев, поселившихся на Мадагаскаре в последние несколько веков. У взрослых особей Aepyornis кости черепа срастаются вместе. У эмбрионов на стадиях, которые обычно представлены в виде окаменелостей, можно гораздо легче распознать кости, а также форму и степень их контакта. Это является важным аспектом при сравнении черепов различных групп, ставящем своей целью проследить морфологическую эволюцию. Наиболее заметной особенностью эмбриона эпиорниса является толщина его костей; в сравнении с ним недавно вылупившиеся птенцы страусов и нанду имеют грацильное телосложение. В ходе исследований «слоновой птицы» мои коллеги сумели сложить мешанину костей в реконструкцию эмбрионального черепа и вместе с этим решили вопросы идентификации элементов черепа взрослой особи. Они оценили степень дифференциации скелета у позднего эмбриона: В какой степени минерализуются костные элементы? Существовали ли уже у эмбриона размерные пропорции и пространственные отношения костных элементов, характерные для взрослой особи?Живорождение у ископаемых позвоночных
У позвоночных существует широкое разнообразие репродуктивных стратегий, которые много раз появлялись параллельно в процессе эволюции. Один из примеров этого явления — плацентация, когда существует орган, соединяющий мать с развивающимся эмбрионом и облегчающий получение питательных веществ, газообмен и выведение отходов. Плацента не является исключительным признаком млекопитающих; она достаточно широко распространена среди других основных групп позвоночных, кроме птиц, черепах и крокодилов. Независимая эволюция такой явно уникальной особенности строения, как образование плаценты, также верна для живорождения, стратегии, при которой эмбрион развивается внутри тела матери, а не в яйце. Яйцеживорождение означает, что яйца развиваются и проклёвываются внутри тела матери, также получая защиту и позволяя избегать излишних затрат на воспроизводство. Живорождение несколько раз независимо появлялось в процессе эволюции у большинства групп позвоночных; у одних только ящериц и змей около восьмидесяти раз.[27] Мы и другие плацентарные млекопитающие — живородящие, конечно, как и все сумчатые, группа, которая включает поссумов, кенгуру, коал и их родственников. Но утконос и ехидна, эти ныне живущие представители когда-то гораздо более многочисленной радиации млекопитающих, отдалённо родственных нам, откладывают яйца. Предпосылка для развития живорождения — это механизм, который позволяет оплодотворению произойти внутри тела, где в итоге и будет развиваться эмбрион. Было обнаружено, что самцы некоторых плакодерм, самых первых челюстноротых рыб, живших в палеозойскую эру, обладали придатками, похожими на совокупительные органы и напоминающими подобные образования у современных акул; это даёт основание полагать, что они практиковали внутреннее оплодотворение. Это не только не выглядит такой уж большой неожиданностью, но также подкрепляет тот факт, что древнейшее свидетельство живорождения, датированное поздним девоном, примерно 380 миллионов лет назад, относилось к самке плакодермы, в которой сохранился плод размером в одну треть длины взрослой особи. Важная окаменелость сохранилась настолько хорошо, что демонстрировала то, что должно было быть пуповиной с кровеносными сосудами, как показала компьютерная томография высокого разрешения. Эта и другие экстраординарные окаменелости происходят из формации Гого в Западной Австралии.[28] Одни из самых красивых окаменелостей, чётко документирующих наличие живорождения, известны для разнообразной группы морских рептилий, все представители которой были высоко специализированными к плаванию и населяли моря в мезозойскую эру. Эволюция живорождения у этих водных рептилий была связана с потерей способности откладывать яйца на суше и происходила параллельно значительным морфологическим изменениям, направленным на вторичное приспособление к жизни в воде, подобным тому, что мы наблюдаем у китов и ламантинов, чьи предки уже были живородящими.
Рисунок 13. Рыба Incisoscutum ritchiei, принадлежащая к плакодермам, с придатками, похожими на совокупительные органы (указаны стрелкой) (Ahlberg и др. 2009). Рисунок любезно предоставил Джон А. Лонг (Лос-Анджелес).
Ихтиозавры принадлежат к числу наиболее высокоспециализированных среди всех групп мезозойских морских рептилий и внешне напоминали дельфинов. Мозазавриды также были морскими рептилиями, и были, вероятно, родственны змеям.[29] Эти группы вымерли в меловой период, в конце которого, примерно 65 миллионов лет назад, также вымерли все динозавры, кроме птиц. Живорождение, несомненно, имело место у ихтиозавров и мозазавров; у них это непосредственно демонстрируется присутствием останков эмбрионов в пределах брюшной полости матери. Ориентация эмбрионов у ихтиозавров и мозазавров показывает, что они рождались хвостом вперёд, а голова появлялась самой последней, как это происходит у ныне живущих полностью водных млекопитающих — китов и ламантинов. Эмбрионы ихтиозавров зарегистрированы для видов, живших на протяжении всех эпох мезозойской эры: триаса, юры и мела. Другой разнообразной группой ископаемых рептилий, включающей преимущественно водные формы, были зауроптеригии, к которым принадлежали наиболее заметные длинношеие и крупные плезиозавры. Свидетельства в пользу живорождения в этой группе главным образом косвенные, основанные на морфологии их скелета.[30] В частности, заслуживает подробного рассмотрения форма тазового пояса, поскольку она даёт нам свидетельство ширины родовых путей. У зауроптеригий, так же, как у ихтиозавров и мозазавров, отсутствует жёсткое соединение между костями бёдер и позвоночным столбом. Наиболее вероятно, что это приспособление к плаванию, поскольку конструкция такого плана позволяет делать быстрые остановки или резкие повороты. Предполагается, что такого рода свободное соединение также позволяет этим животным увеличить до максимальной степени пространство для родовых путей.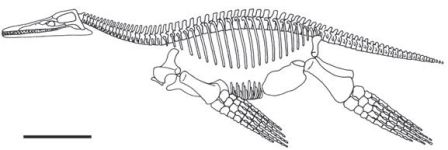
Рисунок 14. Плиозавр Peloneustes philarchus из верхней юры Англии, демонстрирующий слабую связь между костями таза и позвоночником. Из Andrews, 1913 и Sander, 1994, с изменениями.
Физиологические требования к яйцу, находящемуся внутри матки вместо того, чтобы быть вне её, конечно, совершенно иные. Получение кислорода, необходимого для роста, при нахождении в жидкости внутри матки и на воздухе вне её — вот главное отличие. Может показаться, что эволюция живорождения — это трудный или маловероятный эволюционный шаг. Но эта репродуктивная стратегия не только много раз независимо появлялась в процессе эволюции: для неё известны также случаи эволюционной реверсии. Молекулярная филогения удавов позволяет полагать, что в ходе эволюции арабского песчаного удавчика Eryx jayakari происходила эволюция от живорождения к яйцерождению. У этого вида также отсутствует яйцевой зуб, который другие яйцеродящие змеи используют, чтобы выбраться из яйца — дополнительное свидетельство того, что способность откладывать яйца была утрачена и приобретена вновь. Ископаемые свидетельства этого изменения ещё не известны. Несмотря на повторяющуюся эволюцию живорождения среди ископаемых форм, большинство из немногих групп современных морских рептилий представлено яйцекладущими формами. Ныне живущие морские или частично морские рептилии — это морская игуана Галапагосских островов, морские черепахи, шестьдесят два вида морских змей и пингвины (птицы — это рептилии!). Галапагосские игуаны — это замечательные ящерицы, которые щиплют морские водоросли на дне холодного моря, но в остальном они преимущественно наземные и яйцекладущие. (Многие люди видели документальные фильмы, где показаны самки морских черепах, использующие свои ласты, чтобы выбраться на пляж, отложить там яйца, засыпать их песком и вернуться в море.) С другой стороны, большинство морских змей — живородящие виды. Ещё не было найдено ни одного свидетельства того, что живорождение появилось у динозавров, и, хотя этот способ воспроизводства нельзя исключать для этой группы, его появление у них выглядит маловероятным. Ни среди существующих в наше время динозавров (птиц), ни в самой близкородственной динозаврам ныне живущей группе рептилий (крокодилы) нет ни одного живородящего представителя; нет их и в соответствующей летописи окаменелостей. Этот факт представляет собой большую загадку для эволюционных биологов, но, похоже, это связано с ролью яичной скорлупы в обеспечении развивающегося эмбриона важными минеральными веществами. Загадочное отсутствие явления живорождения среди приблизительно 9300 современных видов птиц и всех их ископаемых форм, известных к настоящему времени, нельзя объяснить недостатком геологического времени, в течение которого оно могло эволюционировать, поскольку долгая эволюционная история птиц восходит к юрскому периоду, когда жили Archaeopteryx и другие специализированные динозавры-тероподы. При объяснении этого отсутствия живорождения ссылаются на морфологические или физиологические факторы, явно несовместимые с воспроизводством путём живорождения. Например, птицы обладают высокоспециализированными лёгкими с мелкими капиллярами, которым требуется много времени, чтобы наполниться воздухом после того, как они были заполнены амниотической жидкостью яйца в ходе индивидуального развития. По этой причине большинство птичьих яиц имеет воздушную камеру, в которую поздний эмбрион просовывает свой клюв, чтобы начать заполнять лёгкие воздухом. A стратегия, основанная на живорождении, сразу поставила бы выклюнувшуюся из яйца птицу перед внезапной необходимостью наличия воздуха в лёгких — физиологическая проблема, явно не соответствующая анатомии выклюнувшегося птенца.
Рисунок 15. Дышащий птичий эмбрион незадолго до выклева из яйца. Рисунок Клаудии Джоэл.
Сохранение всеми без исключения птицами яйцерождения должно отражать принципиальные особенности птиц, препятствующие тому, что происходило так много раз у столь многих других групп. Такие предположительные факторы включают полёт, яйцо с изолирующей крепкой скорлупой, которая предотвращает потери воды, особенности определения пола, иммунологические препятствия и особый характер развития лёгких. У каждого из этих факторов есть свои «за» и «против».[31] Фактор полёта в первом приближении выглядит наиболее значимым. Какой матери хотелось бы таскать детёныша во время полёта? Но ни в одной из многочисленных и разнообразных групп нелетающих птиц никогда не эволюционировало живорождение, а беременные летучие мыши могут летать. Возможно, ни одна особенность птиц, взятая отдельно, не является несовместимой с живорождением или с предшествующим ему состоянием в ходе его эволюции — задержкой яйца в организме. Возможно, имеется целый коктейль из особенностей, которые несовместимы с этим. Они должны включать эндотермию, инкубацию яиц и родительскую заботу, выделение мочевой кислоты в качестве главного компонента азотного обмена, изначально птенцовый тип потомства и известковую скорлупу яйца. Биологи математически смоделировали затраты на задержку яйца в организме, которые включают сниженную частоту появления потомства, повышенную материнскую смертность и сниженный отцовский вклад в размножение, и сделали заключение о том, что они перевешивают потенциальные выгоды для большинства птиц. Возможно, некоторые из этих факторов также объясняют отсутствие живорождения у динозавров, черепах и крокодилов.[32] Вымершие предки птиц, как известно, откладывали яйца. Отдельные окаменелости не только подтверждают наличие яиц, но также дают указания на наличие насиживания, связанное с ними.Яйца, родительская забота и насиживание
Изучение ископаемой яичной скорлупы выявило различные стратегии в устройстве скорлупы, призванные обеспечить безопасность, сохранение жидкости и газообмен с окружающей средой.[33] Отличительные особенности характеризуют несколько групп, и на основании гистологических типов скорлупы сложилась номенклатурная система классификации, или паратаксономия, яиц и яичных скорлупок. Наличие эмбрионов в ископаемых яйцах открыло путь для хотя бы частичного соотнесения паратаксономии яиц с таксономией, основанной на скелетных особенностях. Соотнесение яиц с конкретным видом также можно осуществить на основании ассоциации скелета и яиц, предоставляющей нам свидетельство насиживания или разорения кладки. Классический пример путаницы между этими двумя вариантами — случай с Oviraptor, динозавром из мелового периода Монголии. Ископаемые остатки были впервые найдены во время знаменитой экспедиции в пустыню Гоби, организованной Американским музеем естественной истории в Нью-Йорке в 1920-х гг., и о которой можно узнать из книг Шона Кэрролла «Into the Jungle» и Майка Новачека «Dinosaurs of the Flaming Cliffs». В 1923 году был найден скелет беззубого динозавра поверх кладки разбитых яиц. В дальнейшем Генри Фейерфилд Осборн, в то время бывший главой Музея в Нью-Йорке, присвоил новому животному говорящее само за себя название Oviraptor philoceratops: «ovi» означает «яйцо», «raptor» — «разбойник», а «philoceratops» — «любящий рогатых динозавров». Находку черепа, лежащего прямо на гнезде, интерпретировали как знак преступника, застигнутого на месте преступления песчаной бурей и превратившегося в ископаемое как раз во время кражи яиц из гнезда. Благодаря новым исследованиям, которые провели Марк Норрелл и его коллеги из этого же самого учреждения в последние два десятилетия, мы теперь знаем, что совместные находки скелетов взрослых особей Oviraptor и кладок яиц не означают, что они грабили их; вместо этого они откладывали их и исполняли родительские обязанности. Один из видов родительской заботы — это высиживание яиц, при котором тело родительской особи находится в прямом контакте с яйцами на протяжении долгих периодов времени. Классический пример — это знакомая картина птицы-родительницы, сидящей на яйцах: тело поверх гнезда, задние конечности подогнуты под него, живот соприкасается с яйцами на большой площади, а передние конечности сложены вдоль боков. Есть чёткие свидетельства того, что некоторые динозавры инкубировали яйца подобно птицам. Когда родитель оставался при гнезде или яйцах, он, вероятнее всего, активно защищал их от сородичей или от хищников. Мы знаем, что Oviraptor, как и многие другие виды динозавров, заботился о своих яйцах. Насиживание способствует инкубации, повышая температуру яиц.
Рисунок 16. Сочленённый частичный посткраниальный скелет динозавра-овирапторида из позднего мела Монголии, сохранившийся поверх гнезда. Из Clark, Norell и Chiappe, 1999, с изменениями. Изображение любезно предоставил Американский музей естественной истории.
Среди ныне живущих рептилий насиживание яиц в гнезде известно только для змей семейства ложноногих и широко распространено у питонов. Многие змеи, если они находятся у гнезда, ищут пищу. Прямое свидетельство этого известно из мела западной Индии, где останки почти полного скелета змеи Sanajeh indicus были обнаружены обвившимися вокруг недавно проклюнувшегося яйца в гнезде динозавра-зауропода. Другие особи змей, ассоциированные с кладками яиц на этом же местонахождении ископаемых остатков, поддерживают догадку о том, что происходило их кормление молодыми динозаврами. Такие ископаемые остатки образовались при весьма особенных обстоятельствах. Очевидно, в данном случае образование отложений и захоронение в мелкозернистом осадке были необычно быстрыми, вероятно, ускорившись во время бури. Существуют также другие виды инкубации, кроме яиц в гнезде. Среди позвоночных множество разнообразных примеров демонстрируют рыбы и лягушки. Инкубация у рыб и амфибий относится к таким случаям, когда эмбрионы удерживаются где-нибудь на теле или внутри него, но не в яйцеводах. Существует великое разнообразие местоположений яиц, в которых происходит их инкубация: в желудке, в выводковых сумках на спине, в голосовых мешках, в сумках в паху, или же во рту.[34] Темноспондил Trimerorhachis, обычный в пермских отложениях Техаса, вероятнее всего, демонстрирует пример особого вида инкубации среди ископаемых наземных позвоночных. В области глоточных мешков взрослого животного были обнаружены остатки очень мелких особей, что даёт возможность предполагать, что животное было занято инкубацией в тот момент, когда умерло и затем сохранилось в ископаемом состоянии. Альтернативная интерпретация находки — это каннибализм по отношению к собственной молоди. Но кажется более вероятным, что этот хищник с острыми зубами и мощными челюстями питался рыбой и другими позвоночными с помощью своей пасти, а не молодью своего собственного вида с помощью жабр.[35]Ископаемые эмбрионы млекопитающих
В противоположность знаменитым случаям находок эмбрионов динозавров и других рептилий в летописи окаменелостей, ископаемые эмбрионы млекопитающих являются очень редкими, спорными, или представляют собой главным образом лишь курьёз из-за плохой сохранности. Эмбрионы редки даже в местонахождениях, известных благодаря исключительной сохранности ископаемых взрослых млекопитающих. Большинство млекопитающих обладает репродуктивной стратегией, при которой они рождают слаборазвитых, незрелых детёнышей, что делает маловероятным обнаружение эмбрионов. Чаще встречаются ископаемые остатки зародышей млекопитающих тех видов, которые выработали «выводковый» тип появления потомства: более долгая беременность, меньший размер выводка и больше масса тела взрослой особи. Вероятно, мы никогда не узнаем продолжительность беременности у ископаемых видов, но во многих случаях мы сможем узнать размер выводка. Лошадиные эмбрионы из эоценовых местонахождений Германии показывают, что самки вынашивали только один плод, показывая, что ранние лошади в процессе эволюции выработали стратегию рождения немногочисленного потомства, вероятно, сочетающегося с интенсивной родительской заботой. Это типичная стратегия для стадных копытных, и она соотносится с гипотезой о том, что эоценовые лошади жили стадами, основанной на высокой частоте встречаемости ископаемых остатков в некоторых местонахождениях. В одном случае образование из мягких тканей, окружающее зародыш, было с уверенностью интерпретировано как плацента.[36] Небольшой размер выводка также был зарегистрирован для других хорошо сохранившихся ископаемых млекопитающих. Скелет ореодонта Merycoidodon culbertsoni из олигоцена Южной Дакоты, вымершего родственника парнокопытных (Cetartiodactyla), демонстрирует наличие в тазовой области двух хрупких, но довольно полных черепов, которые были описаны как «близнецы» в матке. Более молодые ископаемые эмбрионы млекопитающих известны для некоторых вымерших гигантских наземных ленивцев из плейстоцена Южной Америки, и для мамонтов из Сибири. Их не полностью сформированные кости демонстрируют анатомические детали, которые решают вопросы, связанные с идентификацией костей, которые во взрослом состоянии слишком изменились по сравнению с костями других видов. В некоторых случаях черепа взрослых особей из-за слияния отдельных элементов или изменения пропорций несут неожиданные особенности, если их сравнивать с другими родственными формами, или у них в анатомии отсутствует ожидаемый элемент. Например, на первый взгляд, на черепе взрослого человека отсутствует предчелюстная кость. Но, как обнаружил в девятнадцатом веке Гёте, исчезновение шва — вот причина, которая объясняет кажущееся отсутствие кости, которая фактически присутствует в нашем черепе.Эмбрион или последняя трапеза?
Редкая возможность взглянуть на шаги эволюции истории жизни у весьма особенной разновидности млекопитающих стала реальностью благодаря открытию Maiacetus inuus, раннего кита из Пакистана возрастом 47,5 миллионов лет. Был найден ископаемый образец с меньшей особью внутри его полости тела. Авторы исследования интерпретировали меньший экземпляр как расположенный в положении in vivo внутри матки взрослой беременной самки, собирающейся рожать. Положение головой вперёд при родах у предполагаемого зародыша похоже на положение зародыша у наземных млекопитающих и показывает, что эти животные, в отличие от современных китов, рождали потомство на суше. Хорошо развитый набор зубов у зародыша был истолкован как признак относительной зрелости при рождении, предполагающий, что новорождённые особи у Maiacetus были, вероятно, активными сразу после рождения. Как и у других предковых китообразных, у Maiacetus было четыре ноги, видоизменённых для гребущего плавания, но способных, вероятно, выдерживать его вес на суше, с похожими на плавники лапами. Но некоторые коллеги с богатым опытом в области анатомии китообразных привели доводы против истолкования находки как плода Maiacetus: положение меньшей особи относительно позвоночного столба старшей особи слишком близко голове предполагаемой матери, на уровне желудка, и отсутствие позвонков её хвоста больше соответствует интерпретации меньшей особи в качестве добычи. Но, каким бы ни был результат этой дискуссии, этот пример показывает важность исследования судьбы трупов умерших животных, когда они разлагаются и начинают процесс сохранения в ископаемом состоянии. Наука, посвящённая такого рода исследованиям, тафономия, становится значимой при исследовании ископаемых эмбрионов и других ключевых окаменелостей, сохраняющих мягкие ткани.Сгнившие и окаменевшие эмбрионы
Благодаря исследованиям в области тафономии мы достигли понимания того, как лучше интерпретировать анатомические детали важных окаменелостей. Исследуя характер посмертного разложения, мы знаем, как анатомические составляющие, отличающиеся друг от друга по своим химическим свойствам, теряются на разных стадиях разложения. Также могут возникать топографические изменения структур организма, которые ещё более затрудняют интерпретацию на первый взгляд полных окаменелостей. Примечательный пример такого рода исследований касался темпа и последовательности разложения отдельных деталей строения у видов базальных позвоночных. Задачей Роберта Сансома, Марка Пурнелла и их коллег в Лестере была интерпретация анатомии предположительных ранних ископаемых хордовых путём исследования их ныне живущих аналогов после смерти. В процессе разложения детали анатомии, образованные мягкими тканями, имеют тенденцию к разрушению, но не все особенности исчезают в одинаковом темпе. Хотя эта работа не касалась эмбрионов или особенностей онтогенеза, она упоминается здесь, поскольку иллюстрирует тафономический подход, а также имеет отношение к приобретению первыми, вымершими представителями родословной ветви особенностей строения, чем-то способствующих пониманию происхождения систем развития, которые обеспечивают появление этих особенностей. Сансон и его коллеги изучали разложение только что умерших особей ланцетника Branchiostoma lanceolatum и миноги Lampetra fluviatilis. Среди деталей, важных для изучения у ископаемых остатков, были глаза, хвост и предшественник позвоночника, известный как нотохорд. Органы были помещены для гниения внутрь прозрачных пластиковых коробок, заполненных солёной водой и инкубируемых при 25 градусах Цельсия сроком до двухсот дней. Исследователи зафиксировали процесс распада для каждой анатомической детали и подсчитали изменения в соответствии с простой системой оценки. Деталями строения, более стойкими к разложению были те, которые являлись общими особенностями всех позвоночных, например, нотохорд. Напротив, детали строения, эволюционировавшие позднее, сгнили первыми. Это прискорбно, поскольку позже возникшие детали строения, которые различают близкородственных животных внутри родословной ветви, являются наиболее информативными для целей реконструкции эволюционного древа. Исследование показало, что, будь окаменелость остатками примитивного хордового или разложившегося позвоночного, по отношению к ней требуются тщательные анатомические и химические исследования. Некоторые из кембрийских ископаемых форм, которых касается это тафономическое исследование, при более тщательном изучении могут оказаться более близкими родственниками позвоночным, чем считалось до этого. Вероятно, мы этого никогда не сможем узнать, если принимать во внимание превратности отклонений тафономического характера. Отложения во всём мире полны окаменелостей, которые ещё ждут того момента, когда их обнаружат, равно как и сотни кабинетов в музеях, и эти окаменелости не представляют собой такой сложности для интерпретации, как эти кембрийские формы. Другой пример тафономических исследований, на сей раз более подходящий к теме этой книги, касается процесса разложения и тафономии крошечных эмбрионов. Эти исследования были инициированы открытием в последние двадцать лет кембрийских эмбрионов многоклеточных животных из Китая. Эти окаменелости по расположению клеток и форме похожи на эмбрионов современных морских животных, но они очень мелких размеров и сложны для изучения. Поэтому даже их органическая природа может оказаться под вопросом и в некоторых случаях действительно подвергалась серьёзным сомнениям. Наиболее информативные исследования стадий развития кембрийских эмбрионов от дробления до стадий, непосредственно предшествующих выклеву, включали получение изображений с помощью синхротрона, крупное технологическое достижение, которое позволяет проводить исследование крохотных окаменелостей на клеточном уровне.[37] Порода подвергается химическому воздействию в целях растворения карбоната кальция, окружающего крошечные окаменелости, состоящие из фосфата кальция. Поскольку каждая из этих окаменелостей составляет менее полумиллиметра в длину, это является трудоёмкой задачей. Если принимать во внимание эти ограничения и тот факт, что для поиска таких крохотных, клеточных окаменелостей было сделано ещё очень мало, таксономический и филогенетический охват ископаемых эмбрионов очень ограничен. Фактически, в большинстве случаев вид животного, представленного различными сохранившимися стадиями, остаётся нерешённым вопросом. Некоторые тщательные исследования раскрыли характер дробления у некоторых эмбрионов — это картина деления клеток, ведущего к образованию скопления клеток на первых стадиях развития. Но некоторые из особенностей трудны для распознания с точки зрения как биологии, так и геологии. Тафономические исследования предоставили нам свидетельства того, какие эмбриологические стадии могут сохраниться и как долго и в каких условиях жили эти, главным образом кембрийские, организмы. Пионерными в этой области были исследования Элизабет Рафф и её коллег, которые провели эксперименты с эмбрионами и личинками морских ежей, группы организмов, предпочитаемых у специалистов в области биологии развития из-за того, что о ранних стадиях их онтогенеза известно многое. Рафф и коллеги обнаружили, что сохранение эмбрионов в основном не связано с размерами, и что минерализация тонких особенностей строения, которые могли бы затем быть изученными, была осуществима во многих обстоятельствах. Одни химические условия больше способствовали точному сохранению клеточной анатомии, чем другие. Некоторые стадии обладали незначительным потенциалом для сохранения. Подобные эксперименты в настоящее время проведены с различными видами, но ещё больше должно быть сделано, прежде чем будут разработаны точные или общие протоколы для интерпретации крошечных, сложных органических структур из отдалённого прошлого. При рассмотрении разнообразия в области индивидуального развития и таксономии, которое должно было существовать, и превратностей геологического сохраненияостатков применение универсального метода может быть очень сложным. Но, по крайней мере, экспериментальный подход с использованием ныне живущих видов скажет нам и тем самым позволит нам более критически относиться к тому, чего мы можем достигнуть, обладая новыми инструментами отображения и лучшим пониманием процесса фоссилизации.Глава четвёртая Кости и зубы под микроскопом
Может показаться весьма невероятным, но самым важным рабочим инструментом для большинства палеонтологов, помимо молотка, является микроскоп. Значительная часть людей, изучающих доисторическое биологическое разнообразие, работает в интересах нефтедобывающей промышленности, исследуя мельчайшую пыльцу ископаемых растений или ископаемых фораминифер; последние являются представителями группы одноклеточных организмов, важной для установления стратиграфической корреляции между геологическими подразделениями. Палеонтологам микроскоп позволяет также изучать микроструктуру тканей ископаемых организмов, в особенности кость, которая стала важным предметом исследований, имеющих отношение к изучению индивидуального развития у вымерших таксонов. Изучение живых тканей, или гистология, является обширным полем деятельности, и многое в ней касается идентификации патологий, например, важная диагностическая процедура при исследовании рака. Сравнительные исследования тканей различных организмов, бывших предметом изучения на протяжении столетий, стали возможными благодаря технологическому прогрессу в изготовлении тонких срезов нежных и зачастую мелких комплексных структур различной консистенции и формы. Самые первые работы с тканями на микроскопическом уровне осуществил швейцарец Вильгельм Гис (1831–1904), уроженец Базеля. Он изобрёл микротом, механическое устройство, состоящее из хитроумно расположенного лезвия, которое использовалось для того, чтобы делать тонкие срезы тканей для микроскопического исследования. Благодаря этой новой технологической разработке ему удалось проследить происхождение различных типов тканей животных на эмбриональной стадии. Насколько я знаю, Гис не изучал микроструктуру ископаемых костей; он скорее интересовался эмбрионами и мягкими тканями, например, делая открытия, которые дали ему повод использовать термин «дендриты» для проводящих отростков нервных клеток. Гистологические срезы с использованием преимущественно того же самого метода, который применял Гис примерно 150 лет назад, в настоящее время используются при изучении эмбрионов ныне живущих видов, которые, учитывая их размер, представляют собой очень сложный объект для изучения методом анатомирования.[38] С достаточно давних пор палеонтологи поняли, что тонкие срезы могут показать важные анатомические детали ископаемых остатков. Британский специалист по ископаемым рыбам Уильям Джонсон Соллас (1849–1936), профессор геологии из Оксфорда, и его дочь Игерна стали пионерами в этих исследованиях, когда в 1903 году издали работу о девонской рыбе с неясными родственными связями. Совершённый ими технический прорыв состоял в последовательном стачивании образца, создании рисунков появившихся при этом сечений и переводе их в восковую модель, увеличенную по сравнению с оригиналом. Соллас вначале создал модели ископаемых змеехвосток (офиурид) и граптолитов, группы вымерших беспозвоночных, родственных баланоглоссу, которые были выставлены в Британской научной ассоциации в 1901 году.[39] Соллас сделал модели многих окаменелостей, но похоже, что сохранились лишь немногие из них. Позже метод Солласа был мастерски использован школой шведских специалистов в области сравнительной анатомии. Но изготовление восковых моделей отнимает чрезвычайно много времени. Например, известная восковая модель головы Eusthenopteron, животного, важного звена в переходе между рыбами и наземными позвоночными, отняла пятнадцать лет времени у двух техников на её изготовление. В наше время цифровая камера и правильно подобранные оборудование и программное обеспечение могут сделать ту же самую работу в течение нескольких дней. Это технический прогресс огромного масштаба. Следующий крупный технологический шаг вперёд был представлен в работе 1970 года французскими исследователями Сесиль Поплен и Арманом Ж. де Рикле, которые изобрели метод получения срезов ископаемых образцов с помощью микротома. Они подбирали технику разделения окаменелостей на срезы, которые можно было бы разместить на стекле, как при стандартных гистологических методах. Это были вовсе не тривиальные сложности, поскольку у окаменелостей твёрдая ткань пористая, хрупкая, ломкая, более гетерогенная по составу и менее эластичная при нагрузках, чем кость современных животных. При пропитке ископаемых костей смолами в вакууме и под давлением каждый срез становится более устойчивым. Химический состав ископаемой кости также отличается от её состава у ныне живущих таксонов. Получившиеся в результате срезы сохраняют очертания различных структур, имеющихся в окаменелости. Следовательно, даже если химический состав изменился или исходные компоненты были замещены, границы между ними сохранились. Эта техника использования микротома была первым шагом к замене метода последовательного стачивания, которому приносится в жертву исходная окаменелость. Новейшая техника визуализации внутренней структуры окаменелости не требует получения срезов. С помощью мощной компьютерной томографии с высоким разрешением теперь возможно изучить детали микроанатомии, даже не разрезая окаменелость.Информативные срезы ископаемых костей
Причина того, что гистология ископаемой кости является богатым источником информации, состоит в том, что кость — это живая ткань, которая претерпевает много последовательных процессов резорбции и реконструкции. Этот факт приносит нам пользу после того, как с нами приключается несчастный случай, при котором мы ломаем кость; мы страдаем от этого в пожилом возрасте из-за остеопороза. Для палеонтологов изменение кости в течение жизни означает, что многое из этой записи жизни позвоночного животного можно увидеть. Ключевой момент здесь — умение различать следы первичного роста кости и процессов вторичной перестройки среди отметок, образовавшихся в процессе роста. Самую ненарушенную информацию о росте скелета позвоночного обычно получают из района средней части длинных костей, таких, как в руках (плечевая кость) или в ногах (бедренная кость). Перед изготовлением срезов кости погружаются в синтетическую смолу, затем они шлифуются и полируются, пока не достигают соответствующей толщины, обычно 0,6–0,8 см. Далее эти срезы изучаются под микроскопом при различном освещении и с разными светофильтрами. Обычно предметом исследований палеонтологов являются отдельные обломки костей, поскольку музейные кураторы обычно не любят агрессивного способа исследований, который включает разрезание ископаемых образцов на тонкие кусочки. Это приводит к проблемам, поскольку зачастую нет уверенности в том, что известно точное с позиции анатомии или таксономии место взятия образца. Несмотря на эту трудность, количество новых открытий, сделанных путём исследований тонких срезов окаменелостей, увеличивается экспоненциально. Моё первое знакомство с палеогистологией произошло благодаря моему более молодому коллеге Торстену Шейеру, который работал в моей лаборатории вскоре после моего прибытия в Цюрихский Университет, и благодаря его бывшему научному руководителю работы на соискание степени доктора философии Мартину Сандеру из Бонна, мировому лидеру в этой области. Я посетил лекцию Сандера в Тюбингене, посвящённую зауроподам, крупнейшим среди динозавров, на примере которых он показал, как много может дать палеогистология в понимании характера роста и устройства костей у этих животных. Мне пришло в голову, что этот подход можно было бы использовать для исследования Stupendemys geographicus, самой крупной водной черепахи из когда-либо существовавших, представителя гондванской группы, остатки которой мы собрали в ходе моего полевого проекта в Венесуэле. Сандер направил меня к Шейеру, и мы исследовали Stupendemys и других животных, главным образом млекопитающих и морских рептилий, о чём я и расскажу далее. Панцирь Stupendemys принадлежит к числу самых толстых и самых крупных среди известных панцирей черепах; у одного экземпляра он достигает 3,3 м в длину. С точки зрения гистологии он демонстрирует хорошо выраженную слоистую, словно бутерброд, структуру, где внешние слои, включающие плотную кость, устойчивую к напряжению, окружают ячеистую внутреннюю губчатую кость. Такой тип структуры был важен для уменьшения веса панциря при сохранении прочности в процессе роста. Совокупность отметок, характеризующих рост, была неполной из-за эрозии, которой, несомненно, подверглись образцы, изучавшиеся нами, и которая характеризует местонахождение окаменелостей Урумако в Венесуэле.[40] Однако, вид ткани и просветы между отметками, образовавшимися в процессе роста, сопоставимы с таковыми у ныне живущих черепах с «нормальным» медленным темпом роста. Если предположить темп роста подобным таковому у большинства современных морских черепах с сопоставимыми гистологическими характеристиками, то экземпляр, который мы изучили, должно быть, рос примерно от 60 до 110 лет, прежде чем достиг гигантского размера панциря. Организация ткани, наблюдаемая у современных животных, служит ключом для понимания особенностей гистологии, которые можно наблюдать в ископаемых костях. Остеобласты — это клетки, создающие кость. Также существуют пустоты, называемые лакунами, которые связаны друг с другом посредством канальцев, ответственных за распределение питательных веществ и кислорода. Эти и другие основные особенности строения кости могут наблюдаться в окаменелостях. Остеобласты и остеоциты, кровеносные сосуды или васкулярные каналы и коллагеновые волокна в большинстве случаев разрушаются в ходе фоссилизации, но их положение и форма могут быть зафиксированы в полной мере.[41] Некоторые исследования ныне живущих позвоночных показали, что костные ткани у животных, которые растут быстро, обладают иными особенностями в сравнении с тканями тех, кто растё медленно. Быстрый рост связан с костной тканью, богатой кровеносными сосудами, означающей усиленное снабжение кости кислородом и питательными веществами. Медленный рост коррелирует с менее васкуляризированной костью, часто прерываемой концентрическими линиями, сходными с годичными кольцами дерева. Это открытие — то, что скорость роста влияет на тип откладываемой костной ткани — было опубликовано Родолфо Амприно из Туринского Университета в 1947 году и сейчас известно как правило Амприно. Изучение гистологии костей было продолжено после выхода статьи Амприно, и палеонтологи следовали ходу его мыслей, пытаясь реконструировать аспекты физиологии и истории жизни ископаемых видов.
Рисунок 17. Тонкий срез фрагмента щитка Stupendemys geographicus из позднемиоценовой формации Урумако, Венесуэла. Образец находится в коллекции Университета Франсиско де Миранда в Коро (UNEFM-CIAPP-2002-01).

Рисунок 18. Схематическое представление двух основных видов гистологии костей. Они обычно отмечаются для эктотермных «рептилий» (слева) и эндотермных птиц и млекопитающих (справа). По Chiappe 2007, с изменениями.
В микроструктуре кости также существуют следы, которые предоставляют нам информацию о продолжительности жизни, возрасте и зрелости; это предмет изучения для скелетохронологии. Оценка возраста основывается на идентификации отметок, возникающих в процессе роста, таких, как кольца, образующиеся при замедлении роста, и линии задержек роста (ЛЗР) между зонами более быстрого роста кости. В большинстве случаев отметка, свидетельствующая об одном замедлении роста (кольцо) или его остановке, откладывается каждый год, во время более холодного, или же более сухого сезона, и это вызвано характером питания и физиологией животного наряду с переменными составляющими вроде болезни и репродуктивной цикличности. Из-за этих событий, происходящих в жизни отдельной особи, мы знаем, что образование маркеров роста может быть нерегулярным. Типичная картина отклонения — наличие короткого периода роста между годичным кольцом и ЛЗР, или когда за один цикл роста откладываются два кольца (плюс одна ЛЗР). Кроме минимального возраста, по микроструктуре костей можно дать оценку половой зрелости. Уменьшение ширины зон быстрого роста и обычно изменение вида откладывающейся костной ткани может подразумевать достижение половой зрелости. В наилучших случаях может быть изучена ростовая серия образцов, и данные, полученные из различных элементов скелета, в противоположность единственному образцу, являются намного лучшим материалом для того, чтобы делать достоверные выводы о характере роста вымерших позвоночных. Для оценки минимального возраста животного в каждой кости подсчитываются циклы роста; для заключения о возрасте половой зрелости анализируется размер годичных слоёв. Когда в разных костях одной и той же особи наблюдается резорбция костной ткани, для оценки минимального возраста индивида используется кость с минимальным количеством резорбции и максимальным количеством циклов роста. Разрушение и перестройка кости осуществляется клетками, которые носят название остеокласты.[42] Это интенсивный и энергоёмкий процесс; поэтому развитие кости тесно связано с метаболизмом. Это представляет непосредственный интерес для людей: самые последние исследования в области молекулярной медицины показали, что ожирение, диабет и остеопороз — это взаимосвязанные состояния.[43] В этом контексте были важны эксперименты с лабораторными мышами. Многое из того, что мы знаем о том, как растут и изменяются кости, получено прежде всего в ходе контрольных исследований, когда изучаются кости отдельно взятых животных, для которых исследователи знают пол, историю жизни, условия жизни и рацион. Например, у многих видов позвоночных беременность может привести к значительной перестройке костных тканей.[44] Это было зарегистрировано для многих видов рептилий, таких, как черепахи и крокодилы. В зависимости от группы источником кальция являются строго определённые кости, но не какие-то другие. Одной из них является малая берцовая кость, более тонкая из двух длинных костей, которая также присутствует в задней конечности и у нас, у людей. У некоторых видов крокодилов остеодермы у откладывающих яйца самок функционируют как хранилища кальция в ходе оогенеза. Эта стратегия, вероятно, характеризовала, по крайней мере, некоторых ископаемых рептилий, поэтому, как правило, использование остеодерм для целей скелетохронологии не является оптимальным решением, потому что в них происходит перестройка костного вещества. В случае с черепахами можно было бы подумать, что панцирь представляет собой превосходный источник материала, но пока изученные виды не демонстрируют использование самками этого источника. Возможно, панцирь просто слишком важен для обеспечения биомеханической целостности животного. Вместо него самки используют длинные кости вроде бедренной. Это явление в ходе оогенеза аналогично тому, что происходит у людей во время беременности и лактации, в периоды, характеризующиеся уменьшением плотности минеральной составляющей костей у женщин. Птицы производят твёрдую скорлупу яиц, состоящую больше чем на 90 процентов из кристаллов карбоната кальция. У птиц перед овуляцией в полости костного мозга образуется особый вид эфемерной костной ткани, служащий источником кальция, который позволяет матери образовывать скорлупу яиц. Этот тип кости называется медуллярной костью, и теперь он описан для Tyrannosaurus rex и для других видов, представляющих основные группы динозавров. Это открытие стало важным не только для того, чтобы вывести происхождение птиц из динозавров, но также (что даже ещё важнее) для более точной реконструкции кривых роста для многих видов динозавров с использованием надёжного маркера состояния репродуктивной зрелости. У динозавров этот момент совпал с переходом от стадии ускорения роста к стадии замедления.Характер роста динозавров и происхождение птиц
Мы очень много знаем об онтогенезе костных тканей у динозавров, и гораздо меньше о нём у видов млекопитающих. Главной проблемой, связанной с палеогистологическими исследованиями, был ответ на вопрос, на кого больше были похожи динозавры в физиологическом плане: на эктотермных рептилий, или на эндотермных птиц и млекопитающих. В 1960-х и 1970-х годах Арман де Рикле и его коллеги в Париже выяснили, что кость динозавра не является типичной рептильной костью, но вместо этого она сильно васкуляризирована, подобно костям птиц и млекопитающих. В целом это дало основание полагать, что динозавры быстро росли, и что рост поддерживался высоким уровнем метаболизма. Эти открытия внесли свой вклад в смену парадигмы динозавров — с медлительных, тупых и холоднокровных гигантов на красочных, динамичных и теплокровных существ. Однако, некоторые исследования показали, что быстрый рост не может просто коррелировать с эндотермией, или теплокровностью, поэтому простого ответа на вопрос относительно эндотермии и эктотермии не существует. Вместо этого у динозавров обнаружена богатая и разнообразная картина уникальных типов роста и физиологии. Существует много видов динозавров, они жили на протяжении большого отрезка времени, приблизительно 160 миллионов лет, и достигали максимального для взрослой особи размера в диапазоне от примерно 1 килограмма до 75 тонн. Некоторые динозавры выжили и в процессе эволюции превратились в птиц{5}, и это ещё один крупный переход. Де Рикле, Жак Кастане, Джек Хорнер и их коллеги по всему миру улучшили качество выборки по различным костям и их сравнению с ныне живущими аналогами, и также разработали скелетохронологический метод, который дал информацию о темпах роста. Кевин Падиан и его коллеги сообщили, что у многих видов динозавров в компактной костной ткани имеется кость различных типов, отражающая изменения в темпе роста; похоже, что в конце жизни рост замедляется или даже прекращается. Мы знаем, что, даже если стратегии роста у динозавров сильно различались, эти животные росли в темпе, существенно опережающем таковой у ныне живущих рептилий, далёких от птиц с позиций систематики. Даже крупнейшие из динозавров достигали взрослого размера меньше, чем за три десятка лет. Длинные кости крупных динозавров и птерозавров в большинстве случаев состоят из хорошо васкуляризированной фиброламеллярной кости, что отражает быстрый темп роста. Этот вид костной ткани обнаруживается у коров, лошадей, лосей и других крупных млекопитающих, а также у крупных птиц. Но ранние птицы и многие динозавры не всегда обладают костью такого типа, и если он у них есть, это не обязательно вся компактная костная ткань. Большой объём палеогистологических исследований был проведён в отношении динозавров-зауропод.[45] Особый интерес к этим животным вполне оправдан, поскольку они были самыми крупными наземными животными. Особенности гистологии костей передней и задней конечности были использованы Мартином Сандером и его студентами в Бонне для изучения темпа метаболизма и поиска ответа на вопрос, достигались ли большие размеры за счёт увеличения темпов роста, или же продления времени роста. Крупные юрские формы, потенциально эндотермные, явно росли быстро — несомненно, быстрее, чем самые ранние виды зауропод. Mamenchisaurus, позднеюрский зауропод из Китая, рос с максимальной средней скоростью 2 тонны в год, и это замечательное достижение для метаболизма. Некоторые виды демонстрируют пластичность в развитии, и действительно, различные онтогенетические «стадии» можно распознать, основываясь на различных видах гистологии костной ткани в пределах одного вида. Если использовать для сравнения только современных рептилий с их замедленным темпом метаболизма, то можно прийти к выводу, что огромному юрскому зауроподу Apatosaurus должно было требоваться до 120 лет, чтобы дорасти до своего полного огромного размера. Но, изучая гистологию костей, мы знаем, что Apatosaurus должен был достигнуть своего полного размера в возрасте всего лишь от десяти до двенадцати лет; эта картина больше сходна с наблюдаемой у эндотермных птиц и млекопитающих. Другие виды динозавров также стали объектом палеогистологических исследований, включая, разумеется, харизматичных теропод — группу, к которой принадлежит не только Tyrannosaurus rex, но также и птицы. Возможность определить, когда и как появился эндотермный тип роста у птиц, представляет большой интерес. У Tyrannosaurus масса тела взрослой особи достигала приблизительно 5 тонн, а максимальная скорость роста-2,1 килограмма в день.[46] По массе и скорости роста T. rex был сходен с современным африканским слоном. T. rex доживал до двадцати восьми лет и достигал скелетной зрелости в возрасте примерно двух десятков лет. Как и у других наземных теропод, но в отличие от современных птиц, половая зрелость у T. rex наступала задолго до того, как животное вырастало до своего полного размера. Как и в случае с гигантскими зауроподами, T. rex достигал своего огромного размера главным образом за счёт ускорения темпа роста по сравнению со своими предками. Уменьшение размера тела при эволюционном переходе от не-птичьих динозавров-теропод к птицам документировано многочисленными и довольно полными окаменелостями. В ходе изучения палеогистологии Кевин Падиан и его коллеги достигли понимания изменений в процессе роста, лежащих в основе этого изменения размеров. Они пришли к заключению, что уменьшение размера, связанное с происхождением птиц, включает сокращение продолжительности фазы быстрого роста предков.Кости моа
Объектами палеогистологических исследований также стали другие рептилии и птицы, особенно те, которые демонстрируют крайне выраженные признаки адаптации. Опираясь на них, мы знаем, что продление времени роста было механизмом, задействованным в эволюции двух очень крупных и впечатляющих форм: гигантского североамериканского крокодиломорфа Deinosuchus из позднего мела Северной Америки и Varanus (Megalania) priscus из плейстоцена Австралии, вдвое превосходившего по размеру своего близкого родственника, комодосского варана. Многие животные, обитающие на островах, проявляют на протяжении истории своей жизни своеобразные адаптации, которые могут быть объяснены при помощи палеогистологических исследований, и среди них моа. Моа были нелетающими травоядными птицами, эндемичными для Новой Зеландии. Среди одиннадцати их видов из шести родов два самых крупных вида, Dinornis robustus и D. novaezealandiae, достигали почти 4 метров в высоту с вытянутой шеей и весили примерно 230 килограммов. Моа принадлежат к бескилевым птицам, которые включают африканского страуса, австралийского эму, новозеландского киви и южноамериканского нанду. Вероятнее всего, в результате охоты маори моа, как полагают, вымерли к 1500 году н. э. Моа демонстрируют явно выраженную «K-стратегию», которая означает долгую беременность и немногочисленное потомство — что сопровождается длительным периодом роста и поздним наступлением половой зрелости — в противоположность другим группам птиц, включая других бескилевых. В ходе изучения палеогистологии нескольких длинных костей, Сэмюэль Тёрвей с коллегами зарегистрировали у четырёх из шести родов моа несколько ежегодных задержек роста, или линий задержек роста, обнаружив, что у этих особей типичный размер взрослой особи достигался лишь по истечении многих лет прерывистого роста. Но у гигантских форм моа, у которых самки весили более 200 килограммов, а самцы до 85 килограммов, наблюдаются особенности гистологии костей, отличные от характерных для других, более мелких видов. У гигантских форм рост был ускоренным: наружные слои кости богаты кровеносными сосудами и демонстрируют лишь немногие ЛЗР, если они вообще есть. Виды Dinornis вырастали за промежуток времени около трёх лет, при том, что самые мелкие виды моа, вроде видов рода Euryapteryx, не достигали взрослого размера до девяти лет. Медленный рост и позднее наступление половозрелости появились в процессе эволюции в среде, свободной от хищников и, возможно, с ограниченными ресурсами и сниженной конкуренцией, хотя это лишь предположение. Прибытие людей, как и в других похожих и печальных случаях, вскоре привело к исчезновению этих животных, жизненная история которых была неподходящей для сопротивления интенсивному хищничеству. В настоящее время в Новой Зеландии какапо, единственный в мире нелетающий попугай, возможно, тоже обречён — в результате разрушения среды обитания и прочих форм давления со стороны человека.[47]Развитие костей и локомоция у ископаемых видов и у галапагосских игуан
Каждое утро по пути к своему офису я имею удовольствие проходить мимо наших музейных экспозиций, в которых выставлены серии образцов, демонстрирующих рост, или полные скелеты взрослых особей ихтиозавров, плакодонтов, пахиплеврозавров и плезиозавров. Эти окаменелости прекрасны, и, хотя с таксономической точки зрения они были далеки от моей исходной специализации, у меня появился интерес к ним. У нас есть превосходные коллекции триасовых морских рептилий из местонахождения Монте-Сан-Джорджио в Тичино, италоговорящей области в южной Швейцарии.[48] Среди ихтиозавров представлены самые древние свидетельства наличия живорождения в этой группе. Другие морские рептилии принадлежат к родословной ветви, которая показывает различные степени приспособленности к жизни в воде. Плезиозавры, вероятно, были живородящими и полностью ограниченными жизнью в воде, тогда как некоторые плакодонты были, самое большее, полуводными. Пахиплеврозавры из Монте-Сан-Джорджио включают различные виды, которые отличаются друг от друга по своему водному образу жизни. «Пахи» (как их неофициально именуют) выглядят очень изящными. Голова и всё тело у них удлинены, а конечности обычно сохраняются в положении, в котором они, вероятнее всего, находились во время движения, а именно, прижатыми к телу. Вероятнее всего, они питались рыбой и кальмарами. Если основываться на форме и сочленении их костей, то очевидно, что они двигались вперёд с помощью волнообразных движений, и при этом хвост, должно быть, играл важную роль. Микроструктура костей также важна для понимания локомоции у этих животных; вес и балласт должны играть основную роль. Кости у пахиплеврозавров были тяжёлыми, потому что они были плотными — это служило для противодействия подъёмной силе со стороны лёгких и таким образом облегчало ныряние и поиск корма под водой. Фактически, обиходное немецкое название группы «Dickrippensaurier» означает «толсторёберная ящерица», как и название «пахиплеврозавр», которое также может быть переведено как «толстый плеврозавр» или «толстобокая ящерица». Вопрос, который представляет большой интерес — как развивались кости этих животных. Ясмина Хьюджи из моей лаборатории исследовала эту тему, изучая гистологию ростовой серии четырёх видов пахиплеврозавров. Ясмина обнаружила, что после выклева из яйца происходит непрерывная минерализация костей, которой управляют три процесса: 1. Обызвествлённая хрящевая сердцевина никогда не рассасывается полностью, поэтому на любом этапе обнаруживается лишь небольшая медуллярная полость, или же её нет совсем. 2. Отложение кости начинается одновременно в различных местах в пределах хрящевой сердцевины. 3. Наружный слой кости увеличивается в толщину благодаря непрерывному нарастанию при минимальной или совсем не протекающей резорбции внутреннего слоя кости, формирующейся вокруг основного стержня, надкостницы. Как сравнить «пахи» с ныне живущими аналогами? Изучение современных видов всегда важно для интерпретации данных, полученных от окаменелостей. Ныне живущих представителей пахиплеврозавров не существует, но их самые близкие родственники — это группа, к которой принадлежат ящерицы и змеи. Постепенное удлинение тела и редукция конечностей у ископаемых форм много раз происходили в ходе эволюции ящериц. Многочисленные примеры этого можно обнаружить у сцинков (Scincidae), наиболее разнообразной группы ящериц, насчитывающей примерно тысячу двести видов, половина из которых живородящие. Сцинки — наземные животные, но многие из форм, у которых частично или сильно редуцированы конечности — роющие и обитающие в песке животные, где они показывают особый род локомоции, напоминающий плавание. Вот, почему в моей лаборатории мы исследовали развитие скелета различных видов сцинков и сравнивали его с таковым у пахиплеврозавров. Но мы также хотели увидеть, как влияет жизнь в воде на рост костей. Единственная ныне живущая ящерица, которая регулярно плавает и кормится в воде — это галапагосская морская игуана Amblyrhynchus cristatus. Среди близких родственников Amblyrhynchus есть несколько видов наземных игуан, с которыми можно сделать сравнение, в том числе обыкновенный конолоф Conolophus subcristatus. Ясмина Хьюджи исследовала галапагосских и других игуан для своей докторской диссертации в Цюрихе после того, как мы смогли убедить нескольких кураторов музеев по всему миру предоставить нам кости конечностей этих животных, чтобы сделать срезы. Поскольку игуаны, подобно большинству многоклеточных животных, обладают двусторонней симметрией, исследование, сопряжённое с разрушением образца, затрагивает одну сторону, но оставляет другую неповреждённой. Я всегда напоминаю об этом факте музейным кураторам, когда прошу у них разрешение. Для своих исследований мы берём лишь небольшую пробу в диафизах костей конечностей. Кости этих игуан многое рассказали нам об адаптации и эволюции процесса роста. Морская игуана уникальна среди современных ящериц из-за плотности её костей и истории жизни. Полностью наземные ящерицы демонстрируют равновесие между процессами нарастания кости и её резорбции, которое позволяет кости расти, не теряя прочности. У морской игуаны мы зафиксировали замедленный процесс роста, при котором кости приобретают более высокую плотность. Дальнейшее увеличение плотности кости происходит благодаря процессам перестройки, которые заменяют рыхлую волокнистую кость первых лет жизни на более плотную ламеллярную кость. Гистология костей морской игуаны показывает высокие и постоянные темпы нарастания, что отражает относительно высокую скорость метаболизма по сравнению с другими игуанами.
Рисунок 19. Микроструктура длинных костей взрослого самца морской игуаны Amblyrhynchus cristatus (экземпляр из Музея естествознания в Берлине, NKMB 30260). A: Поперечные срезы диафиза лучевой кости, показывающие толстый и неваскуляризированный наружный слой. B: Группа морских игуан, греющихся на берегу (фотография Урсины Коллер). C: Срез лучевой кости при большом увеличении в нормальном проходящем свете. Ежегодно образующиеся линии задержки роста (ЛЗР) отмечены белыми стрелками, а ЛЗР, появлявшиеся на протяжении года, отмечены серыми стрелками. D: Срез при большом увеличении в поляризованном свете. Заметен чётко выраженный узор из светлых и тускнеющих полос, соответствующих циклам роста (тёмные и светло-серые зоны). Изображения любезно предоставила Ясмина Хьюджи.
Скелетохронология, основанная на подсчете маркеров роста и циклов роста между ними, позволяет сделать оценку возраста достижения половой зрелости и минимального возраста на момент смерти. Скелетохронологические данные игуан были соотносимы с информацией об истории жизни, полученной в ходе полевых исследований. Подобно другим крупным игуанам, морская игуана достигает половой зрелости достаточно поздно; самки достигают половой зрелости в возрасте от трёх до пяти лет, тогда как самцы половозрелы в возрасте от шести до восьми лет. Кости и их линии задержек роста показали ожидаемую картину. Соответствие между данными, полученными при изучении от костей, и полевыми данными от экологов, ещё раз убедило нас в верности выводов. Мы обнаружили коренное различие между морской игуаной и пахиплеврозаврами. У игуаны образуется медуллярная полость, а эффект балласта достигается за счёт дополнительных (образованных надкостницей) слоёв окостенения. У пахиплеврозавров, напротив, медуллярная полость отсутствует, а высокая плотность кости дополнительно возникает за счёт минерализованного хрящевого центра. Это сравнение современных животных с ископаемыми — ещё один пример изменчивости механизмов развития, когда они могут быть результатом различной степени адаптации к водным местообитаниям и различных «отправных точек» (различного состояния соответствующих предков) в процессе эволюции.
Рисунок 20. Поперечный срез плечевой кости, показывающий плотность кости и линии роста у взрослого пахиплеврозавра Neusticosaurus pusillus из триаса Монте-Сан-Джорджио, Швейцария (Палентологический институт и музей, Цюрихский Университет, T4178). Фото K. Waskow, в нормальном проходящем свете. Образец показывает шесть линий задержки роста (белые и серые стрелки). Возраст наступления половой зрелости оценивается в три-четыре года из-за резкого уменьшения ширины четвёртого цикла роста (единственная серая стрелка). Две самых наружных ЛЗР расположены близко друг к другу. Медуллярная полость полностью заполнена обызвествлённым хрящом, который подвергся частичной перестройке. Изображение любезно предоставила Ясмина Хьюджи.
Кости, клетки и гены
Это может показаться невероятным, но данные палеогистологии могут давать косвенную информацию о геноме вымерших позвоночных. Это возможно потому, что у ныне живущих видов существует строгая корреляция между размером клетки и размером генома. Размер клетки может быть количественно определён в тонких палеогистологических срезах кости путём измерения полостей остеоцитов, мест, которые при жизни организма занимали клетки, ответственные за формирование кости. Такие оценки были сделаны для десятков видов динозавров, для птерозавров и для некоторых других вымерших пресмыкающихся и палеозойских наземных позвоночных. Достоверность этих оценок ограничена, поскольку просто размер сам по себе — это очень упрощённый аспект характеристики генома, но он, однако, представляет важность при проведении общего сравнения групп живых организмов. Сообщается, что размер генома коррелирует с темпом метаболизма у четвероногих позвоночных, обладателей мускулистых конечностей, обитателей прибрежной полосы или суши. Клетки меньшего размера более эффективны энергетически и позволяют поддерживать более высокую скорость метаболизма, которая, в свою очередь, связана с гомеотермией. В качестве рекомендации стоит более тщательно исследовать эти отношения у ныне живущих родственников изучаемых групп, и следует достичь гораздо большего понимания значения размера генома, прежде чем будет раскрыт полный потенциал палеогистологии в этой области. Соотношение между эволюцией размера генома и морфологическим разнообразием далеко от простой связи, и даже одна только дупликация гена, которая, как считали когда-то, должна тесно коррелировать с морфологическим разнообразием, не имеет прямого отношения к сложности в целом. Генные дупликации — это удвоение части генома, создание добавочных копий уже существующих генов, в результате чего у новых копий возникает потенциальная возможность получения новых функций.[49]Природа зубов
Жорж Кювье похвалился, что он может реконструировать целое млекопитающее только по одному зубу. Это, конечно, преувеличение, но, вероятно, любой палеонтолог-специалист по млекопитающим при наличии выбора предпочел бы взять в качестве источника информации о вымерших видах зуб, а не какую-то другую часть тела. Зубы — это деталь строения позвоночных животных, которая благодаря высокому содержанию минеральных веществ сохраняется в ископаемом состоянии гораздо чаще, чем остальная часть скелета. Эмаль действительно является самым твёрдым веществом в теле млекопитающего. Это происходит потому, что эмаль содержит гидроксиапатит в большем количестве в единицу объёма, чем кость. Ряд серия окаменелостей документирует изменения скелетных тканей в самый ранний момент эволюционной истории позвоночных. Происхождение зубов вызывает споры по нескольким причинам. Одна из них — то, что взгляды на эволюционное древо включённых в этот процесс животных, одних из самых примитивных позвоночных, всё ещё не согласованы, и в результате этого возможны альтернативные реконструкции последовательности событий. Фактически, некоторые авторы, основываясь на ревизии анатомии и родственных связей плакодерм, панцирных челюстноротых рыб, которые были очень разнообразны в девонский период, утверждают, что зубы, вероятно, появлялись в процессе эволюции больше, чем один раз.[50] Но перед решением этого вопроса следует обратиться к ещё более фундаментальному вопросу: чем в действительности является зуб? Не удивительно, что, чем глубже мы проникаем в тайну происхождения позвоночных, тем менее чёткими становятся определения. Так происходит из-за того, что не все особенности, например, местоположение зубов в черепе и связанных тканях, возникли одновременно или в форме, существующей у уже изученных ныне живущих видов. У ранних позвоночных не существует чёткого характера распределения зубных тканей. Конодонты, вероятно, обладали эмалью (этот вопрос ещё не решён окончательно), но у других бесчелюстных позвоночных была ткань, называемая энамелоидом.[51] Энамелоид должен был существовать у последнего общего предка челюстноротых позвоночных, и эта и другие зубные ткани появились и стали более сложными в ходе их эволюции. Другой вопрос заключается в том, связаны ли критерии определения зубов с индивидуальным развитием, или же являются генетическими, или возможны оба варианта; выяснить это можно только по ископаемым остаткам. Мойя Смит и её коллеги сделали важное открытие, касающееся времени и места экспрессии гена sonic hedgehog, известного, помимо всего прочего, как ключевой регулятор индуцирования развития зубов. Исследования ими работы этого гена у кошачьей акулы показали, что, как и у мышей, места экспрессии соотносятся по времени и месту со временем и местом образования зубов и зубных бугров. Факт наличия общего механизма этого процесса у этих отдалённо родственных видов даёт основание полагать, что такой тип гена, отвечающего за развитие, был у последнего общего предка современных челюстноротых позвоночных. Но положение дел тут гораздо сложнее. Как показали Юкка Йернвалл и его сотрудники, зубы являются полигенным образованием, то есть, на форму зуба оказывают воздействие мутации многих генов. Это особенно справедливо для млекопитающих, что иллюстрируют собою мыши, являющиеся в этом отношении классической моделью для исследований и обладающие мутациями более чем тридцати генов, включённых в этот процесс. Генетические механизмы не просты, и между видами должны существовать определённые различия. Эти факторы невозможно изучить непосредственно по окаменелостям. Типичный зуб позвоночного состоит из тканей, которые происходят из различных зародышевых листков или клеток раннего эмбриона.[52] Две ткани являются самыми основными: эмаль, которая происходит из эктодермы, самого наружного слоя клеток эмбриона, и дентин, который образуется из нервного гребня. Клетки нервного гребня образуются у раннего эмбриона, сосредотачиваясь вдоль дорсальной средней линии нервной трубки в виде цепочки, тянущейся в передне-заднем направлении, и оттуда они мигрируют для образования множества видов тканей, в том числе дентина зубов. Развитие зуба начинается с взаимодействия, образования складок и дифференциации двух слоёв в коже, так же, как в случае чешуй, волос и перьев. Некоторые аспекты развития этих структур гомологичны с точки зрения анатомии, поскольку в их основе лежат общие механизмы развития и генетические основы. Эти механизмы должны присутствовать у животного, которое было последним общим предком видов, обладающих зубами и специализированными особенностями покровов тела, в том числе чешуёй, перьями и волосами.Зубы и история жизни
Ископаемые зубы могут дать некоторые подсказки к распределению во времени событий из истории жизни индивидуума. И не случайно зубы являются предпочитаемым объектом при исследовании онтогенеза. Морфология зубных бугров развивается в направлении от кончика к корню, путём своего рода приращения, напоминающего характер роста раковин моллюсков или рогов жвачных (группа парнокопытных, которая включает коров, жираф и оленей). Последовательность прорезывания зубов также коррелирует с событиями в истории жизни, о чём я говорю в главе 8, касаясь наших человеческих предков. Зубы млекопитающих были благодатным объектом для исследований истории жизни. В конце 1960-х годов объектом внимания стало использование слоёв в зубном цементе и дентине для оценки возраста. Эти слои откладываются с регулярной цикличностью, и те из них, которые связаны с долговременными, повторяющимися из года в год сезонами, можно подсчитать, чтобы получить значение абсолютного возраста индивидуумов. Слои в дентине и цементе различимы при стандартном микроскопическом исследовании с использованием простых гистологических методов. Различия в скорости метаболизма, отчасти определяющиеся сезонными изменениями в окружающей среде, вызывают периодичность в отложении слоёв. Зубные маркеры обычно представлены в виде толстых слоёв, отложившихся в течение сезонасравнительно быстрого роста («лето») и сезона ограниченного роста и, следовательно, более тонких полос. Цемент и дентин редко становятся объектом перестроек или резорбции, и по этой причине в силу многих обстоятельств могут быть лучшим скелетохронологическим инструментом, нежели кости, которые подвергаются значительным изменениям и разрушению в процессе роста.[53]
Рисунок 21. Смена зубов (указано стрелкой) у ископаемого носорога Brachypotherium brachypus из среднего миоцена Эрликона, кантон Цюрих, Швейцария. Фотография любезно предоставлена Палеонтологическим музеем Цюрихского Университета.
Существуют многочисленные примеры исследований истории жизни ископаемых млекопитающих; некоторые из этих вымерших видов сосуществовали с людьми в не столь далёком прошлом, как мамонты и большерогие олени. Состоящие из дентина бивни мастодонтов и мамонтов построены из слоистых образований, которые, подобно другим зубам, нарастали путём приращения. Таким образом, их анализ может дать информацию, касающуюся возраста, скорости роста, возраста наступления половой зрелости отдельных животных и характера смертности в популяции. Такого рода работу провёл Дэн Фишер, который также изучал контекст доисторической окружающей среды, используя анализ стабильных изотопов в бивнях. С помощью таких методов можно получить информацию о температуре и рационе в разное время жизни животного. В недавнем исследовании изучение гистологии зубов большерогого оленя Megaloceros (к сожалению, вымершего) установило, что посмертный возраст для нескольких особей из ирландской популяции составлял от восьми до четырнадцати лет. Это похоже на средний посмертный возраст большинства других видов оленей. Крупный размер Megaloceros, особенно его большие рога, а также необычная плотность его черепа дали бы основание предположить для этих животных долгую жизнь, соотносимую с ожидаемым продлением времени роста по сравнению с его предками. Оценки возраста, которые были получены в ходе изучения зубов, противоречат этому предсказанию. И вновь мозаика из скорости и характера эволюции процессов развития разных частей тела животного делает сравнительные исследования истории жизни более сложными и интересными.[54]Износ зубов
Морфология зубов может радикально изменяться на протяжении времени жизни индивида, в некоторых случаях вплоть до исчезновения бугров и обнажения тканей зубов, изначально покрытых эмалью, когда индивид был моложе. Степень износа можно использовать для определения относительного возраста ископаемых организмов, но этот подход имеет множество неудобств, в том числе изменчивость в скорости и характере износа зуба даже в пределах популяции, а также в зависимости от местности.[55] Существует две причины износа зубов: абразивное воздействие, производимое пищей или частицами, которые её сопровождают, и истирание, вызванное контактом между зубами. В случае позвоночных с многократной сменой зубов простая замена — это очевидная стратегия для борьбы с износом.
Рисунок 22. Нижний зубной ряд трёх особей Dremotherium feignouxi, безрогого жвачного из местонахождения Монтегю-ле-Блин (Бассейн Алье, центральная Франция) раннемиоценового возраста. Обратите внимание на значительные изменения формы зубов из-за износа в течение жизни. Фотографию любезно предоставил Луи Костер (Базель).
Но млекопитающие, для которых характерна максимум двукратная смена зубов, в процессе эволюции приобрели альтернативные стратегии. Травоядные особенно часто имеют дело с абразивным воздействием пищи, и долговечность зубов является критически важным моментом для борьбы с их износом. Для этого в процессе эволюции много раз и в различных группах возникали стратегии, суть которых состоит в появлении более высоких и даже постоянно растущих зубов. Изучение характера износа зубов имеет давнюю традицию в палеонтологии млекопитающих, и в ходе исследований затрагиваются также аспекты истории жизни и вопросы функционального плана. В этом отношении были исследованы даже некоторые из самых ранних млекопитающих, такие, как Haldanodon exspectatus из поздней юры Португалии, и было показано, что их рацион проявлял абразивные свойства. Исключительный пример адаптации, призванной компенсировать износ зубов, был зарегистрирован у Hyaenodon, млекопитающего из отряда креодонтов, которое жило на протяжении длительного времени в кайнозое и имело широкое распространение на нескольких континентах. Hyaenodon был плотоядным и обладал режущими зубами, среди которых верхний и нижний хищные зубы должны были тесно контактировать друг с другом, чтобы осуществлять функцию резания. Чтобы сохранить эту тесную подгонку зубов вопреки их износу, происходило поворачивание верхних зубов в медиальном направлении. Это явление было выражено очень явно: верхние зубы у самых старых особей повёрнуты под углом почти в девяносто градусов, а эмалевая коронка полностью стёрта. Эта специализация, очевидно, помогала увеличить среднюю продолжительность жизни Hyaenodon.[56] Некоторые виды ископаемых плотоядных плацентарных и сумчатых млекопитающих также обладают такой специализацией.Глава пятая Пропорции, рости таксономия
Во времена, когда было сделано очень мало анатомических исследований микроскопических структур, многие люди думали, что внутри яиц содержатся полностью сформированные очень маленькие индивидуумы, или гомункулюсы — это теория, известная как преформизм (рисунок 23). Для других видов предполагалось существование аналогичных «анималькулюсов». В наше время эта идея кажется нам смешной, но в то время не было никаких микроскопов, а о наличии эволюционных преобразований организмов ещё не догадывались. Но что действительно замечательно и несёт гораздо меньше смысла без знакомства с современным положением дел в биологии, так это то, как яйцеклетка и сперматозоид могут совместно давать информацию и способность выживать и развиваться целому организму. Сейчас мы знаем, что ранний этап онтогенеза сложных организмов включает простое деление клеток снова и снова, и в конечном счёте происходит развитие в организм узнаваемого облика. Открытие этого процесса и механизмов, лежащих в его основе на различных уровнях организации — это великолепное научное достижение.
Рисунок 23. «Гомункулюс», каким его изображал Николас Хартсокер (1694); эмбрион находится в сперматозоиде уже в готовом виде. Рисунок Мадлен Гейгер.
Есть два аспекта онтогенеза, периода от зачатия до смерти, которые рассматриваются отдельно: развитие и рост. Развитие относится к процессам дифференциации клеток и формирования основного плана строения тела, с фундаментальными структурными изменениями и первым появлением основных деталей строения. Оно начинается с зачатия и завершается приблизительно в момент формированием основных тканей тела. Рост является более поздней стадией онтогенеза, в ходе которой увеличивается размер. В основе этой стадии лежит эмбриональный план строения, который уже сформировался в ходе процесса развития. Из-за связи фоссилизации с процессами сохранения многое из того, что палеонтология может сказать об онтогенезе, относится к процессу роста. Косвенно она может сказать намного больше, о чём я расскажу в следующих главах. Рост как у ископаемых, так и у ныне живущих форм включает изменения размеров и формы. Значительная область исследования связана с выявлением математических законов, которые управляют этими изменениями. Одна из самых ранних попыток сделать это была предпринята многоуважаемым учёным Д’Арси Томпсоном (1860–1948), профессором зоологии в Cент-Эндрюсском Университете в Шотландии. Он был также экспертом в области математики и греческого языка, и именно он, в числе своих прочих заслуг, подготовил стандартный перевод «Истории животных» Аристотеля. Томпсон описал свой подход в книге «О росте и форме», впервые изданной в 1917 году. Он предложил координатные сетки, с помощью которых было возможно перевести сложные различия в формах в простые геометрические понятия. Их предыдущую и более простую версию, касающуюся различий в пропорциях человеческого лица, можно найти в записках Альбрехта Дюрера. Несколько примеров, которые использовал Томпсон в анализе с помощью координатной сетки, относятся к ископаемым формам, среди которых серия родов ископаемых лошадей, Archaeopteryx, вымершие носороги и плечевой пояс молодой и взрослой особей плезиозавра.
Рисунок 24. Плечевой пояс плезиозавра Cryptocleidus в молодом (a) и взрослом (b) возрасте. Из книги Д’Арси Томпсона «О росте и форме» (1917).
Геометрический подход Томпсона заслужил много внимания и похвал, и повлиял на другие области знаний.[57] Но его широкое применение надолго задержалось. Методом стали интенсивно пользоваться лишь в последние десятилетия, когда были разработаны подходящие алгоритмы для работы с комплексной информацией геометрического плана. Начиная с 1980-х годов пережила свой расцвет дисциплина геометрической морфометрии, и были разработаны изощрённые методы работы и компьютерные программы, позволяющие сравнивать разные виды и прослеживать изменения, происходящие в процессе роста; в случае позвоночных большинство из них касалось их черепов. При геометрическом подходе фиксируется много точек-ориентиров и проводится количественная оценка того, как весь их массив различается от вида к виду или между стадиями в серии образцов, иллюстрирующих рост. Существуют различные подходы, основанные на использовании точек-ориентиров; у каждого из них своя собственная математическая структура и они включают методы, основанные на измерении деформации, суперпозиции и линейных расстояний.[58] Альтернативный и более простой математический подход по сравнению с геометрическим подходом Томпсона был разработан примерно в то же самое время. Этот подход также оказал значительное влияние на исследования роста. В 1936 году Джулиан Хаксли и Жорж Тессье опубликовали одновременно на английском и французском языках работу, которая ввела в оборот простое, но изящное уравнение, которое суммирует отношение между двумя измеряемыми величинами. Это отношение было выражено как y = xa или в логарифмической форме, log y = a log x, где a — величина, вычисляемая с помощью отношения. Математический график роста, или онтогенетическая траектория, после логарифмического преобразования обычно представляется в виде прямой линии. Направление или наклон и положение или пересечение с вертикальной осью графика — переменные величины, которые нужно вычислять. Параметры, которые относятся к исследованию роста — это скорость, продолжительность, начало и завершение. Эволюция включает прерывание, изменения и перестановки этих параметров. Рост может порождать изменение формы, и это вызвано изменениями размеров, которые не находятся в отношении 1:1 с формой. Если отношение размеров к форме постоянно, мы имеем дело с изометрией. Это довольно редкий случай, поскольку мы гораздо чаще сталкиваемся с изменением формы, зависящим от изменения размеров, или аллометрией; этот термин Хаксли и Тессье использовали в своей статье 1936 года.[59] Из-за аллометрических отношений пропорций в процессе роста особи одного и того же вида могут иметь различную форму тела на разных стадиях роста. Из этого также следует, что внешний облик видов может меняться, если они обладают различными видами аллометрических отношений в процессе роста, или же если в процессе роста имеет место один и тот же вид отношений, но рост тормозится, или, напротив, продолжается дольше. В онтогенетическом шкалировании различия между двумя видами могут быть целиком отнесены к различиям в размерах, если они попадают на одну общую для обоих видов траекторию роста. Здесь различия являются следствием продления времени роста, или же его остановки. И наоборот, можно сделать вывод об изменениях в характере роста, если изменчивость не вписывается в одну общую траекторию.
Рисунок 25. Классический пример исследования аллометрического роста, призванного помочь понять различные пропорции тела у вида — это исследование вымершего большерогого оленя Megaloceros giganteus, обладателя рогов, которые для нас выглядят огромными. Построив график отношения размеров тела к размерам рогов у разных видов оленей, Гоулд (1974) продемонстрировал, что большерогий олень вписывается в рамки той же самой онтогенетической траектории, что и другие виды. Таким образом, размер рогов этого «гиганта» фактически является ожидаемым для оленя такого размер. Длина рогов соотносится здесь с длиной одной из костей предплечья.
У математического подхода, разработанного Хаксли и Тессье, было несколько предшественников в конце девятнадцатого века, но лишь в двадцатом веке появилось сообщество биологов, готовых освоить и использовать новый числовой инструмент.[60] Сразу же после публикации уравнения роста, которое изящно выражало отношение между двумя размерными величинами, в этом ключе была исследована эволюция вымерших видов. Аллометрический подход был с энтузиазмом принят сообществом палеонтологов, и результатом этого стали многочисленные исследования эволюционных рядов тех таксонов, у которых приобретение какой-то специфической особенности выглядело результатом аллометрического роста — например, рога у ореодонтов (вымерших родственников верблюда), длина ростральной области у лошадей с эоцена до наших дней, гребень на голове у позднемелового динозавра Protoceratops и удлинённые верхние остистые отростки позвонков, образующие «парус» у пеликозавров, очень дальних, ранних родственников млекопитающих из позднего карбона и ранней перми Техаса. По итогам тех исследований было объявлено, что этими особенностями управляют простые взаимосвязи и прямые и постоянные аллометрические соотношения. Проведённые позже ревизии многих из этих исследований показали иное. Когда было обнаружено больше ископаемых остатков и были построены надёжно документированные эволюционные древа для этих групп организмов, стало понятно, что сама аллометрия также эволюционирует, что организмы представляли собой мозаику особенностей строения, растущих с различной скоростью, и что эти изменения находились под управлением экологических факторов.
Рисунок 26. «Пеликозавр» Edaphosaurus boanerges. По Romer и Price, 1940 и Sander, 1994, с изменениями.
Хороший пример такого пересмотра прежних взглядов — эволюция рогов в филогенетическом ряду бронтотериев, вымерших дальних родственников лошадей и тапиров. Наросты на черепе у бронтотериев демонстрируют положительную аллометрию, как и в случаях других групп вымерших позвоночных. Эта разница в росте приводит к изменению формы; отклонения траекторий роста от изометрии ведут к значительным изменениям размеров, что может давать начало новым типам морфологии. Постепенное увеличение рогов с течением геологического времени изначально интерпретировалось как простое продолжение обычной траектории роста. Более новые исследования показали, что в этом процессе были задействованы и другие изменения, такие, как более раннее начало развития и увеличение скорости роста. Эти изменения в траекториях роста могут быть ответом на более массивное телосложение и увеличение усилий, прикладывавшихся к рогам. Классический пример эволюции аллометрических пропорций — это лошади, безрогие родственники бронтотериев. Эволюция черепа у лошадей включала появление новых типов процесса роста, связанных с новым рационом. Они хорошо документированы в виде сложного древа с многочисленными «боковыми ветвями», большинство из которых ведёт к вымершим видам, а другие ведут к видам, близкородственным современной лошади. Единственный современный род Equus — последний уцелевший представитель весьма разнообразной адаптивной радиации, протекавшей на протяжении последних 55 миллионов лет, итогом которой были примерно три дюжины вымерших родов и нескольких сотен вымерших видов. Около 20 миллионов лет назад лошади претерпели взрывоподобный рост разнообразия морфологии зубов. В процессе эволюции у многих родословных ветвей образовались высококоронковые зубы, подходящие для поедания травы на обширных травянистых равнинах большого количества открытых местообитаний, которые распространились в то время. К тому моменту геологического времени оцениваемый размер тела лошади находился в промежутке между ~5 и 50 килограммами. Напротив, в последние 20 миллионов лет лошади были более разнообразными в плане размеров тела. Некоторые группы стали крупными, другие сохранили относительно постоянный размере тела, а третьи со временем стали мельче. Из-за этих отклоняющихся эволюционных путей картина изменений выявленных закономерностей роста в эволюции лошадей выглядит не линейной, а разнообразной и ветвистой.[61] Полагают, что изменения в пропорциях черепа лошади вызваны изменениями аллометрического роста. Эти пропорции у взрослых особей ископаемых видов различны и связаны с функциями: в частности, это изменения в сторону высококоронковых зубов, сильнее сдвинутых вперёд относительно челюстного сустава, и более крупные челюсти для увеличения площади прикрепления мускулов. Аллометрические отношения между изменчивыми характеристиками черепа, наблюдаемые в ходе индивидуального роста ныне живущей лошади не идентичны тем, которые были выявлены путём сравнения черепов взрослых экземпляров различных ископаемых видов. Вне зависимости от конкретной тенденции и от изучаемой группы позвоночных, будь то лошади, бронтотерии, бегемоты, рептилии, амфибии или рыбы, у них у всех имеются некоторые общие особенности роста. Та часть головы, где расположены мозг и органы чувств, в процессе роста демонстрирует отрицательную аллометрию: они меньше увеличиваются в размерах по сравнению с телом по мере роста животного. Морда, часть анатомии, связанная с челюстями, растёт, демонстрируя положительную аллометрию и становясь крупнее по отношению к увеличению размеров тела в процессе роста. Эта общая особенность справедлива также в отношении людей; например, младенцы имеют относительно большие глаза и маленький рот. На уровне популяций в пределах вида аллометрический рост также может приводить к появлению морфологического разнообразия, как среди пород собак. Псовые в целом, группа, к которой принадлежат не только собаки, но также лисы и волки, характеризуются явно выраженным аллометрическим ростом черепа. В противоположность им кошачьи демонстрируют изометрию черепа и, соответственно, породы кошек морфологически менее разнообразны. То, что изменчиво у одной группы, может быть постоянным у другой.Ископаемые ростовые серии и проблемы таксономии
Оптимальная возможность для изучения развития скелета — это серия прижизненных измерений, когда один и тот же индивидуум исследуется на протяжении всей продолжительности его жизни. Если это повторяется на нескольких особях, то мы получаем представление о том, что происходит в процессе роста конкретного вида. Этот вид изучения очень редко применяется к позвоночным из-за очевидных математических и технических сложностей, связанных с этим. И конечно, исследование с помощью серии прижизненных измерений невозможно в палеонтологии. Ростовая серия у ископаемых организмов — это лишь статичная выборка особей одного и того же вида на различных стадиях развития. Их можно признать таковой просто на основании размеров, степени сращения швов в черепе или окостенения некоторых частей скелета. Независимую оценку возраста или онтогенетической стадии ископаемого организма могут дать гистологические данные (см. главу 4). Вполне возможно собрать заслуживающую доверия ростовую серию для ископаемого организма и фактически исчерпывающий размерный ряд для видов, представляющих большинство основных групп ископаемых позвоночных. Но вопрос идентификации онтогенетического ряда ископаемого вида далеко не тривиален. Эту задачу легче всего выполнить, когда доступно большое количество индивидуумов в стратиграфически проверенной области — а это очень редкая ситуация. Неправильные таксономические выводы обусловлены отклонениями, связанными с процессом сохранения и отсутствием внимания к онтогенетической изменчивости. Известно много случаев, когда таксоны, диагностированные на основе молодых особей, позже оказывались младшими синонимами таксонов, описанных раньше на основании образцов взрослых особей.[62] Например, подробное морфометрическое исследование черепов ранних млекопитающих показало, что образцы, ранее отнесённые к четырём видам двух родов, вероятно, представляют собой различные онтогенетические стадии единственного вида. В другом примере измерения образцов различного размера, принадлежащих, возможно, к самому знаменитому таксону ископаемых организмов, Archaeopteryx, были нанесены на график, и было выяснено, что они находятся на одной и той же морфометрической траектории и таким образом относятся к одному виду Archaeopteryx lithographica. Подход такого рода становится рутиной в палеонтологии позвоночных. Другие примеры включают, среди прочих животных, ископаемых ленивцев, капибар и ихтиозавров.
Рисунок 27. Ростовая серия позднедевонской рыбы-плакодермы Bothriolepis canadensis из Мигуаши, Канада. Сохранились головной и грудной щиты; полная выборка образцов насчитывает более 6725 штук, длина самого большого экземпляра справа составляет примерно 20 см. Масштабная линейка-4 см. Рисунок по Cloutier 2010.
Важность принятия во внимание роста при решении вопросов таксономии важно при изучении оленей. Оленьи рога богато представлены в летописи окаменелостей этих животных, потому что они прочные и большие, и люди собирают их. У большинства видов оленей рога драматическим образом изменяются на протяжении жизни. Рога оленей — это действительно превосходный объект для исследований онтогенеза, поскольку они сбрасываются каждый год, а за этим следует стадия роста, ведущая к появлению сложной структуры. Максимальная сложность оленьего рога увеличивается с каждым годом жизни индивидуума. Теперь представьте себе рога в ископаемом состоянии. То, что обычно находят, и что заполняет множество музейных кабинетов по всему миру — это лишь отдельные рога или, чаще, обломки рогов. Для таксономической привязки отдельно найденных ископаемых оленьих рогов требуется тщательное изучение индивидуальных онтогенетических изменений. В прошлом много новых видов было выделено на основании отдельно обнаруженных экземпляров, что привело к завышенной оценке количества видов ископаемых оленей.
Рисунок 28. Внутривидовая (сверху) и межвидовая (внизу) аллометрия. Пример внутривидового явления — прекрасная серия образцов роста ископаемого сумчатого Nimbadon lavarackorum, родственника вомбата (на основе Black и др. 2010). Пример межвидовой аллометрии включает черепа взрослых особей нескольких видов сумчатых из группы Diprotodonta, членов «отряда» млекопитающих с самым большим диапазоном изменения размеров: от примерно 15 г до примерно 3 тонн у вымершего Diprotodon, крупнейшего вида сумчатых, известного из плейстоцена.

Рисунок 29. Очертания рогов северного оленя по достижении ими максимального размера на протяжении пяти последовательных лет. Служащие национального парка Кэйрнгормс в Шотландии дали этой особи имя Уолли. Т. Смит (2006) представил описание интродуцированной популяции северных оленей, к которой принадлежала эта особь.
Для зубов также отмечено изменение размеров и формы на протяжении жизни индивидуумов, и палеонтологи проводят количественную оценку этой изменчивости, когда принимают решения относительно таксонов ископаемых животных. Например, капибара, самый крупный из ныне живущих грызунов, послужила объектом сравнения при оценке изменений среди ископаемых образцов южноамериканских грызунов и при их таксономических исследованиях. Результатом исследований стало выявление синонимии, то есть, невалидности видов, описанных в прошлом. Зубы представляют собой особый случай при исследованиях роста благодаря тому факту, что в ходе жизни они не подвергаются никакой перестройке, однако из-за износа изменение их формы может быть значительным. Исследования гистологии костей тесно связано с изучением серий образцов роста ископаемых форм, поскольку они могут дать независимую оценку относительного, а иногда даже и абсолютного возраста образца. В течение нескольких последних лет стало всё более и более обычной практикой при описании нового вида динозавров представлять также гистологические данные, даже когда в распоряжении есть лишь единственный образец. Такое внимание к онтогенетической изменчивости делает таксономию более основательной и менее типологической практикой, хотя нельзя сказать, что его совсем не было в работе палеонтологов прошлого. Американский палеонтолог девятнадцатого века Отниэль Ч. Марш, например, использовал степень срастания швов в черепах бронтотериев, чтобы понять, представляет собой образец взрослую или молодую особь. Швы, границы между костями, сглаживаются по мере роста животного. Последовательность, в которой это происходит, является видоспецифичной.Глава шестая Характер роста и процессы увеличения многообразия
Самая плодородная почва для распространения (эволюционного) синтеза, вероятно, лежит в развитии теории применительно к роли внутренних биотических факторов в установлении отличных друг от друга скоростей возникновения и вымирания видов, и в понимании того, как эти основополагающие макроэволюционные переменные складываются, давая в итоге собственно процесс увеличения многообразия.Окаменелости обладают потенциальной возможностью давать прямое свидетельство того, как изменения в стратегии роста могли оказывать воздействие на процессы увеличения разнообразия с течением геологического времени. Новые стратегии могли позволить некоторым видам воспользоваться новыми экологическими возможностями, или же вносили свой вклад в их гибель. Характер эволюции групп организмов в геологическом времени, обычно называемый динамикой клады, является предметом интенсивных исследований в том сегменте сообщества палеонтологов, который работает с беспозвоночными животными. Работа в этой области во многих случаях обращается к обширным базам данных и математическим моделям, которые основаны на нескольких переменных величинах. Темпы изменения разнообразия, означающие, насколько быстро или медленно меняется количество видов в группе с течением геологического времени, традиционно связываются с внешними факторами окружающей среды. При объяснении эволюционного события обычно идёт спор о двух альтернативных внешних факторах: Чёрная Королева против Придворного Шута. В сценарии Чёрной Королевы главными направляющими силами изменений являются биотические взаимодействия, а в сценарии Придворного Шута самыми важными являются физические или абиотические возмущения, такие, как изменения климата.[63] «Внутренние» факторы, то есть те, которые связаны с физиологией и развитием организма и являются результатом его эволюционной истории, также стали предметом возрастающего интереса. Исследования множества групп, которые жили в различные геологические эпохи, показали, что противопоставление друг другу внешних и внутренних факторов является ложным. Переменные компоненты истории жизни развиваются, находясь в пределах, обусловленных внутренними особенностями включённых в этот процесс организмов, которые изменяют окружающую среду, но также и подвергаются её воздействию. Крупномасштабный пример такого рода — это появление фотосинтезирующих бактерий, которые с течением геологического времени изменили атмосферу, высвобождая кислород, что документируют минеральные отложения, возраст которых установлен. Появление в атмосфере свободного кислорода также стало пусковым событием на геологическом уровне, поскольку кислород вносит свой вклад в характер химического выветривания горных пород. Некоторые из условий окружающей среды, с которыми должны были сталкиваться на протяжении своей жизни ныне вымершие организмы, крайне сильно отличались от тех, которые окружают нас сегодня. Например, содержание кислорода в атмосфере в различные отдалённые периоды времени было намного ниже, или же намного выше, чем сегодня. Также стоит подумать над значением концепции «глубин времени» для понимания эволюции индивидуального развития в крупных родословных ветвях. Как теоретические ожидания, так и эмпирические данные показали, что для процессов роста биологического многообразия простые корреляции являются исключением. Предрасположенность к видообразованию или к устойчивости к вымиранию не может сводиться случайным образом к единственной особенности; во всех случаях важно рассматривать все переменные величины, относящиеся к истории жизни. Некоторые различия в окружающей среде между нынешним миром и прошлыми эпохами затрагивают переменные величины, которые постепенно изменялись по ходу геологического времени. Другие же связаны с внезапными, катастрофическими событиями такого масштаба, какой никогда не был зафиксирован на протяжении летописной истории человечества. Некоторые из этих событий отмечены во времена, когда происходили массовые вымирания, например, как вымирание динозавров. С ними связан исключительно активный вулканизм. Последствия от удара астероида, который упал на полуострове Юкатан, признаны многими. Возможно, столь же существенным было воздействие событий, которые привели к появлению Деканских траппов на Индийском субконтиненте; это одно из самых крупных проявлений вулканической активности на суше, когда на континентальной коре образовались гигантские плато из лавы. Траппы плоскогорья Декан представляют собой полмиллиона квадратных километров вулканических пород, в десять раз больше, чем территория Швейцарии. Данный ландшафт состоит из нескольких плоских областей, или траппов, каждая из которых была образована поверхностью одного лавового потока и раскинулась на многие километры. Деканские траппы являются последствиями вулканических явлений, масштаб которых не сравним ни с чем из того, что известно нам из истории человечества. Даже извержение Везувия и его хорошо известные последствия для Помпей бледнеют по сравнению с ними.[64] Радиометрическое датирование показало что извержения, которые привели к появлению деканских траппов, охватывают отрезок времени примерно в 2 миллиона лет и случились около 66 миллионов лет назад. Эти вулканические извержения не только залили лавой обширные территории, но также выбросили в атмосферу большое количество двуокиси серы и мелкой пыли. Климатические изменения, ставшие результатом этих событий, оказали крупномасштабное воздействие на растительность и моря. Морской планктон был, вероятно, в значительной степени уничтожен, что вызвало целый каскад событий, распространявшихся по связям, характеризовавшим экосистемы того времени. Весьма вероятно, что хитросплетения экологических взаимоотношений пришли в хаос. Во время ещё большего по масштабам среди известных человеку эпизодов вымирания, на границе перми и триаса, вулканическая деятельность, которая привела к образованию Сибирских траппов, продолжалась лишь около одного миллиона лет, но охватывала гораздо большую площадь, чем та, которая связана с Деканскими траппами. Образовавшиеся в результате этого вулканические породы в настоящее время занимают территорию примерно в 2 миллиона квадратных километров, меньше трети от оцениваемой первоначальной площади, и всё же почти равны по площади территории Западной Европы. Изменения в химическом составе океана, связанные с этой крупномасштабной вулканической деятельностью, нанесли огромный удар по эволюции жизни в масштабах всего мира. Крупномасштабные акты вымирания оказали воздействие на историю жизни на Земле, и восстановление многообразия групп, которые не полностью вымерли, представляет собой большой интерес и требует понимания особенностей окружающей среды и видов организмов, характеризующих мир после вымирания.Дэвид Яблонски «Origination Patterns and Multilevel Processes in Macroevolution»
Повышение кислотности океана в прошлом и в наше время
После окончания пермского периода, когда произошло самое большое из когда-либо случавшихся событий массового вымирания, ранний триас характеризовался нестабильностью окружающей среды. Из работ геохимиков нам известно, что уровни кислотности океана и количества кислорода в атмосфере колебались. Изменчивые условия оказывали воздействие на метаболизм и в конечном счёте на характер роста у различных групп организмов, включая тех, кто строит скелет из карбоната кальция, например, рифообразующие кораллы (см. главу 10). Повышение кислотности океана — снижение значения pH — воздействовало на характер роста аммонитов и других моллюсков, кораллов и большинства, если не всех остальных групп морских беспозвоночных. Отмеченная вулканическая деятельность привела к достаточно резкому выбросу метана, а также к увеличению концентрации двуокиси углерода не только в атмосфере, но также и в океанах. Это снизило возможность выпадения в осадок морского карбоната кальция и потенциальную возможность обызвествления у организмов, живущих на всех уровнях толщи воды (бентос, планктон) — «кризис биокальцификации», который исследовали мой коллега Хельмут Вайссерт из Швейцарской высшей технической школы Цюриха и многие другие. Исследование ископаемых морских беспозвоночных и истории их жизни зачастую происходит параллельно с изучением изменений в океанах, что сводит воедино геохимию и палеонтологию. Точно так же, как в раннем триасе, в наше время происходит повышение уровня кислотности морской воды по всему миру, поскольку океаны поглощают всё увеличивающееся количество двуокиси углерода в атмосфере. Считается, что это закисление океана является главной угрозой для морских экосистем в ближайшем будущем. Эксперименты показали, что это изменение вредит способности к воспроизводству и росту по крайней мере у некоторых видов веслоногих ракообразных, улиток, морских ежей и офиур. Поскольку изменение происходит очень быстро, многие виды вряд ли смогут выжить. Но не на все группы это воздействие оказывается одинаково. Сэм Дюпонт и его коллеги из Швеции представили экспериментальные свидетельства того, что повышение кислотности океанской воды оказывает положительное воздействие на личинок морской звезды Crossaster: культивируемые при низких значениях pH личинки растут быстрее, без видимого влияния на выживание или формирование скелета. Это был первый отчёт о положительном влиянии повышения кислотности океанской воды на темп роста у какого-либо беспозвоночного. Различающиеся эффекты воздействия на историю жизни организмов должны отвечать за то, кто выживет, а кто станет жертвой при наступлении события массового вымирания и в ходе последующего восстановления биосферы. Как сказали Арно Брейяр и его коллеги, «в выживании групп живых организмов играют свою роль и хорошие гены, и хороший расклад дел».Основные типы эволюции личинок в океанах
Многие виды морских беспозвоночных, обитающие на морском дне, обладают свободноживущей личиночной стадией, которая служит для расселения и очень сильно отличается от взрослой особи морфологически и экологически. Большинство морских беспозвоночных использует одну из двух различающихся между собой стратегий личиночного развития, прежде чем осядет на дно и претерпит метаморфоз в ювенильную форму. При планктотрофии личинки питаются в толще воды на протяжении недель или месяцев. При альтернативной стратегии — не связанной с питанием лецитотрофии — личинки питаются желтком яйца в течение гораздо более короткого времени, которое они тратят на жизнь в толще воды. Как уже предсказало исследование распределения этих стратегий по эволюционным древам ныне живущих видов, в ходе эволюции между ними должны были происходить многократные смены, даже в пределах отдельно взятой группы. Вопрос, который заставляет проводить множество исследований различных беспозвоночных, состоит в том, чтобы понять, были ли эти изменения беспорядочно распределены, или же концентрировались в определённые периоды времени. В целом принято считать, что их движущими силами, вероятнее всего, были возникновение хищничества на океанском дне, когда в процессе эволюции появились новые группы или новые стратегии хищничества, или же изменения в окружающей среде. В дополнение к академическому интересу к этому аспекту эволюции, целью данных исследований также является изучение потоков питательных веществ и экологических взаимодействий в океане, которое в конечном счёте оказывает влияние на рыболовство. Работа с ископаемыми остатками организмов может дать прямую информацию о крупных изменениях сообществ организмов в океанах прошлого, связанных с новыми стратегиями, появляющимися в истории жизни организма. Хороший пример этого — возникновение планктотрофии при переходе от кембрия к ордовику, примерно 490 миллионов лет назад. Этот период времени очень важен, поскольку в это время зарегистрировано резкое увеличение разнообразия жизни по всему миру и появление всех основных групп ныне живущих организмов. Прямое свидетельство эволюции стратегии личиночного развития получено для ископаемых брюхоногих моллюсков. В древние времена, как и в наши дни, некоторые виды имели прямое развитие и, как в случае лецитотрофных личинок, большее количество желтка в их яйцах означало, что у них были более крупные оболочки. Сами оболочки яиц редко сохраняются в ископаемом состоянии, потому что они очень тонкие и мелкие (обычно менее 1 мм). Но их естественные слепки нередки. По-немецки их называют Steinkerne (ядровые камни), и они во многих случаях являются побочным продуктом работы палеонтологов, которые изучают конодонтов, возможно, находящихся в самом основании родословного древа позвоночных, растворяя породу химическими веществами и исследуя остаток. Поскольку размер Steinkerne отражает количество желтка, мы можем с уверенностью делать вывод о том, было ли животное планктотрофным. Александр Нутцель и его коллеги измерили сотни этих Steinkerne из позднего кембрия и раннего ордовика и напрямую зарегистрировали появление планктотрофных форм при переходе между этими периодами приблизительно 490 миллионов лет назад. Для раннего ордовика характерно многообразие бентосных форм, кормившихся взвесью и прикреплявшихся к морскому дну. Некоторые исследователи предположили, что эта особенность запустила эволюцию планктотрофии как стратегии избегания хищничества. Очевидно, что этот вероятный сценарий подразумевает то, что снабжение питательными веществами в мировом океане было достаточным для поддержания существования плавающих личинок. Фактически, это стало результатом того эффекта, который оказали на динамику океанских вод тектонические события в раннем ордовике. Биологи, имеющие дело с океанической жизнью, начинают уделять пристальное внимание работам палеонтологов в этой области, пробуя понять те условия, которые оказывали влияние на организмы в прошлом, и тем самым предугадать, какое воздействие могли бы оказать глобальные изменения окружающей среды на современные организмы. Поток информации движется в обе стороны, и итогом этого становится тщательное изучение истории жизни современных видов, которое даст самое подробное понимание эволюции стратегий личиночного развития в летописи окаменелостей.Климатические изменения и эволюция индивидуального развития у млекопитающих
Исследования характера распределения биологического многообразия и изменений окружающей среды в разрезе геологического времени показало, что в фаунах позвоночных имели место эпизоды изменений, вызванные климатическими изменениями, но были также и такие, при которых не зарегистрировано никаких следов изменений. В ходе климатических изменений в неогене Северной Америки, которые привели к преобразованию обширных областей в пустыни или в очень сухие местообитания, возникло несколько новых групп млекопитающих. Была выдвинута гипотеза о том, что это произошло путём схожих онтогенетических изменений в нескольких родословных линиях одновременно. Передвижение скачками на задних конечностях возникло в сухих местообитаниях по всему миру (напр. у кенгуру в Австралии). Этот вид передвижения связан с различными пропорциями конечностей, которые появляются в процессе эволюции путём возникновения различий в скорости или времени начала роста у разных видов.[65] Это означает, что климатические изменения в итоге могли оказать одновременное воздействие на характер индивидуального развития в различных группах живых организмов, которые приобретают в процессе эволюции схожие адаптации к новым средам обитания. Изменение климата может приводить к эволюционным изменениям размеров, которые обычно связаны с изменениями в истории жизни. Виды лошадей, когда-то широко распространённые и в изобилии представленные в североамериканских фаунах, демонстрируют быстрое уменьшение в размерах перед своим исчезновением в плейстоцене Аляски. Возможно, что естественный отбор оперирует переменными компонентами истории жизни, ведущими к эволюционным изменениям размеров тела.Исследование роста и продолжительности жизни, оставивших свой след в гистологии зубов, может выявить такие изменения. С точки зрения морфологии, из-за аллометрических отношений изменения в размерах во многих случаях означают изменения в форме анатомических структур организмов. Многочисленные примеры этого процесса возникли параллельно во многих группах животных в Африке около трёх миллионов лет назад. Аналогичные явления, возможно, происходили в разное время на других континентах.
Рисунок 30. В процессе эволюции прыгающие млекопитающие независимо появились во многих пустынных областях по всему миру ближе к концу миоценовой эпохи. Пример такого рода — сумчатое млекопитающее Argyrolagus scagliai из Аргентины.[66] Хвост, вероятно, был намного длиннее, и здесь изображены только сохранившиеся позвонки образца из Museo Municipal de Mar del Plata. У современных форм и, возможно, также у прыгающих аргиролагид большая разница в размерах конечностей развилась путём изменения индивидуальных скоростей дифференциации скелета. Из Simpson 1970, с изменениями.
Эволюция млекопитающих в Южной Америке — это благодатный объект для исследований в области палеонтологии позвоночных, но лишь недавно знание хронологии и репрезентативность выборок фауны стали достаточно хорошими, чтобы позволить нам уверенно исследовать характер эволюции. Панчо Гоин, Фреди Карлини и их коллеги из Ла-Платы недавно заметили, что значительные изменения в фауне млекопитающих совпадают с резким снижением глобальных температур в самом конце эоцена — раннем олигоцене. Они назвали это событие Bisagra Patagónica, или «Патагонский шарнир», и оно было зарегистрировано в Южной Аргентине. Многие группы сумчатых и родственников броненосцев вымерли в конце эоцена, и много новых видов этих животных появилось в олигоцене.[67] И что примечательно у раннеолигоценовых групп, так это их новая стратегия в онтогенезе зубов — состояние, когда зубы постоянно растут, называемое гипсодонтией. Эта особенность включает новую совокупность изменений в метаболизме и в истории жизни, и, вероятно, возникла как ответ на износ зубов, вызванный главным образом присутствием вулканического пепла в пище, как выяснил в ходе исследований Рик Мэдден. В противоположность вышеприведённым примерам, существенное снижение температуры, которое произошло в Северной Америке в то же самое время, что и Bisagra Patagónica в Аргентине (на границе эоцена и олигоцена, примерно 34 миллиона лет назад), явно оказало лишь небольшое влияние на эволюцию млекопитающих. Опираясь на превосходные коллекции многочисленных видов млекопитающих из отложений свиты Уайт-Ривер с запада Соединёных Штатов, Дон Протеро и его коллеги показали, что подавляющее большинство видов млекопитающих не изменилось морфологически, несмотря на явно выраженные климатические изменения. Изучение поверхности зубов показало, что должны были произойти изменения в рационе, хотя форма зубов осталась той же самой. Возможно, млекопитающие Северной Америки в том месте и к тому времени приобрели в процессе эволюции большую фенотипическую пластичность, чем их современники в Южной Америке. С другой стороны, хотя эоцен-олигоценовое изменение климата было глобальным, его последствия могли отличаться на локальном уровне, при различной степени вулканической активности на этих двух континентах, что в этом отношении является важным фактором, который стоит учитывать. Воздействие температуры на эволюцию индивидуального развития хорошо прослеживается во многих эмпирических исследованиях ныне живущих видов. Также существуют косвенные подсказки, которые предоставляет летопись окаменелостей. В эоцене, за миллионы лет до более холодного олигоцена, существовало множество разнообразных ископаемых фаун с такими же признаками, как у тропических фаун наших дней, даже в высоких широтах. В современной северо-восточной Колумбии примерно от 58 до 60 миллионов лет назад жила примечательная гигантская змея Titanoboa cerrejonensis, длина тела которой оценивается в 13 метров, а масса в 1140 килограммов. Поскольку температуры повышались и снижались с течением геологического времени, то же самое происходило с верхним пределом размера холоднокровных существ, таких, как змеи, обмен веществ у которых находится под влиянием средней температуры окружающей среды. Джейсон Хэд с сотрудниками подсчитал, что для выживания змеи размером с Titanoboa требовалась бы круглогодично средняя температура от 30 °C до 34 °C. Для сравнения, среднегодовая температура сегодняшней Картахены на побережье Колумбии — около 28 °C.
Рисунок 31. Общая картина вымирания и роста биологического разнообразия в нескольких группах сумчатых млекопитающих во времена крупных климатических изменений, отмеченных для границы эоцена и олигоцена, которые Панчо Гоин и его коллеги назвали Bisagra Patagónica. По Goin, Abello и Chornogubsky, 2010, с изменениями.
Юрские акулы
Способ воспроизводства и способность эволюционировать, адаптироваться в течение немногих поколений, могут изменяться на протяжении эволюционного времени как ответ на изменения окружающей среды абиотического и биотического характера совместно с внутренними изменениями. Пример этого явления показывает ранний этап увеличения видового разнообразия и адаптивной радиации современных акул и скатов в ранней юре, изученный Юргеном Криветом и его коллегами. После волны массового вымирания конца триаса начались оппортунистическое быстрое увеличение видового разнообразия и активная экологическая радиация у акул и родственных им форм, который были связаны с мелкими размерами тела, небольшой продолжительностью жизни и размножением путём откладывания яиц, и, вероятно, были вызваны этими же обстоятельствами. Эти особенности истории жизни позволили быстрее возникать экологическим новшествам в плане строения тела. Итоги исследования современных акул, скатов и химер показали, что у живородящих видов риск вымирания выше, чем у яйцекладущих, даже в случае пропорционально более высокого уровня выедания хищниками особей и яиц в случае яйцерождения. Так происходит из-за того, что эволюционная приспособленность живородящих видов оказывается очень низкой в случае гибели беременных самок, и это делает их более подверженными риску исчезновения. В ранней юре яйцерождение позволило акулам и их родственникам лучше приспособиться к изменению условий окружающей среды. Большая продолжительность жизни и живорождение у большинства ныне живущих видов акул делают их сильно уязвимыми в случае массового вымирания.Выживание и рост многообразия на раннем этапе эволюции млекопитающих
Очень немногие виды наших далёких зверообразных предков пережили пермо-триасовый эпизод массового вымирания примерно 250 миллионов лет назад. Большинство родословных линий прервалось, и многие из их представителей были плотоядными формами. Выжила лишь малая часть этих ранних родственников млекопитающих, и среди них был Lystrosaurus, наиболее многочисленный вид четвероногих позвоночных во время раннетриасового периода восстановления фауны после массового вымирания. Lystrosaurus принадлежит к числу дицинодонтов, получивших название из-за своей специализированной зубной системы, включавшей лишь два клыка. Эти травоядные были крупными, размером от крысы до быка, и разнообразными: известно более семидесяти их родов. Lystrosaurus не только выжил, но также широко распространился географически и был многочисленным в эпоху раннего триаса. На основании своих собственных палеонтологических исследований Дженнифер Бота-Бринк установила, что у Lystrosaurus и других дицинодонтов была высокая скорость роста по сравнению с другими современными им видами. Гистология более чем сотни образцов костей дицинодонтов демонстрирует увеличенные каналы в кортикальном слое, которые могли усиливать снабжение кислородом и питательными веществами, способствуя более эффективной ассимиляции питательных веществ и ускорению роста кости. Lystrosaurus и его ближайшие родственники росли быстрее, чем его эволюционные «кузены». Исключительно высокая скорость роста, вероятно, способствовала выживанию Lystrosaurus во время массового вымирания в конце перми и его многочисленности в раннем триасе, в фазе восстановления фауны после вымирания. Ускоренный рост дицинодонтов, возможно, позволял им быстро достигать размеров тела, обеспечивающих безопасность от нападения хищников. Кроме того, гистология костей предполагает быстрое достижение половой зрелости, которое придало бы Lystrosaurus устойчивость к колебаниям условий окружающей среды. Это открытие, связанное с историей жизни, также соотносится с обычностью ископаемых остатков этих животных, указывая на их многочисленность в отложениях, где отмечено наличие лишь немногих конкурирующих видов травоядных.Острова как место экспериментов в эволюции истории жизни
Острова — это любимый объект исследований для эволюционных биологов, поскольку они стоят ближе всего к естественному эксперименту, с помощью которого проверяются гипотезы. На ограниченном по площади месте с чётко очерченными границами значительно легче документально зафиксировать ключевые физические и биологические переменные величины и изучить, как они воздействуют на эволюционный процесс. Классические исследования островных популяций — это исследования галапагосских земляных вьюрков, ящериц рода Anolis с островов Карибского моря и плодовых мушек с Гавайских островов. Озёра, такие, как Виктория в Африке, также являются местами, где могут проводиться большие естественные эксперименты, поскольку некоторые группы рыбы могут существовать только в своём озере.[68] Острова также являются важным объектом исследований в области биологии развития и изучения ископаемых форм. Многие из изучаемых организмов вымерли лишь недавно, в течение последних сотен или тысяч, а не миллионов лет. В большинстве случаев причиной их исчезновения было пагубное воздействие со стороны человека. Существуют различные виды островов: старые и молодые, маленькие и большие, океанические и континентальные. Континентальные острова — это такие, которые в прошлом соединялись с соседними массивами суши. Они сами по себе находятся на континентальном шельфе и могут соединяться, или же ранее соединялись с материком посредством сухопутного перешейка. Бали, Куба и большинство островов Средиземноморья являются континентальными. Океанический остров — это остров в своём наиболее чистом виде. Он никогда не был связан с материком, и он образуется посредством некоторых геологических процессов (чаще всего после извержения вулкана), и поднимается со дна океана. Галапагосские и Гавайские острова, Маврикий и Реюньон — это всё вулканические острова. Что здесь касается моей темы, так это изоляция, которая имеет место на островах, и вместе с ней специфические условия, которые привели к эволюции различных видов истории жизни; многое из этой область может быть зафиксировано в летописи окаменелостей. В отношении ископаемых млекопитающих нами изучены одни из самых потрясающих или, по крайней мере, наилучшим образом известные примеры эволюции на островах.[69] В ходе одного из исследований подсчитано, что скорость морфологической эволюции у млекопитающих, главным образом у грызунов, на островах выше, чем у материковых популяций, вплоть до соотношения три к одному.[70] Ускоренные изменения на островах предполагались на протяжении долгого времени, и исследования их количественных характеристик включают, например, исследования Адриана Листера, касающиеся популяции благородного оленя на острове Джерси. Летопись окаменелостей показывает многочисленные резкие изменения размеров в классических случаях слонов размером с пони, мышей размером с кролика, бегемотов размером со свинью и оленей размером с собаку. После образования репродуктивной изоляции в процессе эволюции возникают нетипичные размеры, тесно связанные с изменениями в истории жизни.[71] Из-за явления аллометрического роста итогом разницы в размерах являются различия в формах организмов. Поэтому онтогенетический подход к изучению морфологической эволюции островных млекопитающих является интуитивно наиболее верным. Существует общая тенденция: мелкие млекопитающие эволюционируют в сторону увеличения размера, а крупные виды — в сторону уменьшения размеров в течение коротких отрезков геологического времени.[72] Существует много примеров такого рода, вроде вымершего сицилийского слона Elephas falconeri, у которого, по оценкам, рост взрослой особи был менее 1 метра, а масса тела-100 килограммов, лишь примерно 1 процент от массы его материкового предка. Другие примеры родственных слонам животных обнаружены на островах Ченнел, в двадцати милях от побережья Южной Калифорнии. Там эволюционировал Mammuthus exilis, карликовый вид, произошедший от Mammuthus columbi, одного из нескольких видов мамонтов, которые бродили по Северной Америке на протяжении последнего миллиона лет. Карликовые слоны были обычными обитателями Средиземноморья; они обнаружены на Крите, Мальте, Кипре и Сицилии. Было высказано предположение, что черепа этих животных, возможно, послужили первоисточником для греческих легенд о циклопах: у слоновых черепов есть большое отверстие во лбу, с которым соединяется хобот, и которое могло быть по ошибке принято за единственную огромную глазницу, находящуюся в центре лба. В противоположность причудливой ассоциации носового отверстия слона с единственным глазом мифического существа анатомически точный подход делает альтернативный источник вдохновения, циклопическое состояние уродливого младенца, менее вероятным. У мифического существа единственный глаз расположен выше носа, а в случае уродства глаз развивается ниже его.[73]
Рисунок 32. Череп карликового слона Elephas falconeri из позднего плейстоцена Сицилии. Изображение на основе образца, выставленного в Музее природы Зенкенберг, Франкфурт. Рисунок Клаудии Джоэл.
Изучение млекопитающих с различных островов выявляет черты сходства у видов, которые их населяют, и в адаптациях, которые они приобрели в процессе эволюции. Многочисленные ископаемые остатки подтверждают существование в плейстоцене вымерших слонов, оленей и бегемотов на многих островах Средиземноморья, а также оленей и слонов на островах Японии и Юго-Восточной Азии. Таксономическое сходство вероятнее всего объясняется способностями к плаванию в случае крупных видов или способностью выживать на естественных плотах в случае мелких. Существуют также экологические параллели в условиях, с которыми сталкиваются различные млекопитающие в различных регионах. Каждый остров уникален, и в каждом случае различные в силу естественных причин характер воздействия естественного отбора и исторические обстоятельства играли свою роль в формировании фаунистических и флористических сообществ. Однако, принципы и механизмы их воздействия, похоже, универсальны. Экологический контекст индивидуального развития подходит для понимания случаев островных карликовости и гигантизма, явлений, в ходе возникновения которых должны участвовать, переплетаться и взаимно влиять друг на друга биологические переменные величины: размер, продолжительность жизни, возраст достижения половозрелости, скорость роста, доступные ресурсы, конкуренция и потребности обмена веществ. То, как организмы поддерживают процесс своего роста, зависит от темпов потребления пищи и расхода энергии. Часть потреблённой пищи окисляется и используется в энергетических превращениях, а остальное участвует в реакциях синтеза и откладывается в виде биомассы. Были разработаны математические модели, основанные на физиологических измерениях обмена веществ и процесса роста, и они помогли установить, что у животных из различных групп работают одни и те же общие принципы отношений.[74] Это может объяснить, почему мы наблюдаем параллельное развитие морфологических особенностей на различных островах. Есть два пути, приводящих к быстрой эволюции изменений размера на островах. Позднее наступление половозрелости может привести к появлению большего размера тела. Такая картина развития связана с появлением немногочисленного потомства. С другой стороны, оптимизация метаболизма под влиянием иных условий с иным, более ограниченным выбором ресурсов, может привести к появлению способности к воспроизводству в раннем возрасте и к небольшому размеру. Как ископаемые млекопитающие ведут себя в условиях иного, меньшего количества ресурсов, с которым им пришлось столкнуться на островах? Вновь и вновь ископаемые остатки предоставляют документальные свидетельства значительной эволюционной пластичности в стратегиях, связанных с индивидуальным развитием и физиологией, во многих группах. Примером этого является Myotragus, близкий родственник овец и коз, который вымер около пяти тысяч лет назад. Предок Myotragus, вероятно, добрался до Балеарских островов около шести миллионов лет назад, в то время, когда Средиземноморье представляло собой группу солёных озёр. Myotragus, вероятнее всего, эволюционировал в условиях, которые очень сильно отличались от тех, которые существовали на материке: при небольшом количестве ресурсов и при отсутствии хищников. Островной Myotragus стал исключительно мелким животным: самые крупные взрослые экземпляры достигали максимального роста от 45 до 50 сантиметров от земли до холки и, вероятно, весили не более 50–70 килограммов. Как они росли, какова была их энергетическая стратегия? Гистологическое исследование длинных костей Myotragus показало наличие по всему кортикальному слою ламеллярной кости — вида кости, обычного в прочих случаях только среди современных рептилий вроде крокодилов, которые являются эктотермными животными. Гистология костей показывает, что Myotragus рос медленно, не имел единой скорости роста на протяжении жизни, и это было связано с существенной задержкой достижения скелетной зрелости. У Myotragus также была гораздо большая продолжительность жизни по сравнению с его эволюционными кузенами. Не затронутый деятельностью человека, Myotragus вёл медленную и долгую жизнь на острове в Средиземноморье. В целом считалось, что явление педоморфизма (сохранение во взрослом состоянии инфантильных или ювенильных признаков) способно описать многое в эволюции островных млекопитающих. Но организм сам по себе мозаичен, и в нём каждый орган или часть тела, даже будучи объединённым с остальными частями, управляется различными правилами относительного роста. Так, хотя островная форма может обладать общим сходством с ювенильной особью предкового вида, оно может быть лишь поверхностным, а многие из особенностей строения будут демонстрировать специальные адаптации к новой экологии и к новым метаболическим потребностям. Пропорции тела у островных форм нельзя объяснить просто как результат остановки или замедления развития, что придало им сходство с ювенильной стадией предка. Открытие нового вида гоминид из плейстоцена индонезийского острова Флорес, знаменитого Homo floresiensis, даёт нам ясный пример необходимости рассмотрения онтогенеза при исследовании ископаемых остатков и важности явления «мозаичной» эволюции. H. floresiensis был лишь около одного метра ростом и был полностью двуногим; для него был реконструирован мозг, похожий на мозг шимпанзе, объёмом 417 кубических сантиметров для примерно 30-килограммового тела. Такой размер мозга озадачивает, поскольку этот вид делал орудия труда и жил совсем недавно, а его предок-гоминид обладал намного большим размером мозга по сравнению с ним — возможно, это был H. erectus, объём мозга у которого был примерно 900 кубических сантиметров. Вот, почему некоторые учёные утверждали, что H. floresiensis был ничем иным, как популяцией карликов с патологическим состоянием, которое включало микроцефалию.[75] Но, если не брать во внимание эту опровергнутую альтернативу, то, рассматривая аллометрические отношения размера мозга к размерам тела, можно увидеть, что объём мозга H. floresiensis лежит вне рамок ожидаемого, вне зависимости от того, какой именно вид гоминид из потенциальных кандидатов предполагался в качестве его предка. Примеры чрезвычайно сильного сокращения размера мозга, выпадающего за ожидаемые значения, определяемые с позиций аллометрического подхода, известны для других млекопитающих. Элеонор Уэстон и Адриан Листер показали это в количественном отношении для ископаемых гиппопотамов с Мадагаскара. Мозговая ткань требует очень больших затрат в плане метаболизма, и потому была выдвинута гипотеза о том, что этот орган уменьшится в размерах в эволюционных последовательностях, где было выгоднее экономить энергию. Отсутствие простого онтогенетического масштабирования — не редкость в случае быстрого эволюционного уменьшения размеров. Например, Мейке Кёлер и Сальвадор Мойя-Сола сообщили о значительной редукции мозга и органов чувств у Myotragus в ходе его географической изоляции на Майорке. В другом примере Луиза Рот сообщила в 1980-х годах, что островные карликовые виды ископаемых хоботных не просто мелкие с сохранившимися масштабными соотношениями (педоморфными) в пропорциях их посткраниального скелета, но также демонстрируют отклонения, которые, вероятнее всего, являются ответом на локомоторные и метаболические ограничения и адаптации, что было также показано для многих ископаемых видов с островов Средиземноморья. Картина, наблюдаемая у гиппопотамов, Myotragus и, вероятно, у Homo florensiensis, имеет свою параллель в виде процесса одомашнивания, при котором происходит сокращение размеров мозга и редукция органов чувств, что уже отметил Дарвин в своей вышедшей в 1868 году книге об одомашнивании. Фактически, в одной заметке некоторые авторы ошибочно заявили, что морфология Myotragus — это результат не адаптации к новому островному местообитанию, а адаптаций, являющихся итогом одомашнивания. Пример важной особенности в истории жизни, которую стараются отбирать разводчики — это достижение половой зрелости при небольших размерах. Эта особенность явно выглядит более отзывчивой на применение искусственного отбора, чем особенности относительного роста или строения тела.[76] К пониманию факторов, принимающих участие в процессе быстрой эволюции, которую демонстрируют ископаемые остатки, найденные на островах, можно также прийти при изучении воздействия содержания животных в неволе в зоопарках и в других местах. Эта задача была бы аналогична той, которую поставил перед собой Дарвин, когда изучал изменения под воздействием одомашнивания, желая получить понимание процесса эволюции в целом. Содержание в неволе оказывает значительное воздействие на основные аспекты биологии животных, такие, как размеры тела и продолжительность жизни. Изменения, происходящие на протяжении времени жизни отдельной особи, конечно, не наследуются, но можно представить себе, как эти условия неволи, особый режим отбора, могут привести к эволюционным изменениям, если они сохраняются на протяжении ряда поколений. В условиях неволи четыре основополагающих аспекта жизни отличаются от естественных условий: доступность пищи, отсутствие хищников, ограниченное пространство для жизни и меньшее разнообразие или меньшее количество видов, сосуществующих в одном местообитании. Эти аспекты также отличают друг от друга островную и материковую популяции.Глава седьмая Ископаемые остатки и генетика развития
Онтогенез в ископаемом виде не сохраняется. Но сохраняются структурные образования, которые некогда были результатом процесса развития. В этом случае ископаемые остатки взрослых особей могут дать информацию о развитии благодаря сохранению фенотипов, обладающих непосредственной и чёткой корреляцией с определённым процессом, протекающим в ходе индивидуального развития. Чтобы реконструировать такой процесс, важно изучить положение ископаемой формы на эволюционном древе жизни, чтобы быть уверенным в том, что исследования основаны на правильном предположении. Этот подход называется заключением в современные филогенетические скобки{6}. Он был введён специально для того, чтобы можно было делать вывод об анатомии мягких тканей и создавать реконструкции поведения для ископаемых форм, но его также можно использовать для реконструкции онтогенеза вымерших форм. Вымершее животное сравнивается с его ближайшими ныне живущими родственниками. Исследуется распределение некоторой особенности развития среди существующих в наше время таксонов, которые «окружают» вымерший таксон на данном древе. Если особенность существует у ныне живущих видов, входящих в группу, внутри которой находится изучаемая ископаемая форма, следовательно, можно сделать вывод о том, что она существовала у общего предка, и вследствие этого также у ископаемой формы.
Рисунок 33. Динозавры заключены в филогенетические скобки между крокодилами и птицами.
Например, динозавры заключены в «скобки» между птицами и крокодилами. Особенность, имеющаяся и у птиц, и у крокодилов, например, яйцерождение, вероятно, присутствовала бы и у динозавров. С другой стороны, особенность, отсутствующая и у птиц, и у крокодилов, вроде метаморфоза, вероятнее всего, также отсутствовала бы у динозавров. Но дело усложняется, когда у одних из ныне живущих родственников особенность имеется, а у других её нет. Например, крокодилы обладают костями преимущественно ламеллярного типа, которые коррелируют с медленным ростом, тогда как птицы обладают фиброламеллярными костями и способны к быстрому скачкообразному росту. А как обстояли дела у динозавров? В таких неоднозначных случаях, а также для проверки выводов, основанных исключительно на использовании метода филогенетических скобок, особенно важным является рассмотрение случаев сохранения деталей, которые представляют прямые свидетельства существования той или иной особенности. В случае с процессом роста у динозавров исследования гистологии костей дали прямую и определённую информацию о наличии фиброламеллярной кости и своеобразных кривых роста со стадией быстрого роста у некоторых групп этих животных. Можно поставить под вопрос научную ценность выводов, сделанных в отношении организмов, отделённых от нас несколькими сотнями миллионов лет независимой эволюции. Но такие выводы подтверждены тем, что они основаны на эмпирических свидетельствах, собранных на материале ныне живущих организмов. Крупное открытие нескольких последних десятилетий состоит в том, что многие механизмы индивидуального развития и молекулы остаются одинаковыми для таксонов, не сравнимых друг с другом напрямую и отдалённо родственных филогенетически. Известно, что многие молекулярные процессы, которые контролируют изменение фенотипа, сами по себе намного старше, чем группа, у которой эти фенотипы проявляются; Шон Кэрролл назвал это явление «предковой сложностью». Этот консерватизм имеет место у очень непохожих друг на друга и весьма отдалённо родственных видов. Это ставит нас перед фактом наличия глубоких, или фундаментальных уровней, находящихся на ранних отрезках геологического времени и времени индивидуального развития, до которых может быть отслежена история структурных образований, имеющихся у взрослых особей таких отличающихся друг от друга видов, как мухи и люди. В этом случае мы говорим о «глубинной гомологии» этих образований, подразумевая «тождество», или гомологию, на некотором уровне организации (молекулярном, клеточном) и на некотором уровне иерархии на эволюционном древе жизни. Может показаться, что использование настоящего для того, чтобы делать выводы в отношении прошлого, как в случае с «филогенетическими скобками», накладывает на него свои ограничения. По сути, эволюционный биолог Марк Пэйджел пишет, что этот подход «обрекает прошлое на сходство с настоящим. И, возможно, ещё хуже то, что прошлое, картину которого мы получаем, оглядываясь назад — это очень обычное прошлое, усреднённое прошлое». Но у нас нет выбора, и это не означает того, что мы не обнаруживаем важных, и даже неожиданных примеров, когда рассматриваем окаменелости таким образом. И вообще, что такое «обычное» или «усреднённое»? Не существует ни одного современного трилобита, но есть все основания, чтобы полагать, что разделение тела на сегменты, которое они претерпевают в ходе индивидуального развития, было связано с теми же самыми процессами, какие выявлены у ныне живущих членистоногих. В приведённых ниже примерах я постарался продемонстрировать, как мы можем изучать эволюцию индивидуального развития, рассматривая фенотипы взрослых особей вымерших видов.Количество позвонков
Сложные животные состоят в значительной степени из повторяющихся частей, и позвоночные здесь не являются исключением. Это не столь очевидно на первый взгляд, но, если мы исследуем наш собственный скелет или скелет любого другого позвоночного, то мы увидим последовательное повторение позвонков, лишь плавно отклоняющихся друг от друга по размеру и по форме. Ткани, окружающие эти позвонки, также образуют повторяющиеся структуры, в том числе мышцы, нервы и сосуды. Эти «пакеты» анатомических структур, эти сегменты целого тела, возникают в ходе индивидуального развития из строительных блоков, или сомитов. Сомиты образуются на протяжении особого отрезка времени раннего эмбрионального развития. Скорость сегментации определяет в этом случае, сколько позвонков будет иметь взрослый организм. У некоторых змей развивается около трёхсот сегментов, тогда как черепахи останавливаются примерно на двадцати пяти, со строго определённым количеством, восемнадцатью, перед крестцом (часть позвоночного столба, находящаяся прямо перед костями хвоста). Есть своего рода часы сегментации, и в зависимости от вида они тикают по-разному. Иногда они тикают дольше или быстрее в пределах одного вида, особенно в области хвоста. Наш собственный позвоночный столб легко разделяется на отделы, обладающие своими отличительными особенностями. У нас нет никаких рёбер в шее, в шейном отделе. У нас есть грудной отдел с большими рёбрами, которые сходятся друг к другу, чтобы прикрепиться к грудине, и которые образуют клетку, где наши сердце, желудок и другие органы надёжно защищены. У нас нет грудной кости или рёбер на животе, поэтому мы должны удерживать органы на своих местах, приобретая сильную брюшную мускулатуру в нашем поясничном отделе. Наконец, некоторые позвонки слились, чтобы образовать крестцовый отдел, к которому прикрепляется таз, или пояс задних конечностей, а за ним следует каудальный отдел, или хвост. Границы между отделами соответствуют границам экспрессии специальных генов, которые регулируют процесс развития и устанавливают признаки отличий отделов, образующихся вдоль оси тела. Эти гены — члены семейства регуляторных генов Hox. В этом контексте процесса индивидуального развития вещи, кажущиеся обыденными, вроде количества позвонков в различных отделах скелета, выходят в новое измерение. Количественные значения отражают два фундаментальных аспекта раннего этапа развития: образование сомитов и границы экспрессии генов Hox. Тогда изменчивость в количестве позвонков должна отражать эволюционную пластичность этих механизмов. Млекопитающие консервативны в плане количества предкрестцовых позвонков. И у мыши, и у жирафа имеется семь шейных позвонков. И коала, и опоссум, которые оба фактически являются сумчатыми, обладают двенадцатью позвонками в грудо-поясничном отделе, и изменчивость в этой области мала у этой группы в целом. Рептилии, которые включают птиц, гораздо более разнообразны в этом отношении. У лебедя есть около двадцати пяти шейных позвонков, а у утки шестнадцать или меньше. У всех черепах восемнадцать предкрестцовых позвонков — восемь шейных, десять спинных и два крестцовых — хотя примитивная черепаха Odontochelys из триаса Китая, как сообщают, обладает лишь девятью спинными позвонками. Я впервые узнал о существовании этих жёстких шаблонов от Шигеру Куратани.[77] Куратани-сэнсей, как я называл его, будучи в Кобэ, в своей жалкой попытке сработаться с ним, догадался исследовать проблему количества позвонков среди наземных позвоночных, отобразив в виде карты данные, изложенные в классическом труде покойного сэра Ричарда Оуэна, современника Чарльза Дарвина. Совместно с Юичи Наритой он выяснил, какие группы консервативны в плане индивидуального развития, а какие пластичны. Посетив его лабораторию в 2006 году и обсуждая их анализ, я заметил в их работе другую своеобразную картину. У большинства млекопитающих из всех групп сумма числа позвонков в грудной клетке и в области живота, в грудо-поясничной области, равна девятнадцати. Это количество постоянно и у сумчатых, и у яйцекладущих однопроходных, и у многих групп плацентарных млекопитающих, и, вероятно, закрепилось на ранней стадии эволюции млекопитающих. Но, кроме некоторых других беспорядочно распределяющихся исключений, крупная группа плацентарных млекопитающих Afrotheria обладает общим признаком — увеличенным суммарным количеством грудных и поясничных позвонков. Эта картина сохраняется даже тогда, когда исследуются самые древние и наименее специализированные ископаемые виды каждой из этих групп. Afrotheria, группа, изначально признанная лишь на основании молекулярных свидетельств, включает таких разнообразных существ, как даманы, слоны, дюгони, тенреки и златокроты. После того, как мы заметили эту картину, Роб Ашер и Томас Лехманн открыли, что задержка в развитии зубов также является диагностическим признаком для афротериев, как ископаемых, так и ныне живущих.[78] Формирование количества позвонков связано с механизмами, которые у вымерших видов невозможно изучить непосредственно. Но у количества позвонков существует однозначное соответствие сегментации и границам экспрессии Hox-генов. Существующие в наше время филогенетические скобки чётко поддерживают предположение о том, что эти процессы работают у всех наземных позвоночных, ископаемых и современных. Тогда богатая летопись окаменелостей наземных позвоночных даёт нам шанс исследовать индивидуальное развитие у вымерших форм. Таким способом мы смогли исследовать раннюю историю млекопитающих и рептилий, чтобы увидеть картину эволюции и ответить на ряд вопросов: Когда в процессе эволюции возник консерватизм млекопитающих в количестве позвонков? Какой из механизмов был эволюционно более пластичным — сомитогенез или границы экспрессии Hox-генов? Какие ограничения накладывают на пластичность механизмов индивидуального развития экологические адаптации, такие, как вторичноводный образ жизни, наблюдаемый у китов и морских рептилий? Чтобы ответить на эти вопросы, мы вместе с Йоханнесом Мюллером и Торстеном Шейером собрали группу экспертов, которые предоставили актуальную и исчерпывающую информацию по филогенезу и количеству позвонков для групп, на которых они специализировались. Обладая этой информацией, мы провели анализ, результатом которого было выявление различной степени пластичности процессов развития в эволюции вымерших групп и в ранней истории всех основных ныне живущих групп. Экспертный анализ ископаемых остатков показал, что увеличение и уменьшение общего количества позвонков или в границах отделов позвоночника, при сохранении одинакового общего количества, возникало независимо в нескольких эволюционных линиях. Мы отследили назад во времени появление канальности и консерватизма в количестве предкрестцовых позвонков у млекопитающих до момента возникновения этой группы, по крайней мере, около 315 миллионов лет назад, когда количество шейных позвонков колебалось между пятью и шестью. Установление почти постоянного количества шейных позвонков, равного семи, произошло приблизительно на 100 миллионов лет позже, в юрский период. Это открытие подвергает сомнению выдвинутую ранее гипотезу, пытающуюся объяснить консерватизм, впервые отмеченный только у ныне живущих млекопитающих. На основании факта высокой частоты смертности зародышей и молодых особей человека с ненормальным количеством шейных позвонков для объяснения консерватизма в количестве шейных позвонков у млекопитающих предполагалось наличие летального плейотропного эффекта, относящегося к мутациям в области регуляции Hox-генов, которые оказывают воздействие не только на осевой скелет, но также и на появление клеточных новообразований. Лишь млекопитающие с низким темпом обмена веществ демонстрируют отклонения от стандартного количества шейных позвонков, и эта особенность также связана с более низкой подверженностью раковым заболеваниям. Однако наш результат подвергает сомнению эту гипотезу. Самые ранние представители эволюционной линии, ведущей к млекопитающим, уже были весьма консервативными в плане количества шейных позвонков, а их физиология, как видно из палеогистологических данных и других характерных анатомических особенностей, была рептильной. Возможно, это пример того, что Томас Гексли назвал «убийством прекрасной гипотезы ужасным фактом».
Рисунок 34. Tanystropheus longobardicus. Другие реконструкции этого животного изображают его с шеей, которая насчитывает тринадцать позвонков и вытянута перпендикулярно главной оси тела. Из Wild 1974, с изменениями, сделанными Кристианом Клюгом на основе работы Tschanz 1988.
Другую крупную закономерность в эволюции количества позвонков мы отмечаем в группах, которые населяют водные местообитания. В этих группах имеет место отклонение от стандартной картины развития: в процессе эволюции появляется большее количество предкрестцовых позвонков. Это справедливо для всех рептилий и млекопитающих. Ослабление регионализации — то есть, когда различия между группами позвонков исчезают и они становятся похожими друг на друга — похоже, способствует проявлению большей пластичности в развитии.[79] Но в случае водных млекопитающих количество шейных позвонков остаётся постоянным. Это отличает их от рептилий, у которых общее количество позвонков (посредством сомитогенеза) также увеличивается, но наряду с ним увеличивается и пластичность в количестве шейных позвонков, что показывают различные вымершие группы вроде плезиозавров, у которых много шейных позвонков, или пахиплеврозавры и Tanystropheus, у которых количество шейных позвонков намного меньше. Картина, наблюдаемая у динозавров, ясно показывает, что количество позвонков и их распределение по отделам позвоночника не определяется общим размером, которого достигает животное во взрослом состоянии: эмбрионы зауропод не показывают высокой скорости сомитогенеза ради достижения гигантского размера, характерного для этой группы.О Saurichthys и рыбах-мутантах
Я обсуждал главным образом то, как палеонтологические данные могут дополнить наше понимание эволюции истории жизни и механизмов индивидуального развития. Но могут данные из области эволюции индивидуального развития сообщить что-либо палеонтологам? Могут ли открытия из области молекулярной эволюционной биологии развития внести свой вклад в понимание разнообразия вымерших таксонов? Поскольку нуклеиновые кислоты и белки очень редко сохраняются с течением геологического времени, реконструкция молекулярной основы морфологических изменений у ископаемых форм должна основываться на умозаключениях. Здесь я исследую основания для таких выводов, рассматривая их на примере ископаемой рыбы. Среди богатых коллекций среднетриасовых позвоночных в Цюрихе есть замечательная рыба. Внешне она выглядит похожей на современную щуку, с удлинённой и обтекаемой формой тела, которая выдаёт в ней хищника. Её название — Saurichthys, «рыба-ящерица». Этот род располагается довольно близко к основанию родословного древа костных рыб, близко к осетрам, веслоносам и ильной рыбе, своим ныне живущим родственникам. Существуют четыре вида, известных из местонахождения в Монте-Сан-Джорджио в Швейцарии, и они выставлены в Цюрихе, но род имеет всесветное распространение и включает более тридцати видов. Те виды, которые имеются в нашем музее, наиболее очевидно характеризуются различным рисунком и количеством чешуй, как было установлено в 1980-х годах Оливье Риппелом, когда он работал в Цюрихе. Гипотетический последний общий предок, давший начало радиации видов Saurichthys, может быть уверенно реконструирован на основании нескольких других окаменелостей как полностью покрытый однородной ромбовидной чешуёй, с многочисленными, сильно сегментированными плавниковыми лучами и с большим количеством дермальных костей в черепе. Виды рода Saurichthys, происхождение которых можно отследить назад к этому гипотетическому предку, характеризуются различной степенью редукции чешуйного покрова (ряды и вытянутая форма чешуй), редукцией количества плавниковых лучей и их сегментов, сокращением количества дермальных костей в черепе и их формой, отражающейся в форме жаберной крышки, а также снижением степени массивности зубов и челюстей. Фактически, новизна в эволюции во многих случаях ассоциируется с утратой структур. Хорошие примеры этого явления — общая редукция экзоскелета во многих родословных линиях и утрата тяжёлой брони у силурийских и девонских рыб.[80] Но нам пока нет необходимости возвращаться во времени так далеко. Утрата или редукция чешуй у других рыб не представляет собой ничего необычного. Например, у осетров и веслоносов (отряд Осетрообразные) полный чешуйный покров, зарегистрированный у образцов из позднего мела, редуцировался до пяти рядов щитков уныне живущих видов{7}. Утрата чешуй независимо происходила в процессе эволюции, по крайней мере, тринадцать раз у гольянов и карпов (карпообразные) и несколько раз у галаксиевых, саланксовых (корюшкообразные) и икромечущих карпозубых (карпозубообразные). В пределах некоторых видов возможна значительная изменчивость. Одомашненный карп Cyprinus carpio существует в виде различных разновидностей: кожистый карп с фактически отсутствующей чешуёй; зеркальный карп с немногочисленными увеличенными чешуями и зеркальный линейный карп с единственным рядом крупных чешуй, тянущимся вдоль боков. Эти примеры изменчивости и эволюционных изменений вызвали интерес к расшифровке генетических механизмов, лежащих в основе явления редукции чешуи.[81] Известны сотни образцов Saurichthys, и изучение некоторых из них привело к открытию живорождения у этого животного. Я всегда думал, что когда-нибудь я бы хотел изучить эту рыбу и поработать с ней, но это случилось довольно неожиданным образом. Генетика развития и любезный и вежливый отставной доктор медицины из Базеля Лиени Шмид являются ключевыми фигурами в этой истории. В последние годы Лиени Шмид присоединился к Рудольфу Стокару и его коллегам в Лугано, собирая Saurichthys в триасовых породах в южной Швейцарии. Он также посещал курсы эво-дево, на которых обсуждались самые новые работы, связывающие морфологическую эволюцию с генетикой развития. Среди них были исследования некоторых видов рыб, которые хорошо известны с точки зрения генетики и служат моделями для понимания эволюционных изменений. Шмид заметил сходство в характере морфологических изменений, представленных в исследованиях генетики развития видов рыб, с изменчивостью у Saurichthys.
Рисунок 35. Схемы, показывающие разнообразие картины чешуйного покрова у Saurichthys. Последний общий предок должен представлять собой рыбу, полностью покрытую однородной ромбической чешуёй. По Schmid и Sánchez-Villagra 2010, с изменениями.
Он рассказал мне о своих идеях, и мы исследовали вопрос глубже и написали об этом работу, которую я вкратце излагаю здесь. Серия изящных исследований, которые включены в учебники, иллюстрирует генетические и фенотипические изменения на популяционном уровне в экологическом контексте. Лучше всего известный пример — это поразительная изменчивость в строении чешуйчатого покрова у колюшек.[82] У современных видов был зарегистрирован быстрый рост морфологического разнообразия, порождённый изменением регуляторных функций и взаимодействием между генами, экспрессия которых происходила посредством изменяющихся экологических условий. Трёхиглая колюшка (Gasterosteus aculeatus) несёт на своих боках ряд из примерно 36 пластинок — видоизменённых чешуй. В пресной воде последовательно отбиралась мутация эктодисплазинового сигнального пути (совокупности взаимодействий на молекулярном уровне), которая уменьшала количество этих видоизменённых чешуй с 36 до 0–7 в гомозиготном состоянии и до промежуточного количества в гетерозиготном. Молекулярные генетические факторы, ведущие к утрате чешуйчатого покрова, были изучены в лаборатории и в полевых условиях на колюшках, а также на мутантах нескольких видов рыб.
Рисунок 36. Образец Saurichthys curionii из триаса Монте-Сан-Джорджио, Швейцария (Палеонтологический институт и музей, Цюрихский Университет).
Существуют другие мутанты по эктодисплазиновому сигнальному пути. На примере японской медаки (Oryzias latipes) было продемонстрировано, что мутант с утратой чешуи возник из-за мутации в рецепторе этого пути. У данио-рерио (Danio rerio) было идентифицировано несколько мутаций, как в сигнальной молекуле самого эктодисплазина, так и в рецепторе, которые приводили к появлению фенотипов со сходными типами утраты чешуи. Другой молекулярный механизм, приводящий к утрате чешуи, был обнаружен у мутанта данио-рерио и у домашних карпов. Причиной этого является мутация утраты функционирования в рецепторе-1 фактора роста фибробластов. Известна важность факторов роста фибробластов на всех стадиях развития. Жизнеспособный мутант с мутацией этого пути возможен только благодаря дублированию гена, поскольку летальные побочные эффекты мутации в одном гене компенсируются немутированной версией этого же гена, которая в терминологии генетиков носит название «паралог». Мы с Шмидом постулировали, что подобные генетические процессы лежат в основе фенотипических изменений у Saurichthys и у мутантов этих рыб. Молекулярные основы морфологического разнообразия Saurichthys включали мутацию или регуляторные изменения сигнального пути в качестве ключевого механизма. Существуют серьёзные основания полагать, что эти механизмы действовали у триасовых рыб так же, как в наши дни. Анализ последовательности генов, кодирующих компоненты эктодисплазинового сигнального пути, показывает, что эти гены у позвоночных очень консервативны. Известно, что эктодисплазин контролирует формирование зубов и дермальных костей у челюстноротых позвоночных, чешуи у рыб, перьев у птиц и волос и кожных желёз у млекопитающих. Что же касается факторов роста фибробластов, то их работа существенна на очень ранних стадиях развития и они явно унаследованы по меньшей мере всеми позвоночными животными. Эти сигнальные пути представляют собой пример того, что названо «инструментальными белками»{8} и хорошо сохраняется в ходе эволюции. В дополнение к редукции количества чешуй, вышеупомянутые виды рыб благодаря мутации в эктодисплазиновом пути или в пути фактора роста фибробластов демонстрируют тенденцию к выстраиванию оставшихся чешуй в виде рисунка, схожего с тем, который наблюдается у отдельных рядов чешуй у Saurichthys. Кроме того, сохраняющиеся чешуи на боках мутантов вытянуты в дорсовентральном направлении так, что их можно сравнить с удлинёнными боковыми чешуями, как у видов Saurichthys с реброобразными боковыми чешуями. Мутации в компонентах эктодисплазинового пути также ведут, по крайней мере, у некоторых мутантов данио-рерио, к сокращению количества и снижению сложности строения плавниковых лучей, как было отмечено для некоторых морфологических типов Saurichthys. Кроме того, были сообщения об утрате зубов и изменениях размеров дермальных костей у мутантов данио-рерио, опять-таки изменения органов, которые менялись в процессе роста видового многообразия у Saurichthys.
Рисунок 37. Схематичные изображения рыб дикого типа и мутантов различных модельных видов, используемых в исследованиях в области генетики развития. Эктодисплазиновый путь: данио-рерио и японская медака, по Harris и др. 2008; трёхиглая колюшка, по Colosimo и др. 2005. Путь фактора роста фибробластов, по Rohner и др. 2009. Рисунки сделаны Л. Шмидом (Базель), из Schmid и Sánchez-Villagra 2010, с изменениями.
Особенности строения, контролируемые инструментальными белками, в том числе чешуйчатый покров, строение плавниковых лучей и особенности зубной системы, являются особенно важными для целей самозащиты, хищничества и питания, и таким образом для адаптации в целом. Изменение в важном гене, определяющем ход развития, даёт хороший шанс для симпатрического видообразования. Чем больше будет фенотипический эффект единственной мутации, тем больше шанс немедленной репродуктивной изоляции мутантов, если они окажутся жизнеспособными и успешными. Изменения в сигнальных путях, наличие которых мы предполагаем в случае увеличения видового многообразия у Saurichthys, могут, таким образом, дать объяснение явлению очень быстрого видообразования. В противоположность градуалистической модели эволюции, изменение в важном гене, контролирующем развитие, представляет собой реалистичное объяснение существенных различий между видами рода Saurichthys в палеоэкологическом контексте, в котором они возникли. Тот факт, что не были обнаружены промежуточные, плавно отличающиеся друг от друга ископаемые формы Saurichthys и большинства других видов, хорошо вписывается в общую картину.Глава восьмая «Недостающие звенья» и эволюция индивидуального развития
Многие люди привыкли думать об эволюции жизни как о прогрессии, похожей на лестницу, где каждое животное стоит на своей ступени. В случае эволюции позвоночных они могут вообразить рыбу на нижней ступени, саламандру на следующей, затем ящерицу, мышь, и, наконец, человека на верхней ступени. Следуя этому средневековому мифу, казалось вполне естественным предположить, что «низшие» животные эволюционируют в «высших» животных. И, если это так и было, мы должны рассчитывать увидеть «связующие звенья» между ними на всём пути вверх и вниз по «лестнице». Эта идея Великой Цепи Бытия, или Scala naturae, представляет собой образ эволюции, который всё ещё обычен в ненаучной среде, хотя учёные уже давно поняли, что такая концепция является неточной. Миф о «лестнице» связан с неверным пониманием эволюции и не соответствует реальному процессу. Вместо того, чтобы быть похожей на лестницу, эволюция жизни больше напоминает ветвящийся кустарник. Каждая ветка кустарника представляет собой самостоятельную родословную ветвь живых организмов. Места, где из одной точки на кустарнике вырастают две или больше ветвей, указывают на то, что родословные линии, представленные ветвями, в какой-то момент своей истории должны были иметь общего предка. Каждый из ныне живущих видов является «недостающим звеном», поскольку каждый вид является частью древа жизни и обладает мозаикой как «примитивных» пережитков прошлого, так и продвинутых особенностей, которые своим наличием подтверждают изменения. От каждого вида можно многое узнать об этих изменениях. Каждый из миллионов видов животных имеет общих предков с любым другим видом. «Недостающее звено» — это предковый вид, который дал начало двум современным группам, например, людям с одной стороны и шимпанзе с другой. Но было бы не разумно ожидать обнаружить именно тот критически важный вид; для его идентификации потребовался бы полный ряд ископаемых предков-потомков. Конечно, мы можем проследить эволюцию птиц из крылатых динозавров, но мы точно не уверены в том, какой именно ископаемый вид был прямым предком современных птиц. Наши усилия в деле реконструкции истории жизни не должны быть направлены на попытки вообразить себе, как шимпанзе мог превратиться в человека, поскольку этого, конечно, никогда не случалось. Мы скорее попытаемся установить, какие признаки шимпанзе и люди унаследовали от их общего предка, и какие признаки появились в их родословных линиях в процессе эволюции уже после того, как они разошлись. Некоторые авторы правильно говорят, что не только концепция «недостающего звена» архаична, но также его поиск — это устаревший подход к исследованию макроэволюции. На нынешней стадии развития эволюционной биологии фокус в исследованиях переместился с обнаружения переходных таксонов на обнаружение переходных особенностей, которые являются общими для близкородственных форм с общим происхождением. В наше время хорошо известно много примеров такого рода, и они снабжают нас информацией не только об анатомических преобразованиях, но также и о функциональном и экологическом контекстах этих переходов. В этом контексте особенно важны стволовые{9} виды, те, которые свидетельствуют о времени и способе приобретения диагностических признаков ныне живущих видов в ходе эволюции.[83] Пример такого рода даёт нам сама наша родословная ветвь млекопитающих. Согласно молекулярным оценкам, подтверждаемым свидетельствами со стороны ископаемых остатков, последний общий предок всех ныне живущих млекопитающих, то есть последний общий предок человека (плацентарного млекопитающего), кенгуру (сумчатого) и утконоса (однопроходного) жил, по крайней мере, 220 миллионов лет назад. Но история происхождения млекопитающих от нашего последнего общего с рептилиями предка уходит в глубины времени ещё дальше, до отметки, по меньшей мере, в 315 миллионов лет назад. Эти самые ранние 100 миллионов лет эволюционной истории подтверждаются стволовыми видами млекопитающих. Они стоят на эволюционном древе между последним общим предком ныне живущих видов и последним общим предком с ближайшими ныне живущими родственниками, в данном случае с рептилиями. Для нескольких крупных таксонов ископаемые остатки предоставляют довольно полную картину морфологических преобразований, связанных с их происхождением. Летопись окаменелостей многих групп изменилась с почти отсутствующей на довольно разнообразную в течение лишь немногих десятилетий, как в случае с мезозойскими млекопитающими. Непрерывные сообщения о новых открытиях «переходных форм» показывают большой шаг вперёд, который был сделан и делается сейчас в области документации макроэволюционных шагов позвоночных со времён Дарвина. Эти сообщения также показывают, что для органической эволюции не характерны цель и план. Организмы выполняют свои «условия для поддержания жизни» в каждый конкретный момент времени, в котором они живут — и важно именно это, о чём я и буду говорить далее.Глаза камбалы
Среди огромного множества современных рыб, примерно тридцати тысяч видов, существует много форм с признаками анатомической специализации, происхождение которых в большинстве случаев не зарегистрировано в ископаемых остатках. Классическим примером была группа камбал, к которой принадлежат знакомые нам с гастрономической (жаль, что в основном так) палтус и камбала, необычные тем, что оба их глаза расположены на одной стороне головы. Мэтт Фридман из Оксфордского Университета описал ископаемые остатки из эоценовых пород Италии и Франции возрастом около 50 миллионов лет, которые показывают расположение глаз, промежуточное между таковым у нормальных рыб и камбал: один глаз смещён к противоположной стороне черепа, но сделал это не совсем в полной мере. Открытие предковых камбал с неполным смещением глазниц демонстрирует нам то, что происхождение плана строения тела камбалы возникло постепенным, пошаговым путём. Это эволюционное происхождение напоминает индивидуальное развитие асимметрии у камбалы, когда увеличение степени смещения глазницы преобразует симметричного эмбриона в полностью асимметричную взрослую особь. «Промежуточные» окаменелости представляли собой взрослых особей, а не молодь, поэтому, фактически, не было зарегистрировано никакого «онтогенеза в ископаемом виде». Но отдельные окаменелости позволяют сделать красивое сравнение с данными об индивидуальном развитии, полученными для этих животных. Как эти промежуточные формы использовали наполовину смещённые глаза, остаётся загадкой.
Рисунок 38. Heteronectes chaneti из нижнего эоцена Монте-Болька, Италия. На одной стороне (внизу) глазница находится в своём обычном положении, но на другой она расположена намного дорсальнее (сверху), вблизи средней линии головы. Фотографию любезно предоставил Мэтт Фридман (Оксфорд).
Случай с камбалами имеет отношение к одной из наиболее фундаментальных дискуссий в эволюционной биологии, корни которой уходят в такие влиятельные и спорные идеи, как «перспективный монстр». Этот термин был введён Ричардом Гольдшмидтом (1878–1958), который наряду с Гансом Шпеманом и Виктором Гамбургером сделал Германию выдающейся страной в области биологии развития. Он стал профессором зоологии в Калифорнийском Университете в Беркли в 1936 году. Гольдшмидт особо отметил отсутствие «промежуточных форм» в эволюции и предложил в качестве примера асимметричный череп камбалы. Идеи Гольдшмидта были в значительной степени упрощёнными и подвергались жёсткой критике со стороны людей, приводивших доводы в пользу градуалистической эволюции. Он стал в некотором роде карикатурным образом намертво застрявшего в ошибках учёного и остаётся изображённым так в некоторых упрощённых обзорах дебатов в области эволюционной биологии.
Рисунок 39. Перемещение глаза в ходе индивидуального развития камбалы.
Фактически, Гольдшмидт подчеркнул роль индивидуального развития в происхождении его «перспективных монстров», утверждая, что они не просто появлялись из небытия как результат генетической мутации. Его акцент был сделан не просто на генах, а на эпигенетических системах, возникающих в результате комбинации генетических факторов и факторов окружающей среды. Из злодея, каким он был, Гольдшмидт теперь стал чем-то вроде героя.[84] Ископаемые остатки камбал, казалось бы, являются доводом против идеи Гольдшмидта. Они, конечно, противоречат его предсказанию, касающемуся этих рыб, и относятся к обсуждаемой проблеме характера и скорости эволюции. Но идеи Гольдшмидта, однако, важны, и в иных случаях эмпирические свидетельства поддерживают их. Кроме того, хотя мы пытаемся вывести общие принципы, каждая отдельно взятая эволюционная история уникальна, поэтому в обобщениях стоит проявлять осторожность. Этот пример с камбалой, с переходными формами, о которых бы Гольдшмидт и не подумал, что они могли существовать, не является залогом верности градуализма; это лишь пример того, как летопись окаменелостей в некоторых случаях являются единственным способом проверить эволюционную гипотезу непосредственно. Случай с летучей мышью является примером противоположного рода — это случай, когда именно молчание летописи окаменелостей может показать истинную картину эволюции.Крылья летучей мыши
Многое в анатомии летучих мышей является специфичным — например, специализация ушей у многих видов, которая позволяет им использовать эхолокацию, способность слышать звуки очень высокой частоты, чтобы определять расположение добычи, врагов и объектов в пространстве. Ископаемые остатки из эоцена Северной Америки и Европы показывают, что эхолокация появилась на ранних этапах эволюции летучих мышей, хотя уже после полёта. Специализации к слуху в области высокочастотных звуков у летучих мышей, вероятно, возникали в процессе эволюции более одного раза в пределах этой группы. Своего рода эхолокацию также используют землеройки, а иным способом{10} и в другой среде — некоторые киты. Но действительно исключительный аспект летучих мышей — это их крылья, которые позволяют им использовать активный полёт — признак, который среди прочих позвоночных появился в ходе эволюции только у птиц и (до позднего мела) у вымерших птеродактилей. Крыло летучих мышей уникально тем, что оно состоит главным образом из очень удлинённых пальцев, кроме большого пальца, и подвижной кожи между ними. Как могли бы выглядеть передние конечности «недостающего звена»? Среди ныне живущих видов много разных млекопитающих способно к планирующему полёту и для этой цели у них имеется кожа между конечностями. Планирующий полёт выглядит первым шагом к полёту, включающим движение по воздуху с использованием складки кожи. Но различия всё же огромны. Способность постоянно махать передними конечностями с очень большими пальцами — это нечто совсем другое. Исчерпывающие молекулярные исследования ныне живущих млекопитающих ясно указывают на то, что летучие мыши являются самыми близкими родственниками копытных, вроде коров и лошадей, и вместе с ними образуют одну группу с хищными млекопитающими, такими, как собаки и кошки{11}. Вопрос о родственных связях летучих мышей с современными млекопитающими, таким образом, решён, но вопрос о происхождении уникальнейшего морфологического новшества летучих мышей, активного полёта, всё ещё остаётся без ответа. Приблизительно через десять миллионов лет после того, как вымерли динозавры, не относящиеся к птицам, адаптивная радиация существующих в наше время групп млекопитающих становится хорошо документированной ископаемыми остатками. Многие эоценовые окаменелости замечательно полны и вне всяких сомнений классифицируются среди самых ранних летучих мышей. У них уже имеются крылья, и, несмотря на крупномасштабные поиски в богатых отложениях хорошо сохранившихся окаменелостей, неизвестно никаких промежуточных звеньев. Возможно, это эволюционное новшество возникло очень быстро. Биология развития даёт подсказки в отношении механизмов, которые, вероятно, сделали это возможным. Передние конечности, на раннем этапе развития почки конечности относительно более крупные по сравнению с задними — это довольно обычная картина для млекопитающих. Так происходит в период индивидуального развития, который некоторые коллеги называют филотипическим и который характеризуется некоторой морфологической и, возможно, генетической общностью для различных таксонов. От этой в чём-то сходной стартовой точки пути развития летучих мышей и других млекопитающих начинают расходиться. Это было детально документировано для некоторых видов летучих мышей и для обычных мышей. У летучих мышей увеличение размеров пальцев на передней конечности происходит быстрее, чем пальцев на задней конечности — это пример аллометрического роста для конечностей. У обычных мышей и передние, и задние лапы растут со сходной скоростью. Удлинение пальцев летучей мыши в ходе развития достигается благодаря ускоренному темпу разрастания и дифференциации клеток хряща. Карен Сирс и её коллеги обнаружили, что этот процесс связан с более высоким содержанием костного морфогенетического белка (bone morphogenetic protein) Bmp2. Другая работа сравнительного характера показала также, какие молекулы задействованы в сохранении кожи между пальцами в ходе индивидуального развития, что приводит к образованию крыла у взрослой особи. На стадии раннего эмбриона у нас есть кожа между пальцами, как у всех позвоночных с руками и ногами. В некоторый момент времени клетки, образующие эту кожу, начинают отмирать, и поэтому у нас есть наши подвижные пальцы, а не весло или крыло, как лапы уток или крылья летучих мышей. На лапах утки и в передней конечности летучей мыши кожа не отмирает, а сохраняется и даже разрастается. Механизмы, задействованные в этих двух случаях, аналогичны, но, как открыли Скоттом Ветерби и его коллеги, участвующие в них молекулы не одинаковы. Было известно, что у цыплят и мышей, модельных организмов для исследований в области генетики развития, костные морфогенетические белки (Bmp), которые способны стимулировать рост и дифференциацию клеток, запускают процесс гибели клеток в недифференцированной, свободной соединительной ткани (мезенхиме) между развивающимися пальцами «весла» передней и задней конечностей, характерного для эмбрионального состояния. В ноге утки молекула Gremlin служит ингибитором этого действия Bmp, и её работа приводит к образованию перепончатой лапы. Летучая мышь также обладает этим механизмом, но в нём дополнительно участвует другой белок, а именно, фактор роста фибробластов Fgf8, также играющий роль в различных аспектах морфогенеза.
Рисунок 40. Эмбрион крылана Rousettus amplexicaudatus, изображение получено с помощью сканирующего электронного микроскопа. Фотография М. Санчеса, из Giannini, Goswami и Sánchez-Villagra, 2006, с изменениями.
Важность этих открытий состоит в том, что они предоставляют нам эмпирическое свидетельство существования простого механизма, задействованного в процессе образования крыльев — новшества, возникающего в ходе индивидуального развития. Теперь «внезапное» возникновение летучих мышей выглядит менее маловероятным, и свидетельства со стороны окаменелостей и их отсутствие могут указывать именно на такую ситуацию. Существуют ли какие-то ископаемые эмбрионы летучих мышей, которые предоставляют какое-либо дополнительное свидетельство? Единственный, насколько я знаю, происходит из эоценового местонахождения Мессель в Германии, известного, помимо всего прочего, одними из самых ранних и наилучшим образом сохранившимися ископаемыми остатками летучих мышей. Скелет ископаемого эмбриона уже сформирован, а крылья более или менее видны.Панцирь черепах
Другой специфичный план строения тела среди позвоночных, на сей раз среди рептилий, — это план строения тела черепах. Главные признаки специализации скелета — это наличие панциря, отсутствие зубов, короткий хвост и анапсидный тип строения черепа. Черепа у черепах лишены окон для прикрепления мускулов, типичных для других наземных позвоночных. Это состояние может являться как вторичным, так и первичным; мы не можем сказать об этом главным образом из-за того, что мы не уверены в том, где размещать черепах на древе жизни рептилий.[85] Если они находятся в основании древа, то их черепа, вероятнее всего, являются просто результатом сохранения примитивного состояния, поскольку самые ранние рептилии, хорошо представленные в летописи окаменелостей, являются анапсидами. Если же черепахи являются сестринской группой по отношению или к ящерицам, или к крокодилам, то тогда их черепа в значительной степени специализированы. Пока известно недостаточно ископаемых остатков для решения этого вопроса, даже несмотря на некоторые великолепные образцы, которые были описаны недавно. Возможно, помогут будущие находки окаменелостей. В 2008 году Ли и коллеги опубликовали описание нового рода и вида Odontochelys semitestacea из отложений Китая возрастом около 220 миллионов лет. Это животное имело анапсидный череп, но в остальном оно демонстрировало много переходных особенностей, вроде наличия зубов и длинного хвоста.[86] Но, что ещё важнее, пример Odontochelys касается того, что уникально для черепах, их панциря. Черепаший панцирь состоит из дорсальной, или спинной части, вентральной, или брюшной части, и костных мостиков, соединяющих их{12}. Вентральная часть состоит из расширенных брюшных рёбер, слившихся с некоторыми костями исходного для позвоночных плечевого пояса, такими, как ключицы, которые теперь сильно видоизменены. Спинная часть панциря намного сложнее по своему происхождению, поскольку она является результатом слияния рёбер с кожными окостенениями, которые аналогичны чешуям или остеодермам многих рептилий. Наиболее примечательно то, что у черепах лопатка находится под спинным панцирем, или карапаксом; это уникальное состояние: у всех прочих рептилий и у млекопитающих, например, как у людей, лопатка находится снаружи грудной клетки. Наличие панциря и его анатомические связи у черепах настолько уникальны, что для объяснения его происхождения традиционно обращались к идее скачкообразных эволюционных изменений. Желая добиться понимания развития такой странной анатомии, Хироши Нагашима и его коллеги изучили, как формируется панцирь по отношению к лопатке и к мускулатуре тела. Их анатомическое исследование показало, что новизна в строении панциря черепахи — это то, что мои коллеги назвали осевой задержкой роста ребра, и складывание мускулов для подгонки под необычное положение плечевого пояса. Исследование индивидуального развития проводилось не с наивным ожиданием проявления рекапитуляции, а скорее с целью понимания топографического положения мускулов у современных черепах и их развития. Их работа дала ключи для формулировки вероятной гипотезы морфологического преобразования, которую можно проверить на окаменелостях. Odontochelys, самая древняя ископаемая черепаха, является «недостающим звеном» для понимания происхождения панциря, поскольку у неё имеется пол-панциря — хорошо различимая нижняя половина. На спинной стороне широкие и короткие рёбра Odontochelys напоминают рёбра позднего эмбриона ныне живущих черепах. Фактически, анатомия Odontochelys очень хорошо соответствует модели, которую представили Нагашима и его коллеги: рёбра укорочены настолько, что без панциря лопатка расположена не выше и не в дорсальном положении по отношению к рёбрам, но вместо этого находится перед первой парой рёбер.
Рисунок 41. Карапакс черепах состоит из расширенных рёбер и позвоночника, слившихся со скелетными тканями, происходящими из кожи. План строения тела очень отличается от других наземных позвоночных, поскольку у черепах лопатка расположена внутри грудной клетки. Хироши Нагашима и его коллеги предполагают, что у Odontochelys semitestacea из позднего триаса Гуйчжоу (Китай) эмбриональный «гребень карапакса» (пунктирная линия), возможно, развивался у эмбриона лишь временно и не полностью, стимулируя частичное веерообразное разрастание рёбер. Уникальное расположение лопатки в ходе индивидуального развитии черепахи приводит к образованию складки на мускульной части стенки тела в области плеча.
Пример с черепахами иллюстрирует три фундаментальных положения: Ископаемые остатки являются важнейшим свидетельством, показывающим, в каком порядке возникали новшества процесса развития: брюшной панцирь раньше спинного панциря. Вместе с данными эмбрионального развития ископаемые остатки представляют нам хорошо обоснованные сценарии процесса возникновения эволюционных новшеств. Ископаемые остатки могут быть очень информативными в отношении некоторых особенностей и ничего не говорить о других. В случае с черепахами мы многое узнали о панцире, но гораздо меньше о черепе. Другой аспект, который следует рассмотреть на примере черепахи и сальтационного сценария происхождения черепашьего плана строения тела — это объяснение одновременного «скачка», который приводит к появлению в значительной степени интегрированного набора особенностей, с которыми связан панцирь; среди них мускулатура и её управление с помощью нервов, анатомия конечностей и механизмы вентиляции лёгких. Здесь более вероятным был бы такой сценарий, который делает возможным относительно быстрое изменение, но также и учитывает потенциальную корреляцию между различными особенностями строения черепахи.Ноги китов
До 1980-х годов происхождение китов было изучено плохо. На основе лишь одних анатомических исследований было трудно разобраться в том, какая группа наземных млекопитающих была самой близкой к ним. Ископаемые киты были известны с девятнадцатого века, но это не помогало разрешить эту загадку, поскольку они уже были животными, очень специализированными в плане физиологических требований к жизни в воде, и обладали плавниками и хвостовыми лопастями. После этого времени серия замечательных открытий окаменелостей, сделанных международной командой Фила Джинджерича и Ханса Тевиссена, дала нам довольно полный ряд окаменелостей, которые демонстрируют преобразования и документируют шаги, ведущие от обладающих нормально развитыми ногами наземных копытных к млекопитающему без задних конечностей, имеющему лишь остатки тазового пояса. Большинство этих окаменелостей происходит из эоцена Индийского субконтинента и из Египта. Параллельно палеонтологическим открытиям молекулярные исследования чётко определили место китов как самых близких родственников бегемота, и это означает, что китов следует классифицировать в рамках группы парнокопытных (Artiodactyla), к которой принадлежат коровы, жирафы, олени и свиньи. Летопись окаменелостей показывает, что адаптация китов и бегемотов к водному образу жизни возникла в процессе эволюции независимо. Пакицетиды — это наиболее примитивная группа ископаемых китов. Анатомия их уха занимает промежуточное положение между ушами наземных и полностью водных млекопитающих, что означает, что оно, вероятно, могло функционировать в обеих средах. Пакицетиды были способны бегать по суше, что отражено в анатомии скелета их конечностей и позвоночника.[87] Данные о химических анализах соотношения изотопов кислорода и исследования отложений, в которых были обнаружены пакицетиды, дают основание полагать, что они жили в реках или вблизи них, а не в морских местообитаниях. Следующее важное ископаемое животное в цепочке — это Ambulocetus, который вёл земноводный образ жизни. У него были длинные шея и хвост, а крупные ступни задних лап и другие особенности подсказывают, что это животное плавало, как выдра. Ремингтоноцетиды были экологически схожими с ныне живущими крокодилами гавиалами из Юго-Восточной Азии; они обладали длинными и узкими черепами и были способны совершать быстрые боковые движения для схватывания рыбы. Морфология ушной области указывает на их способность к подводному слуху. Ещё несколько ископаемых форм, таких, как Rhodocetus и Dorudon, документируют дальнейшие морфологические преобразования вроде редукции задних ног.
Рисунок 42. Постепенная редукция костей задней конечности в ходе эволюции китообразных, с примерами из их основных групп. Из Thewissen и др. 2006, с изменениями.
Что же ископаемые остатки китов рассказывают нам об эволюции индивидуального развития? Крупные эволюционные преобразования, такие, как совершенно новый план строения тела, приспособленный к жизни в новой среде обитания, являются особенно подходящим объектом исследований в области индивидуального развития, потому что новшества, которые мы видим у полностью сформированных взрослых особей — это результат процесса онтогенеза, который должен был в значительной степени измениться с течением геологического времени. Ископаемые остатки показывают темп и путь образования новых особенностей. Последовательность их возникновения особенно важна. Ханс Тевиссен и его коллеги исследовали некоторые аспекты специализированной генетической системы развития конечностей у видов дельфинов, результатом действия которой является почти полная редукция задних конечностей. У большинства Odontoceti, или зубатых китов, к которым принадлежат дельфины, единственный элемент задней конечности — безымянная кость, тогда как у усатых китов (Mysticeti) в целом сохраняются некоторые из самых проксимальных костей задней конечности. Исследование эмбрионов дельфинов показало, что многие из молекул, задействованных в развитии конечностей у других млекопитающих, также экспрессируются у дельфинов, но в иное время и в другом местоположении. Например, апикальный эктодермальный гребень, дистальный край почки конечности, где происходит экспрессия гена fgf8, у дельфина является лишь временной особенностью строения. Другая важная молекула, sonic hedgehog, которая является посредником в развитии основной части почки конечности у других наземных позвоночных, здесь отсутствует. Тевиссен и его сотрудники выдвинули гипотезу о том, что это отсутствие связано с утратой дистальных элементов конечности, таких, как ступня. Зафиксированная последовательность реально происходившего эволюционного изменения, которую предоставляют нам ископаемые остатки, и знание генетических основ индивидуального развития, задействованных в процессе редукции задней конечности, предлагают подробный сценарий изменений. Не имея ископаемых остатков, мы могли бы лишь предполагать, на что были похожи эти изменения. Но благодаря ископаемым остаткам могут быть реконструированы потенциальные генетические механизмы, задействованные в этом процессе.Глава девятая Развитие у млекопитающих и у человека
Существует около 5300 видов современных млекопитающих. Они представляют собой лишь небольшую часть от общего количества видов, когда-либо существовавших начиная с обособления ведущей к ним эволюционной линии как минимум 315 миллионов лет назад. Затем родословные ветви рептилий и млекопитающих разделились. По оценкам, сделанным на основе ископаемых остатков, прошло ещё около 100 миллионов лет, когда в юрский период появился последний общий предок всех ныне живущих млекопитающих.[88] Хотя цифра в 5300 современных видов выглядит внушительно, она не столь уж внушительна по сравнению с другими группами: существует вдвое больше видов птиц и в пять раз больше видов костистых рыб. Но млекопитающие выглядят весьма особенными, когда мы рассматриваем их значительное экоморфологическое разнообразие — способы локомоции у людей, кенгуру, китов и летучих мышей; размер, варьирующий от землеройки до синего кита. Что у всех млекопитающих есть общего — это уникальные для позвоночных история жизни и физиология, в том числе энергозатратная эндотермия, родительская забота, выделение молока и детерминированный рост. Эти и многие другие особенности должны были характеризовать последнего общего предка ныне живущих млекопитающих. Когда же, начиная с момента отделения от рептилий, возникли эти особенности? Появились ли они одновременно? Если нет, то в какой последовательности? Неважно, как обстояли дела у наших предков, но во время двух критических отрезков геологической истории их выручали удача и хорошие гены. Я уже говорил выше, как Lystrosaurus и его родственники преодолели границу перми и триаса, время, когда множество групп организмов было полностью стёрто с лица земли. Млекопитающие также пережили событие конца мелового периода, когда вымерло большинство{13} динозавров.
Рисунок 43. После дивергенции с рептилиями не позднее 315 миллионов лет назад началась долгая история «предковых млекопитающих» — вплоть до появления по крайней мере 220 миллионов лет назад последнего общего предка современных видов. Ныне живущих млекопитающих представляют ехидна (однопроходное), кенгуру (сумчатое) и человек (плацентарное). Предковые виды, изображённые здесь — это все ключевые виды, упомянутые в тексте, но они — лишь крохотная часть общего многообразия параллельных эволюционных линий, которые развивались на раннем этапе эволюции млекопитающих.
Несколько уникальных анатомических особенностей млекопитающих не связаны непосредственно с особенностями жизненной истории, например, наличие трёх небольших косточек уха в противоположность только одной у рептилий и птиц. Способы, которыми эмбриологические и палеонтологические данные документируют происхождение слуховых косточек млекопитающих, прекрасно соотносятся друг с другом, и их открытие является одним из величайших триумфов сравнительной анатомии. Как я уже рассказывал в восьмой главе, Гольдшмидт сделал упор на отсутствие промежуточных форм в эволюции и важность эпигенетики, и, в дополнение к глазам камбалы, среднее ухо млекопитающих было его основным примером. Оба они оказались плохими примерами в остальном хорошей идеи. Фактически, Гольдшмидт должен бы приветствовать новейшее свидетельство того, что происхождение среднего уха млекопитающих было связано в процессе развития с увеличением размера мозга и, вероятнее всего, со многими другими особенностями, в том числе с изменениями в жевательном аппарате. Сопутствующие друг другу изменения в процессе развития показывают, что эти особенности не могут быть поняты отдельно друг от друга. В ходе онтогенеза современных млекопитающих рост неокортекса связан с постепенным отделением элементов, связанных с челюстями. Некоторые из старых элементов челюсти становятся относительно меньше и меньше по мере того, как они обособляются и начинают формировать будущие элементы среднего уха — это пример отрицательного аллометрического роста. Постепенные анатомические изменения, сходные с изменениями у эмбрионов — это то, что ископаемые остатки документируют в основной линии, ведущей к современным млекопитающим. Эти изменения зафиксировал Тим Роу из Техасского Университета на основании изучения полученных методом компьютерной томографии изображений окаменелостей и реконструкции развития эмбрионов сумчатых млекопитающих современных видов.[89] Наличие взаимосвязи, или отсутствие независимости в ходе эволюционного возникновения млекопитающих также характеризует особенности истории жизни. Документация этих эволюционных изменений ограничена скелетом, поскольку мягкие ткани, включая органы вроде сердца или желудка сохраняются редко, если сохраняются вообще. Не удивительно, что большинство свидетельств получено при изучении зубов и гистологии костей. Сложная картина зубной окклюзии у млекопитающих, когда верхние и нижние коренные зубы, обладающие несколькими буграми, смыкаются друг с другом особым образом, несомненно, коррелирует с высокой скоростью обмена веществ, который, в свою очередь, связан с особенностями истории жизни. Когда большинство рептилий, рацион которых включает других животных, просто ловит свою добычу, и, самое большее, режет её зубами перед тем, как проглотить, млекопитающие имеют тенденцию жевать то, что едят. Выделения слюнных желёз смешиваются с пищей, таким образом частично обрабатывая её во рту перед тем, как она будет переварена в желудке и кишечнике. Развитие молочного вскармливания у млекопитающих связано с особенностями зубов, такими, как позднее прорезание первых функциональных зубов. У млекопитающих образуется только две смены зубов — это состояние, называемое «дифиодонтия» и включающее наборы зубов молодой и взрослой особи. Кормление молоком, вероятнее всего, возникло, когда характер смены зубов стал дифиодонтным.[90] С этим, в свою очередь, был связан детерминированный рост — это означает, что быстрое развитие ювенильной особи завершается по достижении взрослого размера. У рептилий, напротив, рост главным образом недетерминированный, и это означает, что крокодил, например, никогда не останавливается в росте и у него никогда не прекращается замена зубов, хотя скорость роста может замедлиться с возрастом. Каждый из существующих в наше время видов млекопитающих имеет определённую зубную формулу, означающую определённое число резцов, клыков, предкоренных и коренных зубов. Эта дифференциация зубной системы специфична для млекопитающих, как и тот факт, что самая задняя группа зубов, коренные зубы, не сменяется. Ископаемые остаткидокументируют момент, когда установились эта дифференциация зубов и характер их смены. Серия образцов, демонстрирующих процесс роста триасового «стволового» млекопитающего Thrinaxodon{14}, показывает, что у этого животного было восемь заклыковых зубов на одной из ювенильных стадий, и ещё четыре добавлялось сзади по мере продолжения роста. Sinoconodon, который жил в позднем триасе и считается, как и Morganucodon, одним из «первых млекопитающих», явно обладал недетерминированным ростом и многократно сменявшимися резцами. У мелового млекопитающего Gobiconodon «коренные зубы» сменялись. Благодаря комбинированию данных о характере прорезывания зубов и палеогистологических данных о росте становится возможным проследить происхождение особенностей истории жизни, характерных для млекопитающих. В палеогистологических исследованиях важным моментом является выявление наличия фиброламеллярной кости, поскольку она указывает на быстрый остеогенез и вместе с тем быстрый общий рост, связанный также с эндотермией (см. главу 4). Этот вид кости обнаружен у многих, хотя и не у самых ранних члены родословной ветви млекопитающих. Самые ранние предки млекопитающих по физиологии и по особенностям индивидуального развития были больше похожи на современных рептилий, чем на млекопитающих. Пример такого рода — Dimetrodon, который обладал недетерминированым ростом, характерным для ранних амниот, и особенности его гистологии показывают отсутствие фиброламеллярной кости. Помимо прямого указания со стороны гистологии костей, существует косвенный способ, позволяющий выявить детерминированный рост — путём популяционных исследований ископаемых форм при возможности хорошего стратиграфического контроля. Он требует большого количества окаменелостей одного и того же вида из одного и того же слоя отложений. Если в выборке все взрослые особи обладают одинаковым размером, это дало бы основание предполагать наличие детерминированного роста. Как раз это отмечалось по итогам популяционного исследования Morganucodon из позднего триаса — ранней юры, которого многие палеонтологи рассматривают как одно из «первых млекопитающих» из-за набора анатомических особенностей, присутствующих у этого животного. У млекопитающих размер — это в целом хорошее внешнее проявление возраста неполовозрелых особей, поскольку факторы окружающей среды вроде температуры оказывают не настолько сильное воздействие на их размер, как это бывает у рептилий. В отличие от Morganucodon, у Dimetrodon, а также у его близкого родственника Ophiacodon, взрослые животные одного и того же вида имеют различные размеры.[91] У этих самых ранних членов нашей родословной линии, как у рептилий, размер является ненадёжным признаком стадии развития. Поскольку информация об анатомии подтверждает принадлежность особей различных размеров к одному и тому же виду, это наблюдение подразумевает то, что эти самые ранние представители родословной линии млекопитающих не обладали характерным для млекопитающих способом роста и потому для них размер не является хорошим внешним признаком возраста. На время развития эмбрионов у рыб, амфибий и рептилий оказывает значительное воздействие температура, при которой происходит их инкубация, и у многих видов ею определяется даже пол. В зависимости от температуры, при которой происходила инкубация у черепахи, и которая оказывала значительное влияние на темп обмена веществ и тем самым на рост, размеры на такого рода «стадиях» развития могут быть весьма различными. Млекопитающие, в свою очередь, характеризуются более стабильной внутренней температурной средой, так называемой гомеостатической способностью, достигающейся путём поддержания высокой скорости обмена веществ в постоянной внутренней среде. Для первых юрских млекопитающих была характерна меньшая пластичность индивидуального развития, с более стабильной картиной роста по сравнению с часто регистрируемыми случаями ускоренного и замедленного роста, типичными для рептилий и самых ранних представителей ветви млекопитающих. Способность приостанавливать или замедлять рост при неблагоприятных условиях окружающей среды должна была характеризовать, например, Dimetrodon, что подтверждается палеогистологическими данными. Том Кемп из Оксфордского Университета назвал интегрированную эволюцию признаков, описанных выше, «коррелированной прогрессией». Он думает, что эта плавная и почти синергическая картина эволюции, напоминающая сеть взаимоотношений, меняющихся со временем, характеризует не только многое в ранней эволюции млекопитающих, но также происхождение других крупных групп, таких, как черепахи и первые позвоночные с конечностями, или четвероногие. Идея во многом выглядит привлекательной. Конечно, координация изменений в различных частях тела, вероятно, вызвана скорее последовательностью небольших, нарастающих изменений, чем одним единственным шагом. Согласно Кемпу, изменения признаков «эволюционируют аналогично цепочке людей, идущих вперёд, взявшись за руки: любой из них может двигаться в своём собственном темпе, впереди или позади тех, кто находится рядом, но лишь разорвав цепочку». Экспериментальные свидетельства, полученные для таких разных организмов, как лабораторные мыши, растения, насекомые, черепахи и даже люди, служат напоминанием того, что признаки не эволюционируют сами по себе; они являются частью целого организма. Известны эксперименты, в которых исследователи пробовали оценить степень, в которой изменение формы определяется генетической основой. В ходе управляемого размножения или при исследовании информации о чистокровном разведении исследователи изучают эффект селекционных экспериментов, связанных с определённой особенностью. В этих случаях обычно имеет место ответ на отбор со стороны иных аспектов облика сверх того, на который был изначально направлен отбор.
Рисунок 44. Иллюстрация взаимосвязей между несколькими признаками, характерными для млекопитающих, которые, несомненно, эволюционировали параллельно и по принципу «коррелированной прогрессии».
Что касается происхождения картины истории жизни млекопитающего, было бы неправильно предполагать, что первостепенным был какой-то один конкретный меняющийся признак, поскольку они все интегрированы. Изменение в одной переменной величине должно было сопровождаться изменениями в других переменных организма, чтобы они могли функционировать, соответствовать «условиям для поддержания жизни». Рисунок 44 изображает эту идею о сети связей. Картина корреляции и взаимодействий в эволюции, представленная на рисунке, имеет в своей основе эмпирические свидетельства. По мере того, как летопись ранней эволюции родословной линии млекопитающих всё лучше наполняется находками, а исследования истории жизни становятся более определёнными в количественном выражении, у нас появится возможность выверить и дополнить деталями эту картину. Умная идея о коррелированной прогрессии остаётся лишь идеей, если она не подкреплена эмпирическими свидетельствами, а единственный источник прямых свидетельств — это летопись окаменелостей. История жизни и физиология млекопитающих коррелятивно связаны с несколькими анатомическими особенностями, и некоторые из них зарегистрированы на ископаемых образцах. Они показывают нам, что диагностические особенности млекопитающих не возникали одновременно, а некоторые из них эволюционировали конвергентно в линиях, вымерших в дальнейшем. Пример особенности строения, которая независимо возникала в процессе эволюции несколько раз — вторичное нёбо. Вторичное нёбо — это состоящая из кости и мягкой ткани горизонтальная перегородка, которой мы можем коснуться языком, когда он движется в верхней части ротовой полости. Эта особенность, разделение между ротовой и носовой полостями, позволяет нам, млекопитающим, дышать, пока мы едим, и таким образом способствует более эффективной обработке пищи и выработке энергии для поддержания высокой скорости нашего обмена веществ.[92]Эволюция индивидуального развития на раннем этапе формирования многообразия современных млекопитающих
После 100 миллионов лет эволюции нашей родословной линии отдельно от линии рептилий, возник последний общий предок ныне живущих млекопитающих и установились диагностические признаки класса Mammalia по определению. Но эволюция млекопитающих включала дальнейшие существенные изменения, и были другие крупные события в эволюции истории жизни. Возможно, важнейшее из них затрагивало коренное разделение ныне живущих млекопитающих на однопроходных, включающих утконоса и ехидн как их единственных современных представителей, с одной стороны, и сумчатых и плацентарных с другой стороны. Однопроходные откладывают яйца, тогда как сумчатые и плацентарные этого не делают. Это различие — одно из многих других, которые совместно с летописью окаменелостей и оценками, основанными на молекулярных данных, демонстрируют глубокое разделение двух эволюционных линий, восходящее, по меньшей мере, к юрскому периоду. Существование однопроходных в нашем мире очень полезно, поскольку они позволяют нам изучить многое из того, на что были похожи самые ранние сумчатые и плацентарные,[93] хотя сами по себе и утконос, и ехидны являются высокоспециализированными формами, как показывает не только их анатомия, но также и их геном.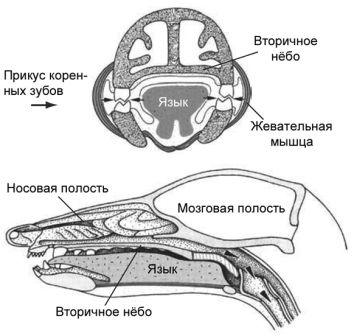
Рисунок 45. Некоторые из особенностей строения черепа млекопитающих, включая крупный язык. Многовершинные коренные зубы смыкаются специфическим образом, со скоординированным действием задействованных в этом жевательных мускулов. Вторичное нёбо разделяет ротовую полость и носовые проходы, облегчая одновременные жевание и дыхание. Из Maier 1999, с изменениями.
Яйца однопроходных чрезвычайно мелкие и не обладают твёрдой скорлупой, поэтому вероятность их сохранения в ископаемом состоянии чрезвычайно низкая. Из соображений парсимонии следует принять тот факт, что самые ранние, стволовые виды линии сумчатые+плацентарные также откладывали яйца, но у нас нет никаких ископаемых остатков, документирующих это. В ходе эволюционного развития линии, ведущей к последнему общему предку сумчатых и плацентарных, возникло живорождение. Некоторые авторы, напротив, утверждают, что различия между сумчатыми и плацентарными в репродуктивной анатомии и физиологии предполагают независимое возникновение живорождения в двух этих группах.[94] Ещё одна проблема, касающаяся различия между сумчатыми и плацентарными, легче трактуется с позиций палеонтологии, поскольку относится к особенностям строения твёрдых тканей, сохранившимся в виде ископаемых остатков. Ныне живущие сумчатые демонстрируют специализацию в развитии их зубов, когда в каждой челюсти сменяется лишь последний премоляр. Используя рентгеновскую компьютерную томографию сверхвысокого разрешения, Ричард Чифелли и его коллеги показали наличие порядка смены зубов, сходного с тем, что характерен для сумчатых, у мелового стволового вида Alphadon. Этот способ также отмечался у известных по более полным остаткам стволовых форм сумчатых из палеоцена, у животных, которые жили в местности, ныне являющейся частью Боливийского Альтиплано. Поскольку этот тип смены зубов ассоциируется с характерным для сумчатых способом воспроизводства, включающим малую продолжительность беременности и очень незрелое состояние на момент рождения, была выдвинута гипотеза о том, что эти особенности истории жизни характеризовали родственные базальные виды сумчатых из мела и палеоцена.[95]Летопись окаменелостей гоминид
Люди — уникальная разновидность млекопитающих, и многое в их происхождении было документировано в плане палеонтологии. Палеоантропологи признают ископаемые остатки, представляющие двадцать видов нашей родословной линии, подсемейства гоминин, и, хотя они показывают общую тенденцию превращения обезьяноподобных особенностей в человекоподобные с течением времени, наше эволюционное древо имеет много ветвей, примеров параллельного развития и эпизодов вымирания. Люди — это не вершина развития единственной и постоянной тенденции. Как же общие с шимпанзе и бонобо особенности предковой обезьяны, жившей примерно семь миллионов лет назад, преобразовались в особенности современных людей? Когда в ходе эволюции гоминин впервые возник человеческий тип онтогенеза? Нам мало известно то, как ископаемые виды гоминин росли и развивались от рождения до смерти — лишь немного ископаемых образцов незрелых особей сохранилось достаточно хорошо. Самой яркой находкой был обнаруженный в 1925 году Раймондом Дартом «ребёнок из Таунга», Australopithecus africanus из Южной Африки, возраст смерти которого оценивается в три или четыре года. У нас есть кое-какая информация в отношении Homo erectus на основе на нескольких образцов. Имеется неполный череп от особи с Явы, которая, по оценкам, умерла в возрасте примерно от одного до четырёх лет. Есть также скелеты мальчика Homo erectus из Нариокотоме (Кения), который достиг возраста от восьми до девяти лет, и подростка из Дманиси (Грузия), чей возраст составляет от одиннадцати до тринадцати лет. Исследования некоторых из этих материалов показали, что характером роста мозга Homo erectus был похож на шимпанзе и отличался от современных людей.
Рисунок 46. Развитие черепа у видов родословной линии людей, шимпанзе и бонобо. Онтогенетическая траектория каждого вида указана серой стрелкой. Современный вид и неандерталец представлены новорождёнными (детьми до прорезывания первых коренных зубов) и взрослыми особями; более ранние гоминиды — теми ископаемыми образцами, которые лучше всего соответствуют этим стадиям жизни. Числа обозначают объём эндокрана, приблизительно соответствующий объёму мозга, в кубических сантиметрах. Вертикальная ось представляет время роста в индивида. Обратите внимание на эволюционную тенденцию к укорачиванию онтогенетической траектории. По рисунку, любезно предоставленному Золликофером и Понс де Леон, после публикации их работы в 2010 году, с изменениями.
Самый ранний из известных гоминид — Sahelanthropus tchadensis. Он представлен лишь образцами взрослых особей, но, согласно моим коллегам из Цюриха Кристофу Золликоферу и Марсии Понс де Леон, сравнительный анализ хорошо сохранившегося черепа этой ископаемой формы с онтогенетической аллометрией крупной человекообразной обезьяны и человека указывает на то, что траектория роста черепа Sahelanthropus после рождения, вероятнее всего, напоминала таковую у шимпанзе. Сравнительное исследование людей и крупных человекообразных обезьян, а также тех обрывков и крупиц информации, которые дают нам ископаемые остатки, показывает, что видоспецифичные морфологические черты, те, которые позволяют нам отличать кости одного вида от другого, формируются до рождения. Это не слишком хорошо, поскольку ископаемых остатков неродившихся особей никогда не находили. С другой стороны, многое из того, что делает людей уникальными, касается изменений в особенностях истории жизни; критическим для них является послеродовой период, и, к счастью, ископаемые остатки, свидетельствующие об этом, у нас имеются.За рамками гипотезы фетализации
Что есть особого в индивидуальном развитии человека? Если сравнивать нас с нашими ближайшими родственниками, крупными человекообразными обезьянами, то мы, люди, характеризуемся рядом особенностей, среди которых долгий период беременности, беспомощные новорождённые с крупным мозгом, долгое детство, позднее прорезывание коренных зубов, растянутый срок роста тела, позднее наступление репродуктивного возраста, долгая пострепродуктивная фаза жизни и большая продолжительность жизни. Многие из анатомических особенностей людей, такие, как редкий волосяной покров на теле, напоминают ранние стадии в развитии современных обезьян. Форма черепа у взрослых современных людей, а также у неандертальцев, в целом сходна с таковой у детёнышей шимпанзе, бонобо, австралопитецин и реконструированного последнего общего предка людей и крупных человекообразных обезьян. Это привело к появлению гипотезы фетализации, которая утверждает, что анатомия взрослого человека подобна неполовозрелым формам предковых видов. Сформулированная в первой половине девятнадцатого века, эта гипотеза была вторично выдвинута в 1977 году Стивеном Джеем Гулдом в контексте его классической работы «Онтогенез и филогенез». Гулд интересовался прежде всего изменениями временных интервалов в ходе индивидуального развития, и люди стали превосходным случаем для изучения. По словам живущего в Базеле биолога Адольфа Портманна, а также других авторов, упомянутых выше, Гулд заявил, что человек — это, по существу, «плод вне матки» на протяжении первого года жизни. Первое время жизни после родов у людей состоит из событий, которые в случае человекообразных обезьян протекают в матке. Считалось, что на «рождение в недозрелом виде» у человека оказывает влияние ширина таза матери, ограничивающая увеличение окружности головы младенца. Утверждалось, что продолжительность роста нервной системы у людей должна выходить за рамки предкового состояния, характерного для крупной человекообразной обезьяны. Гипотеза фетализации доминировала над сравнительным онтогенетическим анализом эволюции человека. Но не все человеческие особенности могут быть описаны с помощью модели фетализации. Некоторые особенности онтогенеза и истории жизни крупных человекообразных обезьян отличаются от человеческих, но не всегда в сторону фетализации: у людей на протяжении первых нескольких лет жизни рост мозга происходит очень быстро в абсолютном исчислении, но медленно в плане сроков достижения некоторого процента от размера мозга взрослого индивида. Другой существенный момент состоит в том, что у человеческих матерей способность к воспроизводству восстанавливается задолго до того, как отнятый от груди ребёнок становится независимым, что позволяет людям производить многих зависимых от матери потомков. Такая стратегия имеет право на существование благодаря кооперативному размножению, характерному для нашего вида. Это очень сильно отличается от ситуации, имеющей место у крупных человекообразных обезьян. Маленькое лицо и большая черепная коробка, характерные для черепа взрослого человека, являются результатом замедленного развития черепа в сочетании с быстрым и длительным ростом мозга. Размеры мозгового отдела черепа у новорождённых современных людей и неандертальцев подобны таковым у взрослых шимпанзе, бонобо и Sahelanthropus, но после рождения он увеличивается примерно в четыре раза по сравнению с размером этого отдела черепа у взрослого шимпанзе. Картина роста человеческого черепа в целом должна была сложиться у последнего общего предка неандертальцев и людей, который жил по крайней мере полмиллиона лет назад. Но важные различия между этими двумя видами, возникшие после их расхождения, зафиксированы в летописи окаменелостей.Младенцы неандертальцев и рост
Многие данные, касающиеся постнатального онтогенеза неандертальцев, наших ближайших вымерших родственников, надёжно документированы. Ископаемые остатки большей частью фрагментарны, деформированы и разрозненны, но благодаря использованию компьютеризированных методов теперь возможно реконструировать значительную часть изменений скелета в период от рождения до взрослой жизни. Различия между черепами человека и неандертальца присутствуют уже у новорождённых, как это происходит у шимпанзе и бонобо. У современных людей маленькое лицо и шаровидная черепная коробка, а подбородок является единственной заметной выпуклостью на лице. С другой стороны, у неандертальцев были более высокие, сильнее выступающие лица и рельефный жевательный аппарат. После рождения у неандертальцев и людей отмечается сходный характер роста черепа, но темпы развития черепа у человека несколько ниже, и потому черепа взрослых людей больше сходны по форме с черепами подростков-неандертальцев. Работа, проведённая в Институте Макса Планка в Лейпциге, выявила различия в относительном росте некоторых областей черепа, которые у людей приводят к появлению выпуклостей на вершине и на боках головы (теменная и височная области), результатом чего является шаровидная форма нашего мозга.Распределение во времени событий из истории жизни
Важным маркером картины роста и событий истории жизни является время прорезывания зубов.[96] Время прорезывания первых постоянных верхних коренных зубов (M1) связано с прекращением роста мозга. У современных людей прорезывание M1 означает возрастной промежуток от 4,74 до 7,0 лет; у Homo erectus это время составляет примерно 4,5 года, что приближается к типичному значению для диких шимпанзе, у которых M1 прорезывается в возрасте примерно 4,0 лет. Как и другие млекопитающие, люди снабжают своё потомство пищей и обеспечивают ему иммунную защиту посредством молочного вскармливания. Это явление представляет собой главный аспект материнского вклада в воспроизводство, и оно также означает задержку рождения последующего потомства. Завершение выделения молока и кормления, и вместе с этим переход на взрослый рацион называется отнятием от груди. Это увеличение возможностей обработки пищи в значительной степени совпадает с появлением более постоянной зубной системы. Прорезывание первых коренных зубов и первые постоянные зубы, появляющиеся в ротовой полости у большинства приматов, коррелируют у многих видов с отнятием от груди. Эта корреляция слабее выражена у гоминоидов, что затрудняет создание простой реконструкции процесса отнятия от груди для ископаемых видов с сохранившимися данными о прорезывании зубов. На основе работы Адольфа Шульца (1891–1976) в Цюрихском Университете была предложена схема соотношения между порядком прорезывания коренных зубов и некоренных зубов второй смены (предкоренные, клыки, резцы), ростом и созреванием для приматов. Быстрый рост, подразумевающий раннее половое созревание и короткую продолжительность жизни, коррелирует с ранним прорезыванием коренных зубов и поздним прорезыванием предкоренных. С другой стороны, медленный рост связан с поздним прорезыванием коренных зубов и ранним прорезыванием второй смены зубов. «Правило Шульца» очень тщательно исследовалось, и было выяснено, что имеется множество исключений из него. Тем не менее, общая тенденция существует. Гоминоиды имеют более низкий уровень смертности, чем прочие обезьяны Старого Света. Последние обычно приступают к размножению в раннем возрасте, тогда как гоминоидов характеризует отсрочка начала размножения на более зрелый возраст. Это коренное различие отражено в распределении времени прорезывания зубов.
Рисунок 47. Согласно «правилу» Шульца, у быстрорастущих млекопитающих коренные зубы прорезываются раньше по отношению к смене молочных зубов постоянными, тогда как раннее прорезывание постоянных зубов характеризует медленно растущих млекопитающих. По Godfrey и др. 2005, с изменениями.
Гистологические особенности зубов также могут стать источником информации, касающейся времени наступления различных событий в истории жизни. Традиционно они изучались посредством разрезания драгоценных образцов. Сегодня для обнаружения микроскопических внутренних линий роста можно использовать изображения, полученные в ходе исследований с использованием синхротронной технологии (ускорителя частиц, использующего магнитные и электрические поля) вместо использования более традиционного и разрушительного подхода, связанного с изготовлением гистологических срезов. Благодаря изучению линий нарастания в зубах можно точно соотнести темпы развития и распределение событий жизни во времени. Использование этого подхода показало, что у австралопитецин и ранних представителей рода Homo был короткий период роста, чем они больше напоминали шимпанзе, нежели ныне живущих людей. Главный интерес состоял в том, чтобы установить момент, когда возникло современное состояние относительно долгого детства. К этому вопросу обратились в ходе исследования ископаемого ребёнка из Марокко, который жил около 160000 лет назад, и его сравнения с другими ископаемыми и современными популяциями людей. Ископаемые остатки показали степень развития зубов, эквивалентную ныне живущим человеческим детям того же самого возраста, около восьми лет, и совокупность особенностей истории жизни современного человека, с современным состоянием продолжительного развития зубов. Индивидуальное развитие познавательных процессов в жизни человека — это ещё один основополагающий аспект для нашего самопознания, но, к сожалению, ископаемые остатки дают слишком мало информации в этом плане, если вообще дают хоть что-то. Как сказали Золликофер и Понс де Леон, «археологическая летопись может быть здесь более информативной, поскольку она представляет собой форму сохранённого в ископаемом состоянии когнитивного поведения. Однако, к сожалению, она ничего не говорит об индивидуальном возрасте изготовителей орудий».Глава десятая О трилобитах, ракушках и букашках
До этого момента я рассматривал главным образом развитие позвоночных животных. В иных, нежели позвоночные, группах животных существует намного больше ныне живущих видов, и конечно, то же самое справедливо для вымерших видов. В этой главе я постараюсь показать некоторые из открытий в этой области и тот большой потенциал, которым обладают исследования этих животных. Чтобы палеонтология развития была применима к беспозвоночным, потребовалась бы полная переработка объекта исследования, и здесь этого, разумеется, не пытались делать. Скелет позвоночного, который как раз и сохраняется в ископаемом состоянии чаще всего, находится внутри тела и, таким образом, представляет собой эндоскелет. Для многих других животных характерен наружный экзоскелет, который покрывает собой мягкие части тела. Так обстоят дела у членистоногих животных, таких, как обладающие богатым видовым разнообразием насекомые, омары и крабы, а также вымершие трилобиты. Как я уже говорил ранее, касаясь палеогистологических исследований, эндоскелет позвоночных подвергается растянутой во времени модификации по мере роста животного. Модификация имеющихся в наличии частей эндоскелета позвоночных — это один из четырёх основных типов роста, которые мы можем наблюдать у животных. Добавление слоёв по мере роста — это другой вид, это нарастание, типичное для минерализованных раковин. Колебания условий окружающей среды и история жизни индивидуума отражены в скорости и характере этого нарастания. Новаторское исследование этого явления привело к открытию того, что в девоне в году было примерно 400 дней, а в пенсильвании{15} около 387, как позже подтвердили астрономы, которые выяснили, что скорость вращения Земли замедляется. Джон Уэллс изучал кораллы в 1960-е годы и заметил у них тонкие слои нарастания, отражающие ежедневные циклы, а также толстые слои, отражающие ежегодный прирост. Собрав эту информацию воедино, он сделал это удивительное открытие. Другой вид роста происходит не посредством добавления ещё большего количества слоёв, а добавлением вместо этого большего количества отдельных элементов: когда морские ежи наращивают новую пластинку на своём панцире или трилобиты добавляют новый сегмент тела. Случай с трилобитами особенно показателен в плане эволюции индивидуального развития, о чём я буду говорить далее. Трилобиты, подобно другим членистоногим, также претерпевали линьку. При таком способе развития имеющийся экзоскелет неоднократно сбрасывается и образуется новый. Поскольку, сформировавшись, экзоскелет не может растягиваться, данное животное должно проходить через стадии «мягкого панциря» между сменами экзоскелета меньшего размера на новый, больший. Абсорбция минеральных веществ из экзоскелета для их дальнейшего использования выглядит энергозатратным процессом, но фактически она имеет место у многих современных членистоногих, хотя явно отсутствовала у трилобитов. Линечные шкурки могут сохраняться в виде окаменелостей, которые способны ввести в заблуждение даже специалиста. Форма экзоскелета может сильно варьировать от одной стадии к другой, и по этой причине нужно множество образцов, чтобы понять, что серии ископаемых образцов, не связанных друг с другом плавным переходом, фактически представляют собой стадии развития одного и того же вида, разделённые промежутками, представляющими собой периоды роста.Трилобиты!
Лишь немногие группы животных, возможно, кроме динозавров и аммонитов, могут конкурировать с трилобитами за право быть самой узнаваемой группой ископаемых животных. Восклицательный знак, проставленный выше, таким образом, вполне оправдан, и он относится к книге Ричарда Форти с таким же названием{16}. Эта разнообразная группа существовала с кембрия, примерно 526 миллионов лет назад, до конца перми, около 252 миллионов лет назад, когда Земля столкнулась с крупнейшим случаем массового вымирания. Кальцитный скелет трилобитов хорошо сохраняется в виде окаменелости. Существует около 22000 описанных видов, и это большое количество видов и их отдельных представителей содержит в себе потенциал для исследования эволюции индивидуального развития в масштабах, колеблющихся от уровня популяции до истории группы в целом. План строения тела трилобитов включает деление на головной/туловищный отделы. Голова несёт на себе парные антенны плюс три пары других придатков. А туловище — это самая важная часть тела для исследований в области индивидуального развития, поскольку мы можем изучать эволюцию количества сегментов тела, их распределение по отделам тела и способы их роста. В эволюции туловищных сегментов трилобитов существует важная тенденция. В ходе ранней эволюции трилобитов, в кембрии, существовало значительное разнообразие количества этих сегментов во взрослом состоянии на внутривидовом, межвидовом и родовом уровнях. Изменчивость на этих уровнях снизилась на более поздней стадии эволюции трилобитов. Была выдвинута гипотеза о том, что это изменение представляло собой «застывание» систем, управляющих развитием, означавшее возросшее количество ограничений после установления «каналов» в развитии. После времени эволюционных «экспериментов» процесс развития более или менее установился, и в нём наверняка были задействованы Hox-гены. Известно, что у членистоногих, как и у позвоночных, Hox-гены играют важную роль в разделении тела на отдельные сегменты, и можно принять как факт то, что контроль сегментации и разбиения на отделы тела у трилобитов мог достигаться подобным же образом. Добавление сегментов в тело происходит на его конце, и, подобно тому, как множество молекул играет сходную роль в этом процессе у насекомых и ракообразных, так же, вероятно, происходило и у трилобитов. В главе 7 я указал, что некоторые группы позвоночных обладают очень пластичными генетическими механизмами сегментации, результатом чего становится изменчивое количество позвонков, тогда как другие группы очень консервативны. Аналогичная ситуация имеет место у трилобитов, и Найджел Хьюз, который провёл много исследований в области эволюции их индивидуального развития, документировал примеры значительной пластичности в развитии сегментации у геологически более молодых форм. Если ограничения и существуют, то они не являются незыблемыми. В ранний период эволюции трилобитов разбиение тела на отделы было неглубоким — все сегменты выглядели более или менее похожими друг на друга. У более поздних трилобитов изменение плана строения тела привело к более ярко выраженному разделению его на отделы. Различные разновидности трилобитов в процессе эволюции независимо приобрели отчётливо выраженную многосегментную хвостовую пластину, состоящую из многочисленных слившихся воедино туловищных сегментов и называемую пигидиумом. Это новшество имело экологическое значение: оно сделало возможным сворачивание тела в капсулообразное защитное положение, когда все мягкие части оказываются эффективно защищёнными. Защита от хищников, вероятнее всего, была главным фактором отбора, стимулирующим эволюцию механизма сворачивания тела. Сегменты в новом, увеличенном хвостовом отделе обычно чётко отличались от предшествующих туловищных сегментов. У трилобитов с данной особенностью строения количество сегментов стало менее изменчивым. У позвоночных существует аналогичная ситуация. Животные, у которых разделение позвоночника на отделы нечётко выражено или вовсе отсутствует, обладают более изменчивым количеством сегментов. Это наблюдается у змей и китов: и у тех, и у других отсутствует тазовый пояс. Различие между стабильным и изменчивым количеством сегментов может выглядеть тривиальным фактом. Но оно имеет самое прямое отношение к исследованиям индивидуального развития, поскольку изменчивость — это ключ к эволюционному потенциалу. Пластичность в развитии может быть фактором, который следует рассматривать при исследовании потенциального риска вымирания вида или группы организмов. Исследователи обнаружили, например, что трилобиты, демонстрировали снижение пластичности с течением геологического времени, и есть повод задуматься над тем, какое воздействие это оказало на характер их эволюции — это предмет исследований в настоящее время. Эволюция трилобитов с течением времени также исследовалась с использованием различных подходов. На протяжении десятилетий многие специалисты пробовали реконструировать части эволюционного древа трилобитов, исследуя особенности строения, сравнивая их у разных видов и далее проводя их числовой анализ. Для этого они используют кладистическую методологию, в который распределение особенностей строения оптимизируется с целью создания «лучшего» эволюционного древа, с применением принципа парсимонии. В основном лучшее из возможных объяснений для совокупности данных представляет собой та топология древа, отражающая главенствующую гипотезу отношений, для которой должно быть сделано наименьшее количество предположений (отсюда «парсимония»{17}). В процессе кодирования морфологических особенностей в целях анализа можно заметить, что у некоторых видов они стабильны. Но другие особенности могут изменяться в пределах вида — это случаи полиморфизма. В 2007 году Марк Уэбстер опубликовал исследование, в котором он количественно определил степень полиморфизма для 982 видов трилобитов с течением геологического времени и обнаружил, что более древние виды, которые находятся ближе к основанию эволюционного древа трилобитов, проявляли больший полиморфизм, чем поздние, более продвинутые виды. Почему же имеет место эта чёткая закономерность? Она может быть следствием изменений в окружающей среде, которые со временем приводили к различным типам давления отбора. Или она могла быть связанной с «внутренним» сужением системы, управляющей индивидуальным развитием. Но как бы то ни было, важно то, что Уэбстер открыл закономерность, сходную с той, которая в общих чертах наблюдается для количества сегментов в масштабах геологического времени. Во всех этих исследованиях индивидуального развития сквозь призму геологического времени очень важно безошибочное определение видов. В случае с ныне живущими видами, если виды распознаны на основе репродуктивной изоляции, экспериментальными манипуляциями можно проверить эту изоляцию. С другой стороны, виды, определяемые таким образом, невозможно выявить в ископаемом виде. Но таксономия трилобитов имеет под собою твёрдые основания. Исследованы сотни и тысячи экземпляров, и степень изменчивости среди них измерена. Непротиворечивые и воспроизводимые критерии, которые работают применительно к ныне живущим членистоногим, используются для определения морфологических границ видов. Другие группы беспозвоночных также известны по тысячам образцов, и исследования в области палеонтологии развития применительно к ним также являются возможными.
Рисунок 48. Два различных плана строения тела у трилобитов. Слева: в данном случае всё тело состоит из более или менее однородных сегментов до самого конца животного. Этот вид — Cambropallas telesto из среднего кембрия Марокко. Справа: самые задние сегменты достаточно сильно отличаются по строению и образуют хвостовую пластину; это пример разбиения тела на отделы. Данный вид — Zlichovaspis rugosa из раннего/среднего девона области Майдер, Марокко. Фотографии любезно предоставил Кристиан Клюг (Цюрих).
Биоминерализация
Прекрасная способность к биоминерализации у нескольких групп беспозвоночных делает их пригодными для палеонтологических исследований, поскольку образующиеся в итоге панцири обладают потенциалом для сохранения в ископаемом состоянии.[97] В процессе эволюции биоминерализация много раз независимо возникала у многих групп живых организмов, и палеонтологические знаки этого просто невозможно не увидеть.[98] Один из классических примеров — «меловые утёсы» южной Англии. Этот белый известняк мелового периода образует белые утёсы Дувра и обращённые к ним утёсы на противоположном берегу Ла-Манша, во Франции. При большом увеличении можно увидеть, что меловые утёсы состоят из миллионов кокколитов, каждый из которых размером лишь в несколько тысячных долей миллиметра в поперечнике. Их произвели одноклеточные морские водоросли, называемые кокколитофорами. Некоторые растения также могут подвергаться биоминерализации. Процесс, который приводит к образованию силикатов в морских водорослях, раковин моллюсков, а также других карбонатов у беспозвоночных, по своей сути аналогичен тому, который участвует в образовании карбонатного скелета у позвоночных. Фосфат и карбонат кальция — это биоминералы, встречающиеся чаще всего. Органические составляющие, такие, как коллаген и хитин, используются в комплексе с ними, обеспечивая структурную поддержку костей и панцирей. Получающиеся в итоге материалы имеют сложную архитектуру на микроскопическом, и даже на наноуровне. Результатом этого может исключительная стойкость к износу или сопротивление на излом. Учёные, исследующие материалы и структуры на наноуровне, очень заинтересовались этими органическими структурами и используют изощрённые методы получения изображений, чтобы обнаружить структуры, которые развились в ходе эволюции у живых организмов, как у ископаемых, так и у современных. Одной из целей этого является получение знаний, которые можно применить при искусственном синтезе органо-неорганических материалов. На более высоком макроуровне эволюция способности к биоминерализации также привлекает всё больше и больше внимания по причине понимания того, что это фундаментальная часть исследований эволюции истории жизни крупных групп организмов, которые населяют океаны сейчас и населяли их в прошлом (см. главу 6). Кораллы, организмы, активно участвующие в процессах биоминерализации, представляют особый интерес в настоящее время. Повышение содержания двуокиси углерода в атмосфере увеличивает кислотность морской воды, что приводит к декальцификации карбонатных скелетов. Смогут ли получившиеся в итоге «голые» кораллы выжить и возобновить процесс кальцификации, если содержание двуокиси углерода в атмосфере уменьшится? Что происходило с кораллами в геологическом прошлом, когда они подвергались давлению отбора, действующему в ином направлении? Примером такого рода являются склерактиниевые кораллы. Их предки были «голыми» кораллами, похожими на актиний, которые пережили массовое вымирание пермского периода. Первые склерактинии появились в среднем триасе; это были первые кораллы после события массового вымирания в конце пермского периода. До недавнего открытия, опубликованного Ярославом Столярски и его коллегами, считалось, что склерактиниевые кораллы образовывали исключительно арагонитовые скелеты. Но превосходно сохранившаяся окаменелость из верхнего мела обладала полностью кальцитным скелетом. Это даёт основание полагать, что эти кораллы могли строить скелеты из различных полиморфных модификаций карбонатов, как делали некоторые, хотя и не все группы морских организмов, отлагающих карбонат кальция. Исследование геохимических свойств ископаемых организмов открыло их способности изменять систему биоминерализации, о чём даже не подозревали ранее. Что определяет эту гибкость? Как и в случае с трилобитами, которые обсуждались выше, не все группы были одинаково гибкими, или «пластичными» в отношении процесса индивидуального развития. Какое воздействие оказывало это обстоятельство на картину эволюции в прошлом, и какое воздействие может оказать на будущее нынешних морских экосистем, можно узнать в ходе изучения организмов, обладающих раковинами и панцирями.О раковинах моллюсков
Благодаря биоминерализации раковины моллюсков хорошо сохраняются в виде окаменелостей. Тысячи их экземпляров собраны в палеонтологических коллекциях, отражая видовое разнообразие и изобилие экземпляров и тем самым позволяя проводить всесторонние исследования картины эволюции сквозь призму геологического времени. Чтобы извлечь пользу из этого богатого источника информации, нам следует охарактеризовать это разнообразие морфологически и таксономически, дав названия видам и разместив их на правильно построенных филогенетических древах, по которым и будут исследоваться закономерности эволюции. Проблема состоит в том, что каждая раковина обладает лишь немногими дискретнымиморфологическими признаками, с помощью которых её можно охарактеризовать, а явления конвергенции в форме и орнаментации представлены в летописи окаменелостей в большом количестве. Сравнение отличающихся друг от друга естественных групп моллюсков, таких, как аммониты и брюхоногие, например, показало, что в основе морфогенеза и роста раковины лежат общие принципы генерации. Поскольку в плане индивидуального развития раковина моллюска является высокоинтегрированным объектом, многие особенности раковины часто изменяются в пределах вида совместно. Этот высокий уровень интеграции делает трудным, и даже невозможным определение свойств, которые полностью независимы от других, но это также означает, что, работая с верно выбранной моделью, мы можем многое предсказать в эволюции формы раковины. Это интенсивно исследовалось на примере наиболее известных ископаемых моллюсков, аммонитов. Они образуют полностью вымершую группу головоногих моллюсков, класса, к которому принадлежат осьминоги и кальмары. Их более отдалённым родственником является Nautilus, также представитель головоногих моллюсков, который фактически является единственным ныне живущим видом этой группы с наружной раковиной. Онтогенез раковины аммонитов был объектом интенсивных исследований, и было выявлено три его основных аспекта: секреция раковины по мере того, как мягкое тело увеличивается в размерах, секреция дополнительных слоёв на внутренней поверхности камер тела и секреция септ в задней части тела, посредством чего образуются камеры в раковине. Первым шагом в онтогенезе аммонитов была эмбриональная стадия, которая обладала раковиной, называемой «аммонителла». Ископаемые остатки её — это маленькие узелки, которые обнаруживаются в хорошо сохранившихся окаменелостях в начале спирали, или же среди кучи неопределённых окаменелостей в углу музейного кабинета. По мере роста раковины возрастает сложность её орнаментации — разумеется, при наличии таковой. Орнаментация принадлежит к числу особенностей строения, как и концентрические и продольные рёбра, шипы, бугорки и кили. Обширные отметины на некоторых раковинах аммонитов позволяют предположить, что могла происходить частичная резорбция раковины, поскольку они отмечают собой разрывы в линиях роста. Орнаментация у аммонитов используется в целях таксономических исследований, и, как было известно на протяжении долгого времени, её степень коррелирует с другими особенностями раковины. Наиболее декорированные виды обладают наименее плотно свёрнутой раковиной, с почти круговыми завитками. Эта корреляция стала известна как закон Бакмана, в честь автора, который первым опубликовал сообщение об этой идее (1892). Ойвинд Хаммер и Хьюго Бюше пришли к заключению, что картина корреляции чётко проявляется внутри вида, но менее чётко на межвидовом уровне. Используя компьютерное моделирование, они продемонстрировали геометрические свойства системы, которые в итоге сводятся к связи развития рёбер и формы устья раковины.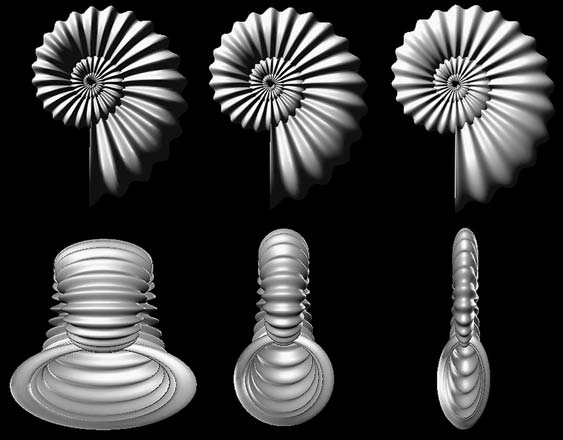
Рисунок 49. Компьютерные модели Хаммера и Бюше (2005) для гипотетических раковин аммонитов, иллюстрирующие корреляцию особенностей для пропорций, меняющихся на основе закона Бакмана. Вид сбоку (сверху) и со стороны устья (снизу). A: Расширенная в стороны (депрессная) раковина, у которой боковые рёбра становятся пропорционально мощнее. B: Раковина, сжатая с боков посредством уменьшения длины поперечной оси, с соответствующим образом слабее выраженными боковыми рёбрами. C: Аммонит с круглой в сечении полостью раковины и равной амплитудой боковой и вентральной ребристости. Поскольку амплитуда вентральных рёбер остаётся постоянной при изменении масштаба поперечной оси, возникает изменчивость в соотношения между амплитудами вентральных и боковых рёбер. По Hammer и Bucher, 2005, с изменениями.
Достижение понимания геометрии раковины, не только у аммонитов, но и у моллюсков вообще, заняло много времени у многих палеонтологов. В 1960-е годы, используя в свое работе осциллограф, Дэвид Рауп стал пионером в области компьютерной параметризации витков раковин, работы, которая заложила основу для возникновения области теоретической морфологии. Его работа, а также другие ранние методы оперировали главным образом двумя измерениями и предполагали наличие оси вращения, которая не имеет реального биологического значения, поскольку возникает a posteriori как результат приращения раковины. Кроме того, многие раковины нельзя рассматривать как закручивающиеся вокруг единственной оси вращения. Традиционные модели не принимали во внимание онтогенетические изменения раковины или периодизацию процессов роста. Авторы более новых подходов стараются учитывать эти проблемы. Одна из последних моделей, разработанная Северином Урди и его коллегами, имитирует форму устья и явление нелинейной аллометрии в процессе роста. Как изменяются эти виды аллометрии, можно проследить, изменяя параметры модели. Конечной целью этого является понимание правил, которым могут подчиняться некоторые текущие эволюционные процессы и межвидовая изменчивость. Биологический аспект этих исследований процесса роста включает подробное изучение мантии, мягкой и эластичной ткани, которая выделяет материал раковины. Мантия не сохраняется в ископаемом состоянии, но любые исследования её свойств и способностей, а также фенотипической пластичности на протяжении времени жизни отдельной особи должны представлять интерес для палеонтологов. Ископаемые остатки предоставляют нам огромный массив данных по морфологии, которые могут быть определены количественно с помощью ещё более сложных и приближенных к реальности морфометрических методы. Пример такого рода — метод, используемый Клодом Монне (не имеющим отношения к художнику) и его коллегами. На основе трёхмерных изображений, полученных путём сканирования с использованием компьютерной томографии на микроскопическом уровне, выделяются морфометрические параметры, которые используются для описания геометрии раковин моллюсков на протяжении всего их онтогенеза и для их сравнения в пределах вида. «Скелетизация» трёхмерных виртуальных раковин позволяет выделить центральную линию, используемую как опорный элемент при получении поперечных сечений раковины и при анализе формы полости с использованием геометрических морфометрических параметров. На основе полученных в ходе этого исследования параметров становится возможным сравнение онтогенетических траекторий различных особей. Аналогичные методы использовали Крис Кружински и его коллеги для количественной оценки морфологической изменчивости склерактиниевых кораллов.
Рисунок 50. Схематические изображения четырёх стадий онтогенеза позднемелового аммонита Hoploscpahites nicoletti, вид сбоку. A) аммонителла; B) неаноконх, первое постэмбриональное время; C) молодая особь; D) взрослая особь. Масштабная линейка = 5 мм. Обратите внимание на увеличение степени орнаментации. Раковина ювенильной особи сильнее сжата с боков и больше скручена, чем неаноконх. По Bucher et al. 1996, с изменениями.
Но, какие бы модели не разрабатывались для понимания эволюции формы раковины, они должны принимать во внимание окружающую среду, поскольку, как было показано в экспериментах и на примере естественных популяций, она оказывает влияние на скорость роста. Внешние переменные параметры среды включают доступность пищи, тип субстрата, солёность, количество растворённого кислорода, мутность, температуру, плотность популяции и экологические взаимоотношения. Например, ныне живущие виды брюхоногих моллюсков могут замедлить скорость своего роста в присутствии хищников, направляя дополнительный материал для построения раковины на локальное утолщение стенок раковины. Увеличение относительной толщины раковины, особенность, часто рассматриваемая как защитный механизм, также вызывается голоданием. Это ещё один пример фенотипической пластичности и диапазона фенотипов, связанных с генотипом, о чём я говорил в главе 2. Одним из основных параметров истории жизни, на который оказывает очень значительное воздействие окружающая среда, является продолжительность жизни.Моллюски с долгой и короткой жизнью
Продолжительность жизни — это основополагающая переменная величина в истории жизни. Подсчитывая годичные слои, образовавшиеся путём нарастания, можно оценить продолжительность жизни мёртвых (в том числе ископаемых) индивидуумов. Изучение ныне живущих видов показало, что на долговечность в целом оказывает влияние температура, при которой живут животные. Эту иную переменную величину также можно оценить для ископаемых образцов. Организмы, которые населяют более высокие широты, имеют тенденцию жить дольше, что связано с температурой. При температурах тела на уровне холодных южных областей океанов химические реакции протекают на малой скорости. Таким образом, процессы обмена веществ также протекают медленно, и итогом этого является жизнь без стрессов. Более или менее постоянные температура воды и солёность не оказывают значительного воздействия на регуляторные механизмы клеток. Любой обменный процесс производит побочные продукты, химические соединения, называемые радикалами, в каждой клетке, которая получает свою энергию при помощи кислорода. Более медленный обмен веществ на холоде производит меньше разрушительных побочных продуктов, которые необходимо переработать или уничтожить. Чем меньше организм испытывает стрессовых факторов, тем дольше он может прожить. Некоторым морским беспозвоночным холодная среда обитания может помочь избежать действия опасных радикалов. Многие виды в процессе эволюции приобрели другие способы продлевать свою жизнь. Например, циприна (арктика) исландская (Arctica islandica), представитель двустворчатых моллюсков, закапывается в бедные кислородом слои океанского дна на несколько дней. Там она затормаживает свой обмен веществ и частота её сердечных сокращений составляет лишь десятую часть от нормальной скорости. Когда моллюск выходит из этого состояния, митохондрии начинают функционировать в обычном режиме, вновь производя радикалы. Но в это же время некоторые гены активизируются и производят ферменты, которые борются с опасными новыми химическими соединениями, возникающими при высокой скорости метаболизма. Благодаря этой стратегии данный двустворчатый моллюск может достигать возраста более двухсот лет. У позвоночных такого свойства нет. Вместо того, чтобы замедлить обмен веществ, в условиях меньшей доступности кислорода митохондрии производят ещё больше радикалов. Различные картины роста и их отношение к температуре изучены для многочисленных ископаемых организмов, например, для моллюска Cucullaea raea из эоценовых пород острова Сеймур на Антарктическом полуострове. В эоцене температуры были гораздо выше, чем температура ниже нуля, которая характерна для нашего времени. Раковины этого двустворчатого моллюска росли очень медленно и только в полной темноте южнополярных зим. На удивление, они полностью прекращали расти во время южного полярного лета, когда пища была более обильной, вероятно, из-за того, что они вкладывали всю свою энергию и ресурсы в воспроизводство. Из-за своих медленных обмена веществ и скорости роста эти моллюски жили чрезвычайно долго. Некоторые демонстрируют линии роста, представляющие более ста лет жизни. Подобно Arctica islandica, они жили дольше, чем многие из людей.Необычная сохранность
Ископаемые отпечатки — это почти единственные свидетельства существования мягкотелых представителей биоты или мягких частей организмов. К счастью, только «почти». Сохранение беспозвоночных в некоторых редких местонахождениях окаменелостей заходит гораздо дальше обычного сохранения экзоскелета или раковины: мельчайшие детали целого тела, включая глаза и придатки, сохраняются в ископаемом состоянии на различных стадиях развития. Их обнаружение и изучение сложны, поскольку обычно они хрупкие и мелкие. Но благодаря новым методам получения изображений и компьютерным программам трёхмерной реконструкции открывается много новой информации. Местонахождения имеются по всему миру и среди них формация Крато из меловых отложений Бразилии, Золенхофен из юры Баварии, райниевые черты из девона Шотландии, и «орстенские» местонахождения по всему миру, лучше всего известные из Швеции и охватывающие период от нижнего кембрия до нижнего ордовика. Открытия и описания онтогенетических стадий столь большого количества таксонов быстро наполняют литературу. В дальнейшем накопление этой информации, синтез и анализ этих открытий, несомненно, окажут огромное влияние на наше понимание самых ранних стадий развития нескольких групп членистоногих. Некоторые международные исследовательские группы, такие, как группы Марка Саттона и Дерека Бриггса, используют сложное программное обеспечение для получения трёхмерных реконструкций в целях изучения крохотных и хрупких окаменелостей, заключающих в себе богатую морфологическую информацию. Пример, который я упоминаю здесь, касается изучения раннего представителя ракообразных из среднего кембрия Швеции, Henningsmoenicaris scutula. Йоахим Хауг и его коллеги идентифицировали десять онтогенетических стадий этого животного и создали, как они любят это называть, «четырёхмерную модель», где четвёртым «измерением» является время. Эти стадии документируют время и способ добавления сегментов и их придатков. Первая стадия имеет четыре сегмента, несущих придатки; это количество увеличивается до семи, после чего происходит дифференциация отделов тела.
Рисунок 51. 4-D модель Henningsmoenicaris scutula, примитивного ракообразного из кембрия Швеции. Рисунок любезно предоставил Йоахим Хауг (Нью-Хейвен).
Хауг и его коллеги документировали развитие сложных глаз, впервые распознаваемых на пятой стадии из десяти имеющихся в наличии. Каждая из последующих стадий документирует изменение от сидячих глаз к стебельчатым. Прекрасная морфология даёт нам не только эстетически приятное описание изменений формы, происходивших около 520 миллионов лет назад, но также данные, важные для понимания ранних этапов разделения ракообразных и других групп членистоногих в процессе эволюции. Какие способы развития характеризовали происхождение крупных групп членистоногих, в том числе трилобитов? Являются ли общие системы индивидуального развития признаком эволюционного родства, или они эволюционировали независимо в различных группах? Хауг и его коллеги заметили, что способ слияния последних туловищных сегментов у Henningsmoenicaris в ходе онтогенеза также присутствует у трилобитов: новые сегменты появляются и присоединяются к прилегающему к ним спереди сегменту, в противоположность картине, имеющей место у более поздних ракообразных. Эти и другие аспекты относительного времени возникновения онтогенетических событий можно нанести на эволюционные древа, чтобы отследить, насколько консервативным или пластичным было индивидуальное развитие на самых ранних стадиях эволюции многоклеточных животных.Эпилог Есть ли мораль в палеонтологии развития?
История жизни — это история изменений, и многое в ней записано окаменелостями в глубинах времени, где возникало и исчезало огромное разнообразие живых организмов. Можно ли извлечь из этого некое моральное послание? Конечно, нет. Не существует морали, присущей научному факту или гипотезе. Не так давно в нашем понимании эволюции, в том числе и эволюции человека, доминировала идея эгоистичного гена. Чуть позже биологические антропологи продемонстрировали нам, что мы, люди, являемся общественными человекообразными обезьянами, и что это биологически внедрено в нас. Это новое знание биологии не изменяет морального императива, который мы свободно выбираем, чтобы руководствоваться им. Поскольку всё изменяется и поскольку мы рано или поздно будем обречены как индивиды и как вид в целом, в чём смысл бытия? Или, забывая о необъятности времени, — поскольку мы, люди, есть ничто иное, как весьма особенные крупные человекообразные обезьяны, ничто иное, помимо биологии — почему мы боремся против нашей «природы»? Альберт Камю видел комфорт в стимуле продолжать движение, несмотря на свою экзистенциалистскую концепцию жизни. Сизиф был навечно осуждён вкатывать на гору валун, который раз за разом скатывался назад, к месту, откуда началось движение. Давайте получать удовольствие, толкая его вверх. Биология не преподаёт никаких экзистенциальных уроков, но может стать поводом для рефлексии. Безграничность времени и осознание того, что безграничность биологического многообразия делает нас всего лишь одним видом на огромном древе жизни, вселяет в нас скромность. Бесконечность красок жизни и изменения — то самое величайшее шоу на Земле — вызывает благоговение. Окружающая среда и индивидуальное развитие тесно переплетаются. Фенотип — это не просто результат проявления генотипа. Окружающая среда может вызвать изменения в развитии; даже ископаемые остатки организмов, будучи такими неполными, демонстрируют это, если знать, как читать их. Мы, люди, представляем собой лишь одну ветвь огромного древа жизни — но зато способную изменять окружающую среду с такой скоростью, которой до сих пор были способны достичь лишь крупнейшие геологические события. То, как мы продолжим воздействовать на окружающую среду, определит судьбу онтогенеза всех живых существ вокруг нас: скромно, считаясь с тем, чего каждый из нас может достичь; возможно, действуя локально, чтобы внести небольшой вклад, и находясь под управлением морального императива; действуя независимо, или даже вразрез с биологией, химией и физикой.Библиография
Abdala, F., and N. P. Giannini. 2000. Gomphodont cynodonts of the Chañares Formation: The analysis of an ontogenetic sequence. Journal of Vertebrate Paleontology 20: 501-6. Adams, D., and M. Carwardine. 1990. Last Chance to See. London: Pan Books. Ahlberg, P., K. Trinajstic, Z. Johanson, and J. A. Long. 2009. Pelvic claspers confirm chondrichthyan-like internal fertilization in arthrodires. Nature 460: 888-89. Alberch, P. 1989. The logic of monsters: Evidence for internal constraint in development and evolution. Geobios, mémoire spéciale, 12: 21–57. Amprino, R. 1947. La structure du tissu osseux envisagée comme expression de différences dans la vitesse de l’accroissement. Archives of Biology 58: 315-30. Anders, U., W. v. Koenigswald, I. Ruf, and B. H. Smith. 2011. Generalized individual dental age stages for fossil and extant placental mammals. Paläontologische Zeitschrift 85: 321-39. Anderson, J. S., R. R. Reisz, D. Scott, N. B. Fröbisch, and S. S. Sumida. 2008. A stem batrachian from the early Permian of Texas and the origin of frogs and salamanders. Nature 453: 515-18. Andrews, C. W. 1913. A descriptive catalogue of the marine reptiles of the OxfordClay, based on the Leeds Collection in the British Museum (Natural History), London. London: British Museum. Asher, R. J., N. Bennett, and T. Lehmann. 2009. The new framework for understanding placental mammal evolution. Bioessays 31: 853-64. Asher, R. J., and T. Lehmann. 2008. Dental eruption in afrotherian mammals. BMC Biology 6: 14. Azorit, C., J. Muñoz-Cobo, J. Hervás, and M. Analla. 2004. Aging through growth marks in teeth of Spanish red deer. Wildlife Society Bulletin 32: 702-10. Balanoff, A., and T. Rowe. 2007. Osteological description of an embryonic skeleton of the extinct elephant bird, Aepyornis (Palaeognathae: Ratitae). Society of Vertebrate Paleontology Memoir 9: 1-53. Barnosky, A. D. 2001. Distinguishing the effects of the Red Queen and Court Jester on Miocene mammal evolution in the northern Rocky Mountains. Journal of Vertebrate Paleontology 21: 172-85. Bell, M. A. 2009. Implications of a fossil stickleback assemblage for Darwinian gradualism. Journal of Fish Biology 75: 1977-99. Benton, M. J. 2000. Stems, nodes, crown clades, and rank-free lists: Is Linnaeus dead? Biological Review 75: 633-48. Benton, M. J. 2004. Vertebrate Palaeontology. 3rd ed. Malden, MA: Blackwell. Benton, M. J. 2009a. Paleontology and the history of life. In M. Ruse and J. Travis, eds., Evolution: The First Four Billion Years (pp. 80-104). Cambridge, MA: Belknap Press. Benton, M. J. 2009b. The Red Queen and the Court Jester: Species diversity and the role of biotic and abiotic factors through time. Science 323: 728-32. Benton, M. J. 2010. Studying function and behavior in the fossil record. PLoS Biology 8: e1000321. Billet, G. A., C. de Muizon, and B. M. Quispe. 2008. Late Oligocene mesotheriids (Mammalia, Notoungulata) from Salla and Lacayani (Bolivia): Implications for basal mesotheriid phylogeny and distribution. Zoological Journal of the Linnean Society 152: 153–200. Bininda-Emonds, O. R. P., J. E. Jeffery, and M. K. Richardson. 2003. Inverting the hourglass: Quantitative evidence against the phylotypic stage in vertebrate development. Proceedings of the Royal Society of London B: Biological Sciences 270 (1513): 341-46.Bininda-Emonds, O. R. P., J. E. Jeffery, M. R. Sánchez-Villagra, J. Hanken, M. Colbert, C. Pieau, L. Selwood, C. T. Cate, A. Raynaud, C. K. Osabutey, and M. K. Richardson. 2007. Forelimb — hind limb developmental timing across tetrapods. BMC Evolutionary Biology 7: 182. Black, K. H., M. Archer, S. J. Hand, and H. Godthelp. 2010. First comprehensive analysis of cranial ontogeny in a fossil marsupial from a 15-million-year-old cave deposit in northern Australia. Journal of Vertebrate Paleontology 30: 993-1011. Blackburn, D. G., and H. E. Evans. 1986. Why are there no viviparous birds? American Naturalist 128: 165-90. Blumberg, M. S. 2008. Freaks of Nature. Oxford: Oxford University Press. Bolk, L. 1926. Das Problem der Menschenwerdung. Jena: Fischer Verlag. Borel, E. 1913. Mécanique statistique et irréversibilité. Journal de Physique, 5e série, 3: 189-96. Botha-Brink, J., and K. D. Angielczyk. 2010. Do extraordinarily high growth rates in Permo-Triassic dicynodonts (Therapsida, Anomodontia) explain their success before and after the end-Permian extinction? Zoological Journal of the Linnean Society 160: 341-65. Bowler, P. J. 1983. The Eclipse of Darwinism: Anti-Darwinian Evolutionary Theories in the Decades around 1900. Baltimore: Johns Hopkins University Press. Brasier, M. D., O. R. Green, A. P. Jephcoat, et al. 2002. Questioning the evidence for earth’s oldest fossils. Nature 416: 76–81. Brayard, A., G. Escarguel, H. Bucher, C. Monnet, T. Brühwiler, N. Goudeman, T. Galfetti, and J. Guex. 2009. Good genes and good luck: Ammonoid diversity and the end-Permian mass extinction. Science 325: 1118-21. Brinkman, D. 1988. Size-independent criteria for estimating relative age in Ophiacodon and Dimetrodon (Reptilia, Pelycosauria) from the Admiral and lower Belle Plains Formations of west-central Texas. Journal of Vertebrate Paleontology 8: 172-80. Brinkmann, W. 1996. Ein Mixosaurier (Reptilia, Ichthyosauria) mit Embryonen aus der Grenzbitumenzone (Mitteltrias) des Monte San Giorgio (Schweiz, Kanton Tessin). Eclogae geologicae Helvetiae 89: 1321-44. Bryant, H. N. 1995. The threefold parallelism of Agassiz and Haeckel, and polarity determination in phylogenetic systematics. Biology and Philosophy 10: 197–217. Bryant, H. N., and A. P. Russell. 1992. The role of phylogenetic analysis in the inference of unpreserved attributes of extinct taxa. Philosophical Transactions of the Royal Society of London B 337: 405-18. Bucher, H., N. H. Landman, J. Guex, and S. M. Klofak. 1996. Mode and rate of growth in ammonoids. In N. H. Landman, K. Tanabe, and R. A. Davis, eds., Ammonoid Paleobiology. Topics in Geobiology 13: 407-61. Buckman, S. S. 1887–1907. A Monograph of the Ammonites of the Inferior Oolite Series. London: Palaeontographical Society. Buffetaut, E., J. Li, H. Tong, and H. Zhang. 2007. A two-headed reptile from the Cretaceous of China. Biology Letters 3: 80–81. Bürgin, T. 1999. Middle Triassic marine fish faunas from Switzerland. In G. H. Arratia and H. P. Schultze, eds., Mesozoic Fishes 2 — Systematics and Fossil Record (pp. 481-94). Munich: Pfeil Verlag. Butler, P. M. 1995. Ontogenetic aspects of dental evolution. International Journal of Developmental Biology 39: 25–34. Camus, A. 1942. Le mythe de Sisyphe. Paris: Gallimard. Canoville, A., and M. Laurin. 2010. Evolution of humeral microanatomy and lifestyle in amniotes, and some comments on paleobiological inferences. Biological Journal of the Linnean Society 100: 384–406. Carlini, A. A., M. R. Ciancio, and G. J. Scillato-Yané. 2010. Middle Eocene — early Miocene Dasypodidae (Xenarthra) of southern South America: Faunal succession at Gran Barranca — biostratigraphy and paleoecology. In R. H. Madden, A. A. Carlini, M. G. Vucetich, and R. F. Kay, eds., The Paleontology of Gran Barranca: Evolution and Environmental Change through the Middle Cenozoic of Patagonia (pp. 106-29). Cambridge: Cambridge University Press. Carroll, L. 1871. Through the Looking-Glass. London: Macmillan. Carroll, S. B. 2008a. Evo-devo and an expanding evolutionary synthesis: A genetic theory of morphological evolution. Cell 134: 25–36. Carroll, S. B. 2008b. Into the Jungle: Great Adventures in the Search for Evolution. San Francisco: Benjamin Cummings. Carpenter, K. 1999. Eggs, Nests, and Baby Dinosaurs: A Look at Dinosaur Reproduction. Bloomington: Indiana University Press. Cartelle, C., and G. De Iuliis. 2006. Eremotherium laurillardi (Lund)Xenarthra, Megatheriidae), the Panamerican giant ground sloth: Taxonomic aspects of the ontogeny of skull and dentition. Journal of Systematic Palaeontology 4: 199–209. Carter, A. M. 2008. What fossils can tell us about the evolution of viviparity and placentation. Placenta 29: 930-31. Castanet, J. 1994. Age estimation and longevity in reptiles. Gerontology 40: 174-92. Chiappe, L. M. 2007. Glorified Dinosaurs: The Origin and Early Evolution of Birds. New York: Wiley-Liss. Chiappe, L. M., R. A. Coria, L. Dingus, F. Jackson, A. Chinsamy, and M. Fox. 1998. Sauropod dinosaur embryos from the Late Cretaceous of Patagonia. Nature 396: 258-61. Chiappe, L. M., J. Marugán-Lobón, S. Ji, and Z. Zhou. 2008. Life history of a basal bird: Morphometrics of the early Cretaceous Confuciusornis. Biology Letters 4: 719-23. Chinsamy-Turan, A. 2001. Dinosaur growth — egg to adult. In Encyclopedia of Life Sciences. New York: Wiley. Chinsamy-Turan, A. 2005. The Microstructure of Dinosaur Bone: Deciphering Biology with Fine-Scale Techniques. Baltimore: Johns Hopkins University Press. Chinsamy-Turan, A., and J. H. Hurum. 2006. Bone microstructure and growth patterns of early mammals. Acta Paleontologica Polonica 51: 325-38. Chritz, K. L., G. J. Dyke, A. Zazzo, A. M. Lister, N. T. Monaghan, and J. D. Sigwart. 2009. Palaeobiology of an extinct Ice Age mammal: Stable isotope and cementum analysis of giant deer teeth. Palaeogeography, Palaeoclimatology, Palaeoecology 282: 133-44. Cifelli, R. L., and C. de Muizon. 1998. Tooth eruption and replacement pattern in early marsupials. Compte Rendus de l’Academie des Sciences de Paris, Sciences de la Terre et des Planètes 326: 215-20. Clark, J. M., M. A. Norell, and L. M. Chiappe. 1999. An oviraptorid skeleton from the late Cretaceous of Ukhaa Tolgod, Mongolia, preserved in an avian-like brooding position over an oviraptorid nest. American Museum Novitates 3265: 1-36. Cloutier, R. 2010. The fossil record of fish ontogenies: Insights into developmental patterns and processes. Seminars in Cell and Developmental Biology 21: 400–413.Coates, M. I., and M. Ruta. 2007. Skeletal changes in the transition from fins to limbs. In B. K. Hall, ed., Fins to Limbs (pp. 15–38). Chicago: University of Chicago Press. Cole, K. S., ed. 2010. Reproduction and Sexuality in Marine Fishes: Patterns and Processes. Berkeley: University of California Press. Colosimo, P. F., K. E. Hosemann, S. Balabhadra, G. Villarreal Jr., J. Grimwood, J. Schmutz, R. M. Myers, D. Schluter, and D. M. Kingsley. 2005. Widespread parallel evolution in sticklebacks by repeated fixation of ectodysplasin alleles. Science 307: 1928-33. Crompton, A. W., and W. L. Hylander. 1986. Changes in mandibular function following the acquisition of a dentary-squamosal jaw articulation. In N. Hotton III, P. Maclean, J. J. Roth, and E. C. Roth, eds., The Ecology and Biology of Mammal-Like Reptiles (pp. 263-82). Washington, DC: Smithsonian Institution. Crompton, A. W., and Z.-X. Luo. 1993. Relationships of the Liassic mammals Sinoconodon, Morganucodon oehleri, and Dinnetherium. In F. S. Szalay, M. J. Novacek, and M. C. McKenna, eds., Mammal Phylogeny (pp. 30–44). New York: Springer. Curry, K. A. 1999. Ontogenetic histology of Apatosaurus (Dinosauria: Sauropoda): New insights on growth rates and longevity. Journal of Vertebrate Paleontology 19: 654-65. Cuvier, G. 1835-39. Leçons d’anatomie comparée. Vols. 1–6. Reprint, Ann Arbor: University of Michigan Press, 1980. Darwin, C. R. 1868. The Variation of Animals and Plants under Domestication. London: John Murray.{18} Dawkins, R. 2009. The Greatest Show on Earth: The Evidence for Evolution. New York: Simon and Schuster.{19} De Beer, G. 1962. Embryos and Ancestors. 3rd ed. London: Clarendon Press. Delfino, M., and M. R. Sánchez-Villagra. 2010. A survey of the rock record of reptilian ontogeny. Seminars in Cell and Developmental Biology 21: 432-40. Dewar, E. 2007. The taxonomic stability of large mammals in the White River Chronofauna masked their changing dietary ecology. Journal of Vertebrate Paleontology 27: 68A. Dobzhansky, T. 1973. Nothing in biology makes sense except in the light of evolution. American Biology Teacher 35: 125-29. Domazet-Lošo, T., and D. Tautz. 2010. A phylogenetically based transcriptome age index mirrors ontogenetic divergence patterns. Nature 468: 815-18. Donoghue, P. C. J. 2005. Saving the stem group — a contradiction in terms. Paleobiology 31: 553–58. Donoghue, P. C. J., A. Graham, and R. N. Kelsh. 2008. The origin and evolution of the neural crest. Bioessays 30: 530-41. Donoghue, P. C. J., and M. A. Purnell. 2005. Genome duplication, extinction, and vertebrate evolution. Trends in Ecology and Evolution 20: 312-19. Duboule, D. 1994. Temporal colinearity and the phylotypic progression: A basis for the stability of a vertebrate Bauplan and the evolution of morphologies through heterochrony. Development (Suppl.) 135-42. Duboule, D. 2010. The evo-devo comet. EMBO Reports 11: 489. Dupont, S., B. Lundve, and M. Thorndyke. 2010. Near future ocean acidification increases growth rate of the lecithotrophic larvae and juveniles of the sea star Crossaster papposus. Journal of Experimental Zoology, Part B, Molecular and Developmental Evolution 314: 382-89. Dupré, J. 2010. The conditions of existence. [Review of John Reiss’s Not by Design.] American Scientist 98: 170. Durant, W., and A. Durant. 1968. The Lessons of History. New York: Simon and Schuster. Edgren, R. A. 1960. A seasonal change in bone density in female musk turtles, Sternothaerus odoratus (Latreille). Comparative Biochemistry and Physiology 1: 213-17. Erickson, G. M., P. J. Makovicky, P. J. Currie, M. A. Norell, S. A. Yerby, and C. A. Brochu. 2004. Gigantism and comparative life-history parameters of tyrannosaurid dinosaurs. Nature 430: 772-75. Fleagle, J. G. 1999. Primate Adaptation and Evolution. 2nd ed. New York: Academic Press. Fortey, R. 1998. Life: An Unauthorised Biography. New York: HarperCollins. Fortey, R. 2000. Trilobites! London: HarperCollins.{20} Fortey, R. 2004. Earth: An Intimate History. London: Vintage Books. Fox, D. L., and D. C. Fisher. 2001. Stable isotope ecology of a late Miocene population of Gomphotherium productus (Mammalia, Proboscidea) from Port of Entry Pit, Oklahoma, USA. Palaios 16: 279-93. Franzen, J. L. 2006. A pregnant mare with preserved placenta from the Middle Eocene maar of Eckfeld, Germany. Palaeontographica (A) 278: 27–35. Friedman, M. 2008. The evolutionary origin of flatfish asymmetry. Nature 454: 209-12. Furrer, H. 2003. Der Monte San Giorgio im Südtessin — Vom Berg der Saurier zur Fossil — Lagerstätte internationaler Bedeutung. Neujahrsblatt, Naturforschende Gesellschaft in Zürich 206: 1-64. Gallis, F. 1999. Why do almost all mammals have seven cervical vertebrae? Developmental constraints, Hox genes and cancer. Journal of Experimental Zoology, Molecular and Developmental Evolution 285: 19–26. García, V. B., L. O. Lucifora, and R. A. Myers. 2008. The importance of habitat and life history to extinction risk in sharks, skates, rays and chimaeras. Proceedings of the Royal Society of London B 275: 83–89. Garstang, W. 1922. The theory of recapitulation: A critical restatement of the biogenetic law. Journal of the Linnean Society of London, Zoology 35: 81-101. Gauthier, J. A., A. G. Kluge, and T. Rowe. 1988. Amniote phylogeny and the importance of fossils. Cladistics 4: 105–209. Gayon, J. 2000. History of the concept of allometry. American Zoologist 40: 748-58. Gee, H. 2001. Deep Time: Cladistics, the Revolution in Evolution. London: Fourth Estate. Giannini, N., A. Goswami, and M. R. Sánchez-Villagra. 2006. Development of integumentary structures in Rousettus amplexicaudatus (Mammalia: Chiroptera: Pteropodidae) during late-embryonic and fetal stages. Journal of Mammalogy 87: 993-1001. Gilbert, S. F., and D. Epel. 2008. Ecological Developmental Biology. Sunderland, MA: Sinauer Associates. Gingerich, P. D., M. ul-Haq, W. von Koenigswald, W. J. Sanders, B. H. Smith, and I. S. Zalmout. 2009. New protocetid whale from the Middle Eocene of Pakistan: Birth on land, precocial development, and sexual dimorphism. PLoS ONE 4: e4366. Göbbel, L., and R. Schultka. 2002. Das wissenschaftliche Programm von Johann Friedrich Meckel d. J. (1781–1833) und seine Bedeutung für die Entwicklung der Wissenschaft vom Leben. Annals of Anatomy 184: 519-22. Godfrey, L. R., K. E. Samonds, P. C. Wright, and S. J. King. 2005. Schultz’s unruly rule: Dental developmental sequences and schedules in small-bodied, folivorous lemurs. Folia Primatologica 6: 77–99. Goin, F. J., M. A. Abello, and L. Chornogubsky. 2010. Middle Tertiary marsupials from central Patagonia (early Oligocene of Gran Barranca): Understanding South America’s Grande Coupure. In R. H. Madden, A. A. Carlini, M. G. Vucetich, and R. F. Kay, eds., The Paleontology of Gran Barranca: Evolution and Environmental Change through the Middle Cenozoic of Patagonia (pp. 36–47). Cambridge: Cambridge University Press. Goldschmidt, R. 1933. Some aspects of evolution. Science 15: 539-47. Gompel, N., and B. Prud’homme. 2009. The causes of repeated genetic evolution. Developmental Biology 332: 36–47. Goodrich, E. S. 1913. Metameric segmentation and homology. Quarterly Journal of Microscopical Science 59: 227-48. Gostling, N. J., P. C. J. Donoghue, and S. Bengtson. 2007. The earliest fossil embryos begin to mature. Evolution and Development 9: 206-7. Gostling, N. J., C. W. Thomas, J. M. Greenwood, X. Dong, S. Bengtson, E. C. Raff, R. A. Raff, B. A. Degnan, M. Stampanoni, and P. C. J. Donoghue. 2008. Deciphering the fossil record of early bilaterian development in the light of experimental taphonomy. Evolution and Development 10: 339-49. Goswami, A. 2006. Cranial modularity shifts during mammalian evolution. American Naturalist 168: 270-80. Goswami, A., N. Milne, and S. Wroe. 2011. Biting through constraints: Cranial morphology, disparity, and convergence across living and fossil carnivorous mammals. Proceedings of the Royal Society of London B, Biological Sciences, 278: 1831-39. Gould, S. J. 1974. The origin and function of “bizarre” structures: Antler size and skull size in the “Irish elk,” Megaloceros giganteus. Evolution 28: 191–220. Gould, S. J. 1977. Ontogeny and Phylogeny. Cambridge, MA: Belknap Press. Gould, S. J. 1988. Time’s Arrow Time’s Cycle. Cambridge, MA: Harvard University Press. Gould, S. J., and N. Eldredge. 1993. Punctuated equilibrium comes of age. Nature 366: 223-27. Grande, L., and E. J. Hilton. 2006. An exquisitely preserved skeleton representing a primitive sturgeon from the upper Cretaceous Judith River formation of Montana. Journal of Palaeontology 80: 1-39. Gray, S. W. 1946. Relative growth in a phylogenetic series and in ontogenetic series of one of its members. American Journal of Science 244: 792–807. Gregory, T. R. 2005. Genome size evolution in animals. In T. R. Gregory, ed., The Evolution of the Genome (pp. 4-71). Boston: Elsevier Academic Press. Guthrie, R. D. 2003. Rapid body size decline in Alaskan Pleistocene horses before extinction. Nature 426: 169-71. Haeckel, E. 1866. Generelle Morphologie der Organismen: allgemeine Grundzüge der organischen Formen-Wissenschaft, mechanisch begründet durch die von C. Darwin reformirte Descendenz-Theorie. Berlin: Reimer. Haeckel, E. 1874. Anthropogenie oder Entwicklungsgeschichte des Menschen. Leipzig: Engelmann. Hafner, J. C., and M. S. Hafner. 1988. Heterochrony in rodents. In M. L. McKinney, ed., Heterochrony in Evolution: A Multidisciplinary Approach (pp. 217-35). New York: Plenum Press. Hall B. K. 1999. Evolutionary Developmental Biology. 2nd ed. New York: Springer. Hall B. K. 2002. Palaeontology and evolutionary developmental biology: A science of the 19th and 21st centuries. Palaeontology 45: 647-69. Hall B. K. 2005. Bones and Cartilage. Amsterdam: Elsevier Academic Press. Hammer, O., and H. Bucher. 2005. Buckman’s first law of covariation: A case of proportionality. Lethaia 38: 67–72. Harris, M. P., N. Rohner, H. Schwarz, S. Perathoner, P. Konstantinidis, and C. Nüsslein-Volhard. 2008. Zebrafish eda and edar mutants reveal conserved and ancestral roles of ectodysplasin signaling in vertebrates. PLoS Genetics 4: e1000206. Haug, J. T., A. Maas, and D. Waloszek. 2010. † Henningsmoenicaris scutula, † Sandtorpia vestrogothiensis gen. et sp. nov. and heterochronic events in early crustacean evolution. Earth and Environmental Science Transactions of the Royal Society of Edinburgh 100: 311-50. Haug, J. T., D. Waloszek, C. Haug, and A. Maas. 2010. High-level phylogenetic analysis using developmental sequences: The Cambrian †Martinssonia elongata, † Musacaris gerdgeyeri gen. et sp. nov. and their position in early crustacean evolution. Arthropod Structure and Development 39: 154-73. Head, J. J., J. I. Bloch, A. K. Hastings, J. R. Bourque, E. A. Cadena, F. A. Herrera, P. D. Polly, and C. A. Jaramillo. 2009. Giant boid snake from the Palaeocene Neotropics reveals hotter past equatorial temperatures. Nature 457: 715-17. Hendrikse, J. L., T. E. Parsons, and B. Hallgrímsson. 2007. Evolvability as the proper focus of evolutionary developmental biology. Evolution and Development 9: 393–401. Hoekstra, H. E., and J. A. Coyne 2007. The locus of evolution: Evodevo and the genetics of adaptation. Evolution 61: 995-1016. Horder, T. J. 2008. A history of evo-devo in Britain: Theoretical ideals confront biological complexity. Annals of the History and Philosophy of Biology 13: 101-74. Hou, C., W. Zuo, M. E. Moses, W. H. Woodruff, J. H. Brown, and G. B. West. 2008. Energy uptake and allocation during ontogeny. Science 322: 736-39. Houck, M. A., J. A. Gauthier, and R. E. Strauss. 1990. Allometric scaling in the earliest fossil bird, Archaeopteryx lithographica. Science 247: 195-98. Hughes, N. C. 2005. Trilobite construction: Building a bridge across the micro- and macroevolutionary divide. In D. E. G. Briggs, ed., Evolving Form and Function: Fossils and Development: Proceedings of a Symposium Honoring Adolph Seilacher for His Contributions to Paleontology, in Celebration of His 80th Birthday (pp. 139-58). New Haven: Peabody Museum of Natural History, Yale University. Humphrey, L. T. 2010. Weaning behaviour in human evolution. Seminars in Cell and Developmental Biology 21: 453-61. Irie, N., and S. Kuratani. 2011. Comparative transcriptome analysis reveals vertebrate phylotypic period during organogenesis. Nature Communications 10.Jablonski, D. 2010. Origination patterns and multilevel processes in macroevolution. In M. Pigliucci and G. B. Müller, eds., Evolution: The Extended Synthesis (pp. 335-54). Cambridge, MA: MIT Press. Jackson, J. B. C., S. Lidgard, and F. K. McKinney, eds. 2001. Evolutionary Patterns: Growth, Form, and Tempo in the Fossil Record. Chicago: University of Chicago Press. Jacob, T., E. Indriati, R. P. Soejono, K. Hsu, D. W. Frayer, R. B. Eckhardt, A. J. Kuperavage, A. Thorne, and M. Henneberg. 2006. Pygmoid Australomelanesian Homo sapiens skeletal remains from Liang Bua, Flores: Population affinities and pathological abnormalities. Proceedings of the National Academy of Sciences 103: 13421-26. Janis, C., and M. Fortelius. 1988. On the means whereby mammals achieve increased functional durability of their dentitions, with special reference to limiting factors. Biological Reviews 63: 197–230. Jenkins, F. A., and C. R. Schaff. 1988. The early Cretaceous mammal Gobiconodon (Mammalia, Triconodonta) from the Cloverly Formation in Montana. Journal of Vertebrate Paleontology 8: 1-24. Johanson, Z., A. Kearsley, J. den Blaauwen, M. Newman, and M. M. Smith. 2010. No bones about it: An enigmatic Devonian fossil reveals a new skeletal framework — A potential role of loss of gene regulation. Seminars in Cell and Developmental Biology 21: 414-23. Jolicoeur, P. 1963. The multivariate generalization of the allometry equation. Biometrics 19: 497-99. Kalinka, A. T., K. M. Varga, D. T. Gerrard, S. Preibisch, D. L. Corcoran, J. Jarrells, U. Ohler, C. M. Bergman, and P. Tomancak. 2010. Gene expression divergence recapitulates the developmental hourglass model. Nature 468: 811-14. Katsnelson, A. 2010. The bones of contention. Nature 466: 914-15. Kavanagh, K. D., A. R. Evans, and J. Jernvall. 2007. Predicting evolutionary patterns of mammalian teeth from development. Nature 449: 427-32. Keibel, F. 1906. Die Entwicklung d. äußeren Körperform d. Wirbeltierembryonen, insbes. d. menschl. Embryonen aus d. ersten 2 Monaten. In O. Hertwig, ed., Handbuch der Vergleichenden und Experimentellen Entwickelungslehre der Wirbeltiere (pp. 1-176). 1 vol., 2 pts. Jena: Fischer Verlag. Kemp, T. S. 2007a. The concept of correlated progression as the basis of a model for the evolutionary origin of major new taxa. Proceedings of the Royal Society of London B 274: 1667-73. Kemp, T. S. 2007b. The origin of higher taxa: Macroevolutionary processes, and the case of the mammals. Acta Zoologica 88: 3-22. Kielan-Jaworowska, Z., R. L. Cifelli, and Z.-X. Luo. 2004. Mammals from the Age of Dinosaurs: Origins, Evolution, and Structure. New York: Columbia University Press. Kiessling, W., M. Aberhan, and L. Villier. 2008. Phanerozoic trends in skeletal mineralogy driven by mass extinctions. Nature Geoscience 1: 527-30. Kingsley, D. M., and C. L. Peichel. 2007. The molecular genetics of evolutionary change in sticklebacks. In S. Östlund-Nilsson, I. Mayer, and F. A. Huntingford, eds., Biology of the Three-Spined Stickleback (pp. 41–81). Boca Raton, FL: CRC Press. Klein, N., T. Scheyer, and T. Tütken. 2009. Skeletochronology and isotopic analysis of a captive individual of Alligator mississippiensis Daudin, 1802. Fossil Record 12: 121-31. Klevezal, G. A. 1996. Recording Structures of Mammals: Determination of Age and Reconstruction of Life History. Rotterdam: Balkema. Klingenberg, C. P. 2010. There’s something afoot in the evolution of ontogenies. BMC Evolutionary Biology 10: 221. Klug, C. 2001. Life-cycles of Emsian and Eifelian ammonoids (Devonian). Lethaia 34: 215–33. Köhler, M., and S. Moyá-Solá. 2004. Reduction of brain and sense organs in the fossil insular bovid Myotragus. Brain, Behavior, and Evolution 63: 125-40. Kolb, C., M. R. Sánchez-Villagra, and T. M. Scheyer. 2011. The palaeohistology of the basal ichthyosaur Mixosaurus Baur, 1887 (Ichthyopterygia, Mixosauridae) from the Middle Triassic: Palaeobiological implications. Comptes Rendus Palevol 10: 403-11. Kondo, S., Y. Kuwahara, M. Kondo, K. Naruse, H. Mitani, Y. Wakamatsu, K. Ozato, S. Asakawa, N. Shimizu, and A. Shima. 2001. The medaka rs-3 locus required for scale development encodes ectodysplasin: A receptor. Current Biology 11: 1202-6. Kopper, J. S., and W. H. Waldren. 1967. Balearic prehistory: A new perspective. Archaeology 20: 108-15. Kriwet, J., W. Kiessling, and S. Klug. 2009. Diversification trajectories and evolutionary life-history traits in early sharks and batoids. Proceedings of the Royal Society of London B 276: 945-51. Kruszyński, K. J., J. A. Kaandrop, and R. van Liere. 2007. A computational method for quantifying morphological variation in scleractibnian corals. Coral Reefs 26: 831-40. Kryžanowsky, S. G. 1939. Das Rekapitulationsprinzip und die Bedingungen der historischen Auffassung der Ontogenese. Acta Zoologica 20: 1-87. Kundrát, M., A. R. I. Cruickshank, T. W. Manning, and J. Nudds. 2008. Embryos of therizinosauroid theropods from the Upper Cretaceous of China: Diagnosis and analysis of ossification patterns. Acta Zoologica 89: 231-51. Kuraku, S., and A. Meyer. 2009. The evolution and maintenance of Hox gene clusters in vertebrates and the teleost-specific genome duplication. International Journal of Developmental Biology 53: 765-73. Lalueza-Fox, C., B. Shapiro, P. Bover, J. A. Alcover, and J. Bertranpetit. 2002. Molecular phylogeny and evolution of the extinct bovid Myotragus balearicus. Molecular Phylogenetics and Evolution 25: 501-10. Larsson, H. C. E. 1998. A new method for comparing ontogenetic and phylogenetic data and its application to the evolution of the crocodylian secondary palate. Neues Jahrbuch für Geologie und Paläontologie, Abhandlungen 210: 345-68. Laurin, M. 2010. The subjective nature of Linnaean categories and its impact in evolutionary biology and biodiversity studies. Contributions to Zoology 79: 131-46. Laurin, M., and R. Reisz. 1995. A reevaluation of early amniote phylogeny. Zoological Journal of the Linnean Society 113: 165–223. Laurin, M., and R. Soler-Gijón. 2010. Osmotic tolerance and habitat of early stegocephalians: Indirect evidence from parsimony, taphonomy, paleobiogeography, physiology, and morphology. In M. Vecoli and G. Clément, eds., The Terrestrialization Process: Modelling Complex Interactions at the Biosphere-Geosphere Interface (pp. 151-79). London: Geological Society of London. Lee, M. S. Y. 2009. Hidden support from unpromising data sets strongly unites snakes with anguimorph “lizards.” Journal of Evolutionary Biology 22: 1308-16. Leroi, A. M. 2003 Mutants: On Genetic Variety and the Human Body. New York: Viking Penguin.{21} Levins, R., and R. Lewontin. 1987. The Dialectical Biologist. Cambridge, MA: Harvard University Press. Li, C., X. Wu, O. Rieppel, L. Wang, and L. Zhao. 2008. An ancestral turtle from the late Triassic of southwestern China. Nature 456: 497–501. Li, Q.-G., K.-Q. Gao, J. Vinther, M. D. Shawkey, J. A. Clarke, L. D’Alba, Q. Meng, D. E. G. Briggs, and R. O. Prum. 2010. Plumage color patterns of an extinct non-avian dinosaur. Science 327: 1369-72. Lieberman, D. E. 2009. Homo floresiensis from head to toe. Nature 459: 41–42. Lin, J.-P., S. M. Gon III, and J. G. Gehling et al. 2006. A Parvancorinalike arthropod from the Cambrian of South China. Historical Biology 18: 33–45. Lister, A. M. 1989. Rapid dwarfing of red deer on Jersey in the last Interglacial. Nature 342: 539-42. Lockley, M. G., C. A. Meyer, and V. F. Santos. 1994. Trackway evidence for a herd of juvenile sauropods from the late Jurassic of Portugal. Gaia 10: 43–48. Long, J. A. 2010. The Rise of Fishes: 500 Million Years of Evolution. 2nd ed. Baltimore: Johns Hopkins University Press. Long, J. A., and K. Trinajstic. 2010. The late Devonian Gogo Formation Lagerstatte of Western Australia — exceptional early vertebrate preservation and diversity. Annual Reviews in Earth and Planetary Sciences 38: 255-79. Long J. A., K. Trinajstic, G. C. Young, and T. Senden. 2008. Live birth in the Devonian period. Nature 453: 650-52. Losos, J. B., and R. E. Ricklefs. 2009. Adaptation and diversification on islands. Nature 457: 830-36. Luo, Z.-X. 2007. Transformation and diversification in early mammal evolution. Nature 450: 1011-19. Luo, Z.-X. 2011. Developmental patterns in Mesozoic evolution of mammal ears. Annual Review of Ecology, Evolution, and Systematics 42: 355–80. Luo, Z.-X., P. Chen, G. Li, and M. Chen. 2007. A new eutriconodont mammal and evolutionary development in early mammals. Nature 446: 288-93. Luo, Z.-X., A. W. Crompton, and A.-L. Sun. 2001. A new mammaliaform from the early Jurassic and evolution of mammalian characteristics. Science 292: 1535-40. Luo, Z.-X., Z. Kielan-Jaworowska, and R. L. Cifelli. 2004. Evolution of dental replacement in mammals. Bulletin of the Carnegie Museum of Natural History 36: 159-75. Lynch, V. J., and G. P. Wagner. 2009. Did egg-laying boas break Dollo’s law? Phylogenetic evidence for reversal to oviparity in sand boas (Eryx: Boidae). Evolution 61: 207-16. Lyson, T. R., G. S. Bever, B.-A. S. Bhullar, W. G. Joyce, and J. A. Gauthier. 2010. Transitional fossils and the origin of turtles. Biology Letters 6: 830-33. MacNamara, K. J. 1997. Shapes of Time. Baltimore: Johns Hopkins University Press. Maier, W. 1999. On the evolutionary biology of early mammals. With methodological remarks on the interaction between ontogenetic adaptation and phylogenetic transformation. Zoologische Anzeiger 238: 55–74. Maloof, A. C., C. V. Rose, R. Beach, B. M. Samuels, C. C. Calmet, D. H. Erwin, G. R. Poirier, and Nan Yao et al. 2010. Possible animal-body fossils in pre-Marinoan limestones from South Australia. Nature Geoscience 3: 653. Marjanović, D., and M. Laurin. 2009. The origin(s) of modern amphibians: A commentary. Evolutionary Biology 36: 336-38. Marroig, G., L. T. Shirai, A. Porto, F. B. de Oliveira, and V. De Conto. 2009. The evolution of modularity in the mammalian skull II: Evolutionary consequences. Evolutionary Biology 36: 136-48. Marsh, O. C. 1887. Notice of new fossil mammals. American Journal of Science 34: 323-24. Martin, T., and M. Nowotny. 2000. The docodont Haldanodon from the Guimarota coal mine. In T. Martin and B. Krebs, eds., Guimarota: A Jurassic Ecosystem (pp. 91–96). Munich: Pfeil Verlag. McFadden, B. 2005. Fossil horses: Evidence of evolution. Science 307: 1728-30. McKenna, M. C., and S. K. Bell. 1997. Classification of Mammals above the Species Level. New York: Columbia University Press. Mellett, J. S. 1969. Carnassial rotation in a fossil carnivore. American Midland Naturalist 82: 287-89. Meng, J., Y. Hu, Y. Wang, and C.- K. Li. 2003. The ossified Meckel’s cartilage and internal groove in Mesozoic mammaliaforms: Implications to origin of the definitive mammalian middle ear. Zoological Journal of the Linnean Society 138: 431-48. Meng, J., Y. Wang, and C. Li. 2011. Transitional mammalian middle ear from a new Cretaceous Jehol eutriconodontan. Nature 472: 181-85. Merks, R. M. H., A. G. Hoekstra, J. A. Kaandorp, P. M. A. Sloot, P. Hogeweg. 2006. Problem-solving environments for biological morphogenesis. Computing in Science & Engineering 8: 61–72. Mihlbachler, M. C. 2008. Species taxonomy, phylogeny, and biogeography of the Brontotheriidae (Mammalia: Perissodactyla). Bulletin of the American Museum of Natural History 311: 1-475. Mikhailov, K. E. 1991. Classification of fossil eggshells of amniotic vertebrates. Acta Palaeontologica Polonica 36: 193–238. Millien, V. 2006. Morphological evolution is accelerated among island mammals. PLoS Biology 4: e321. Minelli A. 2007. The Development of Animal Form: Ontogeny, Morphology, and Evolution. Cambridge: Cambridge University Press. Mitgutsch, C., L. Olsson, and A. Haas. 2009. Early embryogenesis in discoglossoid frogs: A study of heterochrony at different taxonomic levels. Journal of Zoological Systematics and Evolutionary Research 47: 248-57. Mivart, G. J. 1871. On the Genesis of Species. New York: Appleton. Motani, R., and H. You. 1998. The forefin of Chensaurus chaoxianensis (Ichthyosauria) shows delayed mesopodial ossification. Journal of Paleontology 72: 133-36. Müller, G. B. 2007. Evo-devo: Extending the evolutionary synthesis. Nature Reviews Genetics 8: 943-49. Müller, G. B., and S. A. Newman. 2005. The innovation triad: An evodevo agenda. Journal of Experimental Zoology, Part B, Molecular and Developmental Evolution 304: 487–503. Müller, J., T. M. Scheyer, P. Barrett, I. Werneburg, J. J. Head, P. G. P. Ericson, D. Pol, and M. R. Sánchez-Villagra. 2010. Homeotic effects, somitogenesis, and the evolution of vertebral numbers in recent and fossil amniotes. Proceedings of the National Academy of Sciences 107: 2118-23. Murdock, D. J. E., and P. C. J. Donoghue. 2011. Evolutionary origins of animal skeletal biomineralization. Cells, Tissues, Organs 194: 98–102. Myers, T. S., and A. R. Fiorillo. 2009. Evidence for gregarious behavior and age segregation in sauropod dinosaurs. Palaeogeography, Palaeoclimatology, Palaeocology 274: 96-104. Nagashima, H., F. Sugahara, M. Takechi, R. Ericsson, K. Y. Ohya, Y. Narita, S. Kuratani. 2009. Evolution of the turtle body plan by the folding and creation of new muscle connections. Science 325: 193-96. Neill, W. T. 1971. The Last of the Ruling Reptiles: Alligators, Crocodiles, and Their Kin. New York: Columbia University Press. Novacek, M. 1996. Dinosaurs of the Flaming Cliffs. New York: Anchor Books. Nowak, R. M. 1999. Walker’s Mammals of the World. Baltimore: Johns Hopkins University Press. Nützel, A., O. Lehnert, and J. Frýda. 2006. Origin of planktotrophy: Evidence from early molluscs. Evolution and Development 8: 325-30. Ogg, J. G., G. Ogg, and F. M. Gradstein. 2008. The Concise Geologic Time Scale. Cambridge: Cambridge University Press. Okada, N., Y. Takagi, T. Seikai, M. Tanaka, and M. Tagawa. 2001. Asymmetrical development of bones and soft tissues during eye migration of metamorphosing Japanese flounder, Paralichthys olivaceus. Cell and Tissue Research 304: 59–66. O’Keefe, F. R., and P. M. Sander. 1999. Paleontological paradigms and inferences of phylogenetic pattern: A case study. Paleobiology 25: 518-33. Olson, E. C. 1979. Aspects of the biology of Trimerorhachis (Amphibia, Temnospondyli). Journal of Paleontology 53: 1-17. Olson, E. C., and R. A. Miller. 1958. Morphological Integration. Chicago: University of Chicago Press. Olsson, L., G. S. Levit, and U. Hoßfeld. 2010. Evolutionary developmental biology: Its concepts and history with a focus on Russian and German contributions. Naturwissenschaften 97: 951-69. Organ, C. L., S. L. Brusatte, and K. Stein. 2009. Sauropod dinosaurs evolved moderately sized genomes unrelated to body size. Proceedings of the Royal Society of London B 276: 4303-8. Organ, C. L., A. Canoville, R. R. Reisz, and M. Laurin. 2011. Paleogenomic data suggest mammal-like genome size in the ancestral amniote and derived large genome size in amphibians. Journal of Evolutionary Biology 24: 372-80. Organ, C. L., A. M. Shedlock, A. Meade, M. Pagel, and S. V. Edwards. 2007. Origin of avian genome size and structure in non-avian dinosaurs. Nature 446: 180-84. Osborn, J., and A. W. Crompton. 1973. The evolution of mammalian from reptilian dentitions. Breviora 399: 1-18. Padian, K., and K. D. Angielczyk. 2007. “Transitional forms” versus transitional features. In A. J. Petto and L. Godfrey, eds., Scientists Confront Intelligent Design and Creationism (pp. 197–230). New York: Norton. Pagel, M. D. 1991. Constructing “everyanimal.” Review of body size in mammalian paleobiology. Edited by J. Damuth and B. J. MacFadden, Cambridge University Press, 1990. Nature 351: 532–33. Pantalacci, S., A. Chaumot, G. Benoît, A. Sadier, F. Delsuc, E. J. Douzery, and V. Laudet. 2008. Conserved features and evolutionary shifts of the EDA signaling pathway involved in vertebrate skin appendage development. Molecular Biology and Evolution 25: 912-28. Payne, J. L., D. J. Lehrmann, J. Wie, M. J. Orchard, D. P. Schrag, and A. H. Knoll. 2004. Large perturbations of the carbon cycle during recovery from the end-Permian extinction. Science 305: 506-9. Peterson, K. J., R. E. Summons, and P. C. J. Donoghue. 2007. Molecular palaeobiology. Palaeontology 50: 775–809. Ponce de León, M. S., L. Golovanova, V. Doronichev, G. Romanova, T. Akazawa, O. Kondo, H. Ishida, and C. P. E. Zollikofer. 2008. Neanderthal brain size at birth provides insights into human life history evolution. Proceedings of the National Academy of Sciences 105: 13764-68. Poplin, C., and A. J. de Ricqlès. 1970. A technique of serial sectioning for the study of undecalcified fossils. Curator 13: 6-19. Portmann, A. 1941. Die Tragzeiten der Primaten und die Dauer der Schwangerschaft beim Menschen: ein Problem der vergleichenden Biologie. Revue Suisse de Zoologie 48: 511-18. Prothero, D. 2009. Greenhouse of the Dinosaurs: Evolution, Extinction, and the Future of Our Planet. New York: Columbia University Press. Quammen, D. 1997. The Song of the Dodo: Island Biogeography in an Age of Extinction. New York: Scribner’s. Radinsky, L. 1984. Ontogeny and phylogeny in horse skull evolution. Evolution 38: 1-15. Raff, E. C., J. T. Villinski, F. R. Turner, P. Donoghue, and R. A. Raff. 2006. Experimental taphonomy shows the feasibility of fossil embryos. Proceedings of the National Academy of Sciences 103: 5846-51. Raff, R. A. 1996. The Shape of Life. Chicago: University of Chicago Press. Raff, R. A. 2007. Written in stone: Fossils, genes, and evo-devo. Nature Reviews Genetics 8: 911-20. Raup, D. M. 1961. The geometry of coiling in gastropods. Proceedings of the National Academy of Sciences 47: 602-9. Ray, S., S. Bandyopadhyay, and D. Bhawal. 2009. Growth patterns as deduced from bone microstructure of some selected neotherapsids with special emphasis on dicynodonts: Phylogenetic implications. Palaeoworld 18: 53–66. Raymond, K. R., and D. R. Prothero. 2010. Comparative variability of intermembranous and endochondral bones in Pleistocene mammals. Palaeontologia Electronica 13 (1) 4A. Reid, R. E. H. 1997. How dinosaurs grew. In J. O. Farlowand M. K. Brett-Surman, eds., The Complete Dinosaur (pp. 403-13). Bloomington: Indiana University Press. Reiss, J. O. 2009. Not by Design: Retiring Darwin’s Watchmaker. Berkeley: University of California Press. Reisz, R. R., D. Scott, H.-D. Sues, D. C. Evans, and M. A. Raath. 2005. Embryos of an early Jurassic prosauropod dinosaur and their evolutionary significance. Science 309: 761-64. Reisz, R. R., and J. J. Head. 2008. Turtle origins out to sea. Nature 456: 450-51. Reisz, R. R., and J. Müller. 2004. The fossil record and molecular time scales: A paleontological perspective. Trends in Genetics 20: 237-41. Renesto, S. 2005. A new specimen of Tanystropheus (Reptilia: Protorosauria) from the Middle Triassic of Switzerland and the ecology of the genus. Rivista Italiana di Paleontologia e Stratigrafia 111: 377-94. Richards, R. J. 1992. The Meaning of Evolution. Chicago: University of Chicago Press. Richards, R. J. 2008. The Tragic Sense of Life: Ernst Haeckel and the Struggle over Evolutionary Thought. Chicago: University of Chicago Press. Richardson, M. K., S. Gobes, A. van Leewen, A. Poelman, C. Pieau, and M. R. Sánchez-Villagra. 2009. Heterochrony in limb evolution: Developmental mechanisms and natural selection. Journal of Experimental Zoology, Molecular and Developmental Evolution 312B: 639-64. Richardson, M. K., J. Hanken, M. L. Gooneratne, C. Pieau, A. Raynaud, L. Selwood, and G. M. Wright. 1997. There is no highly conserved embryonic stage in the vertebrates: implications for current theories of evolution and development. Anatomy and Embryology (Berlin) 196: 91-106. Richardson, M. K., and G. Keuck. 2002. Haeckel’s ABC of evolution and development. Biological Reviews 77: 495–528. Richtsmeier, J. T. 2003. Growth. In B. K. Hall and W. Olson, eds., Key Concepts and Approaches in Evolutionary Developmental Biology (pp. 161-68). Cambridge, MA: Harvard University Press. Ricklefs, R. E. 2007. Tyrannosaur ageing. Biology Letters 3: 214-17. Ricqlès, A. de. 1976. On bone histology of fossil and living reptiles with comments on its functional and evolutionary significance. In A. d’A. Bellairs and C. B. Cox, eds., Morphology and Biology of Reptiles Linnean Society Symposium, ser. 3 (pp. 123-49). London: Academic Press. Ricqlès, A. de, F. J. Meunier, J. Castanet, and H. Francillon-Vieillot. 1991. Comparative microstructure of bone. In B. K. Hall, ed., Bone, vol. 3 (pp. 1-78). Boca Raton, FL: CRC Press. Rieppel, O. 2001. Turtles as hopeful monsters. BioEssays 23: 987-91. Rieppel, O., D.-Y. Jiang, N. C. Fraser, W.-C. Hao, R. Motani, Y.-L. Sun, and Z.-Y. Sun. 2010. Tanystropheus cf. T. longobardicus from the early Late Triassic of Guizhou Province, southwestern China. Journal of Vertebrate Paleontology 30: 1082-89. Rieppel, O., and M. Kearney. 2002. Similarity. Biological Journal of the Linnean Society 75: 59–82. Rivera, A. S., M. S. Pankey, D. C. Plachetzki, C. Villacorta, A. E. Syme, J. M. Serb, A. R. Omilian, and T. H. Oakley. 2010. Gene duplication and the origins of morphological complexity in pancrustacean eyes, a genomic approach. BMC Evolutionary Biology 10: 123. Rohner, N., M. Bercsényi, L. Orbán, M. E. Kolanczyk, D. Linke, M. Brand, C. Nüsslein-Volhard, and M. P. Harris. 2009. Duplication of fgfr1 permits Fgf signaling to serve as a target for selection during domestication. Current Biology 19: 1642-47. Romer, A. S. 1948. Relative growth in pelyco-saurian reptiles. In A. L. Du Toit, eds., Robert Broom Commemorative Volume. Special Publication of the Royal Society of South Africa (pp. 45–55). Romer, A. S., and L. W. Price. 1940. Review of the Pelycosauria. Geological Society of America Special Paper 28: 1-538. Roth, V. L. 1992. Inferences from allometry and fossils: Dwarfing of elephants on islands. In D. Futuyma and J. Antonovics, eds., Oxford Surveys in Evolutionary Biology (pp. 259-88). Oxford: Oxford University Press. Rougier, G. W., J. R. Wible, and M. J. Novacek. 1998. Implications of Deltatheridium specimens for early marsupial history. Nature 396: 459-63. Rowe, T. 1996. Brain heterochrony and origin of the mammalian middle ear. In M. Ghiselin and G. Pinna, eds., New Perspectives on the History of Life (pp. 71–95). Memoir 20. San Francisco: California Academy of Sciences. Rowe, T. B., T. E. Macrini, and Z.-X. Luo. 2011. Fossil evidence on origin of the mammalian brain. Science 332: 955-57. Sánchez-Villagra, M. R. 2010a. Developmental palaeontology in synapsids: The rock record of ontogeny in mammals and their closest relatives. Proceedings of the Royal Society of London B 277: 1139-47. Sánchez-Villagra, M. R. 2010b. Suture closure as a paradigm to study late growth in recent and fossil mammals: A case study with giant deer and dwarf deer skulls. Journal of Vertebrate Paleontology 30: 1895-98. Sánchez-Villagra, M. R, ed. 2010c. Developmental vertebrate palaeontology. Seminars in Cell and Developmental Biology 21: 399–470. Sánchez-Villagra, M. R. 2011. Missing Links — Zeugnisse der Vergangenheit. In U. Reyer and P. Schmid-Hempel, eds., Darwins langer Arm — Evolutionstheorie heute. Zürich: VDF Hochschulverlag. Sánchez-Villagra M. R., O. A. Aguilera, and A. A. Carlini, eds. 2010. Urumaco and Venezuelan Palaeontology: The Fossil Record of the Northern Neotropics. Bloomington: Indiana University Press. Sánchez-Villagra, M. R., A. Goswami, V. Weisbecker, O. Mock, and S. Kuratani. 2008. Conserved relative timing of cranial ossification patterns in early mammalian evolution. Evolution & Development 10: 519-30. Sánchez-Villagra, M. R., M. F. Kay, and F. Anaya-Daza. 2000. Cranial anatomy and paleobiology of the Miocene marsupial Hondalagus altiplanensis and a phylogeny of argyrolagids. Palaeontology 43: 287–301. Sander, P. M. 1988. A fossil reptile embryo from the Middle Triassic of the Alps. Science 239: 780-83. Sánchez-Villagra, M. R. 1989. The Pachypleurosaurids (Reptilia: Nothosauria) from the Middle Triassic of Monte San Giorgio (Switzerland) with the description of a new species. Philosophical Transactions of the Royal Society of London B, Biological Sciences 325: 561–666. Sánchez-Villagra, M. R. 1994. Reptilien. Stuttgart: Enke. Sander, P. M., A. Christian, M. Clauss, R. Fechner, C. T. Gee, E.-M. Griebeler, H.-C. Gunga, J. Hummel, H. Mallison, S. F. Perry, H. Preuschoft, O. W. M. Rauhut, K. Remes, T. Tütken, O. Wings, and U. Witzel. 2010. Biology of the sauropod dinosaurs: The evolution of gigantism. Biological Reviews 86: 117-55. Sander, P. M., and N. Klein. 2005. Developmental plasticity in the life history of a prosauropod dinosaur. Science 310: 1800–1802. Sansom, R., S. Gabbott, and M. Purnell. 2010. Non-random decay of chordate characters causes bias in fossil interpretation. Nature 463: 797–800. Scheyer, T. M. 2007. Skeletal histology of the armor of the Placodontia: The occurrence of “postcranial fibro-cartilaginous bone” and its developmental implications. Journal of Anatomy 211: 737-53. Scheyer, T. M., N. Klein, and P. M. Sander. 2010. Developmental palaeontology of Reptilia as revealed by histological studies. Seminars in Cell and Developmental Biology 21: 462-70. Scheyer, T. M., and M. R. Sánchez-Villagra. 2007. Carapace bone histology in the giant pleurodiran turtle Stupendemys geographicus: Phylogeny and function. Acta Palaeontologica Polonica 52: 137-54. Schmid, L., and M. R. Sánchez-Villagra. 2011. The potential genetic bases of morphological evolution in the Triassic fish Saurichthys. Journal of Experimental Zoology, Molecular and Developmental Evolution 314B: 519-26. Schoch, R. R. 2009. Life-cycle evolution as response to diverse lake habitats in Paleozoic amphibians. Evolution 63: 2738-49. Schoch, R. R. 2010. A fresh look at an early theory of evolution and development: A new translation of Schmalhausen’s Factors of Evolution. Acta Zoologica 91: 495-96. Schöne, B. R., S. D. Houk, A. D. F. Castro, J. Fiebig, W. Oschmann, I. Kroncke, W. Dreyer, and F. Gosselck. 2005. Daily growth rates in shells of Arctica islandica: Assessing sub-seasonal environmental controls on a long-lived bivalve mollusk. Palaios 20: 78–92. Schulte, P. L., I. Alegret, and J. A. Arenillas et al. 2010. The Chicxulub asteroid impact and mass extinction at the Cretaceous-Paleogene boundary. Science 327: 1214-18. Schwartz, J. H. 1999. Sudden Origins: Fossils, Genes, and the Emergence of Species. New York: John Wiley & Sons. Schwartz, G. T., L. R. Godfrey, and P. Mahoney. 2007. Inferring primate growth, development, and life history from dental microstructure: The case of the extinct giant Malagasy lemur, Megaladapis. In S. E. Bailey and J.-J. Hublin, eds., Dental Perspectives on Human Evolution: State of the Art Research in Dental Anthropology (pp. 147-62). New York: Springer. Schweitzer, M. H., R. M. Elsey, C. G. Dacke, J. R. Horner, and E. T. Lamm. 2007. Do egglaying crocodilian (Alligator mississippiensis) archosaurs form medullary bone? Bone 40: 1152-58. Sears, K. E., R. R. Behringer, J. J. Rasweiler, and L. A. Niswander. 2006. Development of bat flight: Morphologic and molecular evolution of bat wing digits. Proceedings of the National Academy of Sciences 103: 6581-86. Sewertzoff, A. N. 1927. Über die Beziehung zwischen der Ontogenese und der Phylogenese der Tiere. Jena Zeitschrift der Naturwissenschaften 63: 51-180. Sharman, G. B. 1970. Reproductive physiology of marsupials. Science 167: 1221-28. Sholtis, S., and K. Weiss. 2005. Phenogenetics: Genotype — phenotype interactions and variation. In B. Hallgrimsson and B. K. Hall, eds., Variation (pp. 499–523). New York: Academic Press. Shubin, N. 2008. Your Inner Fish: A Journey into the 3.5-Billion-Year History of the Human Body. New York: Pantheon Books.{22} Shubin, N., C. Tabin, and S. Carroll. 2009. Deep homology and the origins of evolutionary novelty. Nature 457: 818-23. Sigurdsen, T., and D. M. Green. 2011. The origin of modern amphibians: A re-evaluation. Zoological Journal of the Linnean Society 162: 457-69. Simpson, G. G. 1945. The principles of classification and a classification of mammals. Bulletin of the American Museum of Natural History 85: 1-350. Simpson, G. G. 1953. Major Features of Evolution. New York: Columbia University Press. Simpson, G. G. 1970. The Argyrolagidae, extinct South American marsupials. Bulletin of the Museum of Comparative Zoology 139: 1-86. Sire, J., P. C. J. Donoghue, and M. K. Vickaryous. 2009. Origin and evolution of the integumentary skeleton in non-tetrapod vertebrates. Journal of Anatomy 214: 409-40. Smith, A. B. 1998. What does palaeontology contribute to systematics in a molecular world? Molecular Phylogenetics and Evolution 9: 437-47. Smith, B. H. 1992. Life history and evolution of human maturation. Evolutionary Anthropology 1: 134-42. Smith, T. 2006. The Real Rudolph: A Natural History of Reindeer. Phoenix Mill, UK: Sutton. Smith, T. M., P. Tafforeau, D. J. Reid, R. Grün, S. Eggins, M. Boutakiout, and J. J. Hublin. 2007. Earliest evidence of modern human life history in North African early Homo sapiens. Proceedings of the National Academy of Sciences 104: 6128-33. Sober, E. 2008. Evidence and Evolution: The Logic behind the Science. Cambridge: Cambridge University Press. Sollas, W. J., and I. B. J. Sollas. 1903. An account of the Devonian fish Palaeospondylus gunni Traquair. Philosophical Transactions of the Royal Society of London 196: 267-94. Stein, K., Z. Csiki, K. Curry Rogers, D. B. Weishampel, R. Redelstorff, J. L. Carballido, and P. M. Sander. 2010. Small body size and extreme cortical bone remodeling indicate phyletic dwarfism in Magyarosaurus dacus (Sauropoda: Titanosauria). Proceedings of the National Academy of Sciences 107: 9258-63. Stern, D. L., and V. Orgogozo. 2009. Is genetic evolution predictable? Science 323: 746-51. Steyer, J. S., and R. Damiani. 2005. A giant brachyopoid temnospondyl from the Upper Triassic or Lower Jurassic of Lesotho. Bulletin de la Société Géologique de France 176: 243-48. Stolarski J., A. Meibom, R. Przeniosto, and M. Mazur. 2007. A Cretaceous scleractinian coral with a calcitic skeleton. Science 318: 92–94. Surget-Groba, Y., B. Heulin, C.-P. Guillaume, M. Puky, B. Smajda, D. Semenov, V. Orlova, L. Kupriyanova, and I. Ghira. 2006. Multiple origins of viviparity, or reversal from viviparity to oviparity? The European common lizard (Zootoca vivipara, Lacertidae) and the evolution of parity. Biological Journal of the Linnean Society 87: 1-11. Suzuki, H. K. 1963. Studies on the osseous system of the slider turtle. Annals of the New York Academy of Sciences 109: 351–410. Taquet, P. 2006. Georges Cuvier: Naissance d’un génie. Paris: Odile Jacob. Thewissen J. G., M. J. Cohn, L. S. Stevens, S. Bajpai, J. Heyning, and W. E. Horton Jr. 2006. Developmental basis for hind limb loss in dolphins and origin of the cetacean body plan. Proceedings of the National Academy of Sciences 103: 8414-18. Thewissen, J. G. M., L. N. Cooper, J. C. George, and S. Bajpai. 2009. From land to water: The origin of whales, dolphins, and porpoises. Evo-Devo Outreach 2: 272-88. Thewissen, J. G. M., and W. A. McLellan 2009. Maiacetus: Displaced fetus or last meal? PloS ONE 4: e4366. Tischlinger, H., and R. Wild. 2009. Den Schwanz verloren-das Leben gerettet! Fossilien 4: 203-9. Tschanz, K. 1988. Allometry and heterochrony in the growth of the neck of Triassic prolacertiform reptiles. Palaeontology 31: 997-1011. Turner S., C. J. Burrow, H.-P. Schultze, A. Blieck, W.-E. Reif, C. B. Rexroad, P. Bultynck, and G. S. Nowlan. 2010. False teeth: Conodont- vertebrate phylogenetic relationships revisited. Geodiversitas 32: 545-94. Turvey, S. T., O. R. Green, and R. N. Holdaway. 2005. Cortical growth marks reveal extended juvenile development in New Zealand moa. Nature 435: 940-43. Uhen, M. D. 2000. Replacement of deciduous first premolars and dental eruption in archaeocete whales. Journal of Mammalogy 81: 123-33. Ungar, P. S. 2010. Mammal Teeth: Origin, Evolution and Diversity. Baltimore: Johns Hopkins University Press. Urdy, S., N. Goudemand, H. Bucher, and R. Chirat. 2010. Allometries and the morphogenesis of the molluscan shell: A quantitative and theoretical model. Journal of Experimental Zoolog y, Molecular and Developmental Evolution 314B: 208–302. Van der Geer, A., G. Lyras, J. de Vos, and M. Dermitzakis. 2010. Evolution of Island Mammals: Adaptation and Extinction of Placental Mammals on Islands. Oxford: Wiley-Blackwell. van Nievelt, A. F. H., and K. K. Smith. 2005. To replace or not to replace: The significance of reduced tooth replacement in marsupial and placental mammals. Paleobiology 31: 324-46. Van Valen, L. M. 1973. An evolutionary law. Evolutionary Theory 1: 1-30. Vinogradov, A. E., and O. V. Anatskaya. 2006. Genome size and metabolic intensity in tetrapods. Proceedings of the Royal Society of London B 273: 27–32. Vrba, E. S. 1996. Climate, heterochrony, and human evolution. Journal of Anthropological Research 52: 1-28. Vrba, E. S. 2004. Ecology, evolution, and development: Perspectives from the fossil record. In B. K. Hall, R. D. Pearson, and G. B. Müller, eds., Environment, Development, and Evolution: Towards a Synthesis (pp. 85-105). Cambridge, MA: MIT Press. Vrba, E. S. 2005. Mass turnover and heterochrony events in response to physical change. Paleobiology 31: 157-74. Wägele, J.-W. 2005. Foundations of Phlogenetic Systematics. Munich: Pfeil Verlag. Wagner, A. 2011. The origins of evolutionary innovations. Oxford: Oxford University Press. Wagner, G. P., G. Booth, and H. Bagheri. 1997. A population genetic theory of canalization. Evolution 51: 329-47. Wang, S. C., and P. Dodson. 2006. Estimating the diversity of dinosaurs. Proceedings of the National Academy of Sciences 103: 13601-5. Weiss K. M., and A. V. Buchanan. 2009. The Mermaid’s Tale: Four Billion Years of Cooperation in the Making of Living Things. Cambridge, MA: Harvard University Press. Werneburg, I. 2009. A standard system to study vertebrate embryos. PLoS ONE 4: e5887. Werneburg, I., and M. R. Sánchez-Villagra. 2009. Time of organogenesis supports basal sauropsid position of turtles in the amniote tree of life. BMC Evolutionary Biology 9: 82. Werneburg, R. 2009. The Permotriassic branchiosaurid Tungussogyrinus Efremov, 1939 (Temnospondyli, Dissorophoidea), from Siberia restudied. Fossil Record 12: 105-20. Westheide W., and R. Rieger, eds. 2009. Spezielle Zoologie. Teil 2: Wirbeltiere. 2nd ed. Heidelberg: Spektrum. Weston, E. M., and A. M. Lister. 2009. Insular dwarfism in hippos and a model for brain size reduction in Homo floresiensis. Nature 459: 86–88. Wilcken, P. 2010. Claude Lévi-Strauss: The Poet in the Laboratory. London: Bloomsbury. Wild, R. 1974. E. Kuhn-Schnyder und B. Peyer: Die Triasfauna der Tessiner Kalkalpen. XXIII. Tanystropheus longobardicus (Bassani) (Neue Ergebnisse). Schweizerische Paläontologische Abhandlungen 95: 1-162. Wilson, L. A. B., and M. R. Sánchez-Villagra. 2010. Diversity trends and their ontogenetic basis: An exploration of allometric disparity in rodents. Proceedings of the Royal Society of London B 277: 1227-34. Wissler, L., H. Funk, and H. Weissert. 2003. Response of early Cretaceous carbonate platforms to changes in atmospheric carbon dioxide levels. Palaeogeography, Palaeoclimatology, Palaeoecology 200: 187–205. Witmer, L. M. 1995. The extant phylogenetic bracket and the importance of reconstructing soft tissues in fossils. In J. J. Thomason, ed., Functional Morphology in Paleontology (pp. 19–33). Cambridge: Cambridge University Press. Witten, P. E., and A. Huysseune. 2009. A comparative view on mechanisms and functions of skeletal remodelling in teleost fish, with special emphasis on osteoclasts and their function. Biological Reviews 84: 315-46. Witzmann, F. 2009. Cannibalism in a small growth stage of the early Permian branchiosaurid Apateon gracilis (Credner, 1881) from Saxony. Fossil Record 12: 7-11. Woltereck, R. 1909. Weitere experimentelle Untersuchungen über Artveränderungen, speziell über das Wesen quantitativer Artunterschiede bei Daphniden. Verhandlungen der Deutschen Zoologischen Gesellschaft 19: 110-72. Xiao-Chun, W., L. Zhan, Z. Bao-Chun, and D. Zhi-Ming. 2003. A polydactylous amniote from the Triassic period. Nature 426: 516. Zeller, U. 1999. Marsupial reproduction: Origin and evolutionary transformations. Zoologischer Anzeiger 238: 117-30. Zhang, F., S. L. Kearns, P. J. Orr, M. J. Benton, Z. Zhou, D. Johnson, X. Xu, and X. Wang. 2010. Fossilized melanosomes and the colour of Cretaceous dinosaurs and birds. Nature 463: 1075-78. Zimmerman, W. 1968. Evolution und Naturphilosophie. Berlin: Duncker & Humblot. Zollikofer, C. P. E., and M. S. Ponce de León. 2010. The evolution of hominid ontogenies. Seminars in Cell and Developmental Biology 21: 432-40. Zug, G. R., L. J. Vitt, and J. P. Caldwell. 2001. Herpetology: An Introductory Biology of Amphibians and Reptiles. 2nd ed. New York: Academic Press. Zylberberg, L., F. J. Meunier, and M. Laurin. 2010. A microanatomical and histological study of the postcranial dermal skeleton in the Devonian sarcopterygian Eusthenopteron foordi. Acta Palaeontologica Polonica 55: 459-70.Предметный указатель
Австралия, 7, 51, 77, 112 автотомия хвоста, 44-45 Агассиц, Луи, 12-13 адаптации, 3, 77, 81, 90, 112, 140; — к водной среде обитания, 83, 154; — к плаванию, 53; — к ферментации в кишечнике, 43; — параллельные генотипические, 3; — у островных видов, 120–124; — экологические, 123, 132 адаптивная радиация, 8, 98 аксиальная бифуркация, 12, 13рис. акулы. См. хрящевые рыбы аллометрия, 43, 95, 97, 98, 99, 102рис.,168, 187; — в процессе роста, 96рис., 97–99, 119, 148, 160; — внутривидовая, 102рис.; — межвидовая, 102рис.; — птенцовый тип потомства, 47–48, 56, 60–61, 167, — у Alphadon, 167 Аляска, США, 112 Американский Музей Естественной Истории, 28, 57, 58рис. аммониты, 33, 108, 178, 184; — аммонителла, 185, 187рис.; — Бакмана правило, 186рис.; — онтогенез, 185, 187рис.; — скульптурирование раковины, 184-87 амниотическая жидкость, 55 Амприно правило, 71 Амприно, Родольфо, 71 амфибии, 21–23, 44, 59, 99, 163, 199 п.12. См. также лягушки и саламандры; — безногие, 21; — лепоспондилы, 22 анапсидный череп, 151 анатомия, 4, 10, 16, 40, 43, 55, 60; — жевательного аппарата, 22; — ископаемых хордовых, 62; — летучих мышей, 147; — на момент рождения, 29, 48; — панциря черепахи, 150–154, 153рис.; — плода, 47; уха, 155; — челюсти млекопитающего, 16–18, 18рис. Англия, 14, 17, 182 Анды, 7 Антарктический полуостров, 190 апикальный эктодермальный гребень, 156 арабский песчаный удавчик. См. Eryx арагонитовый скелет, 183 Аргентина, 114 аристогенез, 28 атомизация, 32, 33 Африка, 17, 113, 118. См. также Южная Африка Ашер, Роб, 131Бавария, Германия, 28, 45, 191 Базель, Швейцария, 67, 135, 170 Бакмана закон, 185, 186рис. бактерии, 106 Балеарские острова, 122, 124 Бали, 118, батоидеи. См. хрящевые рыбы бедренная кость, 69, 74, 156рис. безымянная кость. См. таз беременность, 13, 47, 56, 60, 61, 73–74, 78, 116, 167, 170, 203-4 Беркли, США, 144, Берлин, Германия, 45, беспозвоночные, 12, 41, 67, 105, 109, 110, 176, 182, 189, 190. См. также иглокожие, моллюски и насекомые биокальцификации кризис, 109 биологическое разнообразие, 2, 3, 5, 34, 66, 108, 196 биоминерализация, 182-84 близнецы: ископаемые, 60 Блумберг, Марк С., 25, 36 Боливийское Альтиплано, 167 Бонн, 69, 76 Борель, Эмиль, 2 Бота-Бринк, Дженифер, 117 Ботанический сад, Париж, 10 бранхиозавры, 23 Брейяр, Арно, 109 Бразилия, 191 Бриггс, Дерек, 191 «Британская энциклопедия», книга, 1-2 бронтотерии, 97, 98, 99, 104 брюхоногие моллюски, 111, 184, 188 Бюше, Хьюго, 33, 185
Вагнер, Гюнтер, 9 Вайссерт, Хельмут, 109 Везувий, 107 Великая Цепь Бытия. См. Scala naturae Венесуэла, 69, 70 верблюды, 39 Вернебург, Ингмар, 23 Вернебург, Ральф, 23 веслоногие ракообразные, 109 Ветерби, Скотт, 149 внутреннее оплодотворение, 51 водные рептилии. См. рептилии водоросли, 182 волосы/шерсть, 7–9, 19, 47, 87, 138, 210 восстановление после массового вымирания, 108, 117
Гавайи, 118-19 галапагосская морская игуана, 81–82, 82рис.; — зубы, 87, 89, 112, 161, 173-74; — минимальная оценка возраста, 100; — оценка возраста половозрелости, 81, 83рис.; — срезы костей, 66–67, 69, 71рис., 83рис., 84, 133 Галапагосские острова, 54, 119; — вьюрки, 118; — морская игуана, 54, 78-81 Галле, Германия, 11 Гамбургер, Виктор, 144 Гарвардский Университет, 17, 198 Геккель, Эрнст, 13–14, 19, 21, 31, 198 Гексли, Томас, 1, 133 генетика развития, 34, 126-40, 149, 157 генотипы, 3 геометрическая морфометрия, 94, 188. См. также рост: геометрический подход геохимия, 2, 109 Германия, 21, 23, 45, 144, 150, 203 гетеротопия, 19, 20рис. гетерохрония, 19, 20рис. гетероцеркальный хвост, 13 Гилберт, Скотт, 35 гиппопотам, 99, 154; ископаемый, 119, 120, 123-24 гипсодонтия, 114 Гис, Вильгельм, 67, 203 гистология, 49, 56, 66–91, 104, 201, 203. См. также палеогистология; — гистология костей, 68, 69, 71, 72рис., 76–81, 82рис., 83рис. 103, 117, 122, 127, 161-62; глаза, 3, 34–35, 47–48, 62, 99, 120, 143-44, 146рис., 160, 190-91, 205, — сложные, 191 глазница, 42, 120, 145рис. гоацин. См. птицы: выводковые Гоби, пустыня, 57 Гого, формация, 51, 202 Гоин, Панчо, 113 головной мозг, 16, 31, 99, 123, 124, 168; — дифференциация, 48; — размер, 123, 160, 169рис., 171; — человеческий, 170-72 головоногие моллюски, 184 гологенез, 28, 29рис. Гольдшмидт, Ричард, 144-47, 160 гоминиды, 27, 123. См. также крупные человекообразные обезьяны; — летопись окаменелостей, 167–170, 169рис. гомойотермия, 84, 163, 164рис. гомология, 42, 87, 128 гомункулюсы, 92 Гондвана, 69 Госвами, Анджали, 42 глубинная гомология, 128 «глубокое время», 2–3, 106, 159рис., 191, 197 граптолиты, 67 Грузия, 168 грызуны, 47, 103, 119. См. также капибары, морские свинки и хомяки Гудрич, Э. С., 44 Гулд, Стивен Джей, 170-71
данио-рерио, 137, 139рис., 140, 199n15 Дарвин, Чарльз, 2–3, 7, 10, 14, 32, 40, 124, 207 Дарт, Раймонд, 168 двустворчатые моллюски, 189-90 двусторонняя симметрия, 33, 81 де Рикле, Арман., 68, 74-75 девонский период, девон, 6рис., 51, 67, 85, 135, 177, 191, 203, 209 Деканские траппы, 107-8, 207 дельфины, 155-56 дентин, 86–88, 205 Джерси, Соединённое Королевство, 119 Джинджерич, Фил, 154 дивергенции время: оценка, 5 динамика клады, 105 динозавры, 4, 46–59, 74–77, 104, 107, 127, 127рис., 133, 148, 159, 178, 202, 208. См. также Oviraptor, Protoceratops, зауроподы, прозауроподы и тероподы; пернатые, 11, 19, 142, 198. дифиодонтия, 161, 164рис. дицинодонты, 117 длинноволновое ультрафиолетовое излучение, 45 длинные кости, 69, 73–75, 78, 82рис., 122. См. также бедренная кость, большеберцовая кость, малоберцовая кость и плечевая кость Дманиси, Грузия, 168 ДНК, 5 добавление признаков на конечных стадиях развития, 19, 20рис. Донахью, Фил, xiii дрейф континентов, 7 Дувр, Англия, 182 дупликация гена, 84, 138, 204 дюгони, 131 Дюпон, Сэм, 109 Дюран, Ариэль, 1 Дюран, Уилл, 1 Дюрер, Альбрехт. 94
Евразия, 17 Европа, 14, 108, 147, 198 Египет, 154 естественный отбор, 26, 29, 36, 112 ехидны, 51, 159рис., 166,
жабры, 23, 59 желудка содержимое, 22, 37 живорождение, 50–56, 79, 80, 116, 135, 167 жирафы, 87, 130, 154
закисление океана, 109 зауроподы, 48, 50, 58, 69, 75–77, 133, 204. См. также Apatosaurus, Diplodocus и Mamenchisaurus зауроптеригии, 52–53. См. также пахиплеврозавры, плакодонты и плезиозавры златокроты, 131, 208 змеи, 12, 25, 51–54, 58, 80, 115-16, 129, 179, 202. См. также Eryx, Sanajeh и морские змеи Золенхофен, 191 Золликофер, Кристоф, 168, 175 зубная кость, 15-17 зубы, 7, 14–15, 22, 34, 35, 49, 59, 61, 98–99, 103, 114, 117, 135, 140, 151, 161, 173-74. См. также дентин; — бугры, 41; — гистология, 85–89, 112; — замещение, 42, 88рис., 161, 167, 174рис.; — зубная формула, 161; — износ, 47, 89–91, 114; — клыки, 40; — коренные, 40, 170, 173, 174рис.; — окклюзия, 161, 164рис., 165рис.; — развитие, 86, 131, 138-40, 205n15; — режущие, 91; — характеристики, 41;
иглокожие, 33. См. также Crossaster и морские ежи известняк, 182 изометрия, 95, 97, 99 Индийский субконтинент, 107, 154 Индия, 58 Индонезия, 123 «инструментальные белки», 138, 140 интеграция, 8, 40–43, 184 Ирландия, 89 история жизни, 29–32, 47, 49, 61, 71, 73, 77, 78, 81, 87–89, 90, 106-19, 124, 134, 158, 160-74, 177, 183, 188, 189, 201, 207 Италия, 144 ихтиозавры, 52–53, 79, 101, 204, 206
Йена, Германия, 21 Йернвалл, Юкка, 36, 41, 86
К-стратегия, 78 кайнозой, 6рис., 90 какапо, 78, 204 кальмары, 79, 184 кальций, 73–74; соли 182 кальцитный скелет, 178, 183 кальция карбонат, 64, 74, 108-9, 183 кальция фосфат, 64 камбалы, 143-47, 146рис., 160 каменноугольный период, карбон 6рис., 21, 97 канальцы, 70 каннибализм, 59 капибары, 101, 103 Карибские острова, 118 Карлини, Фреди, 113 карпы, 135, 137, 139рис. Карру, Южная Африка, 19, 198 Картахена, Колумбия, 116 Кастанье, Жак, 75 кембрий, 6, 63, 64, 110, 111, 178, 191 кенгуру, 5, 112, 143, 158, 159рис. Кемп, Том, 163-64 Кёлер, Мейке, 124 Килмейер, Карл Фридрих, 14 Кипр, 120 Китай, 11, 49, 63, 76, 130, 199 китообразные. См. дельфины и киты киты, 52, 61, 132, 147, 158, 179; — эволюция задней конечности, 154-57, 156рис., кладистическая методология, 180 классификация, 28, 32, 56, 198 климатические изменения, 106, 107, 112-16, 115рис., 202 Клингенберг, Крис, 41 коалы, 51, 130 Кобе, Япония, 130 ковариация, 40 кокколитофоры, 182 кокколиты, 182 коленные чашечки, 39-40 коллаген, 70, 182, 203 Колумбия, 115-16 колюшки, 136-37, 139рис., 209 комодосский варан, 77 компьютерная томография, 49, 51, 68, 160 конечности, 12, 23, 34–35, 43–44, 57, 61, 79, 81, 112, 113рис., 147-48, 154-57, 163, 207. См. также руки, ноги и длинные кости конодонты, 85, 111, 205 кооперативное размножение, 171 копытные, 60, 75, 87, 88, 130, 148, 154. См. также олени кораллы, 108, 177, 183, 188 коровы, 75, 87, 148, 154 коррелированная прогрессия, 163, 164рис.,165 костный морфогенетический белок, 149 кость — васкулярная, 70, 74, 75; — губчатая, 70; — компактная, 70; — компактность, 82, 83рис.; — компактная костная ткань, 75, 80, 82рис., 122; — ламеллярная, 72рис., 81, 122, 127; — медуллярная, 74; — микроструктура, 70–74, 72 рис. См. также гистология; — нарастание, 69, 80, 81; — отложение, 80; — перестройка вещества, 69, 73, 81, 83рис., 88; — резорбция, 68, 73, 80, 81, 88; — реконструкция, 68; — рыхлая волокнистая, 81; — следы процесса роста, 70–72, 81; — фиброламеллярная, 72рис., 75, 127, 128, 162, 164рис.; кости, 66–91, 104, 170, 182. См. также длинные кости и челюсти; — дермальные, 134, 135, 138, 140; — мембранные, 40; — минерализация, 80; — полости для костного мозга, 74, 80, 82; — энхондральные, 40 кошачьи, 99. См. также кошки и Smilodon fatalis кошачьи акулы. См. хрящевые рыбы кошки, 148. См. также Smilodon fatalis кровеносные сосуды. См. микроструктура костей крабы, 176 краниум. См. череп Крато, формация, 190 Кривет, Юрген, 116 Крит, 120 крокодилы, 20, 45, 51, 54, 56, 73, 122, 127, 127рис., 151, 155, 161, 202 Кромптон, А. У., 17, 198 Кружински, Крис, 188 крупные человекообразные обезьяны, 168, 170-71, 195 КТ. См. компьютерная томография Куба, 118 Куратани, Шигеру, 130, 199–200, 208 Кэрролл, Шон, 57, 128 Кювье, Жорж, 10–11, 26, 85, 198
Ла-Манш, 182 Ламарк, Жан-Батист, 10, 28 Ла Плата, Аргентина, 113 Лейпциг, Германия, 172 Лестер, Англия, 62 лемуры, 7 ленивцы, 39, 60, 110, 206. См. также Paramylodon и Nothrotheriops лепоспондилы: онтогенез, 22 летучие мыши, эволюция крыльев, 147-50 Лехманн, Томас, 131, 208 ЛЗР. См. линии задержки роста Ли, Чун, 151 линии задержки роста, 71–72, 78, 82рис., 83рис. линька, 33 Листер, Адриан, 119, 123 личинки, 21–23, 32, 37, 109, 110-12; — морских ежей, 64 локомоция, 44, 78,-80, 112, 148, 158 Лос Анджелес, США, 39 лось, 75 лошади, 75, 97–99, 148. См. также Equus; Лугано, 135 люди, 1, 4, 7, 9, 11, 27, 31, 35, 39, 44, 46, 49, 73–74, 78, 94, 99, 118, 122, 128, 141-43, 151, 195-96, 208. См. также Homo; — позвоночный столб, 129, 132; — развитие, 14, 15рис., 24рис., 25, 158-75, 159рис., 169рис., 172, 208; — ребёнок из Таунга, 168; — череп; 11, 61, 171-72 лягушки, 21, 22рис., 25, 38, 59, 200
Мадагаскар, 7, 49–50, 123 Майер, Вольфганг, 29 макроэволюция, 105, 142-43 МакФи, Джон, 2 малоберцовая кость, 49, 73 Мальта, 120 мамонты, 60, 88, 119 Марокко, 174 мезенхима, 149 мезозавры, 7, 45 мезозой, 6рис., 52, 143, 199 Мексика, 39 мел, 182 меловой период, мел, 6рис., 8, 11, 21, 49, 52, 57, 58, 77, 97, 135, 147, 159, 162, 167, 182, 183, 191, 209 Мессель, Германия, 150 метаморфоз, 23, 110, 127 микротом, 67–68, 203 Миллер, Роберт, 41 минога, 62 Митгатч, Кристиан, 21 млекопитающие, 4, 7–8, 11, 14–19, 17рис., 18рис., 29, 39, 41–43, 47, 51–52, 59–61, 85–90, 100, 102рис., 112-14, 113рис., 115рис., 117-23, 138, 148, 151, 154-56, 165рис., 198, 202, 205-8, 210-11. См. также сумчатые, однопроходные и плацентарные; — гистология, 72рис., 74–76; — количество позвонков, 130-33; — модульность черепа, 42–43; — отделение от ветви рептилий, 158; — предковые млекопитающие, 159рис., 161; — последний общий предок ныне живущих, 142-43; — прорезывание зубов, 174; — развитие, 25, 158-75; — рост многообразия, 117; — среднее ухо. См. ухо; — уникальные особенности, 7 моа, 77-78 модульность, 34–45, 200; — в черепе млекопитающих, 42 мозазавриды, 52–53, 202 Мойя-Сола, Сальвадор, 124 молекулярная биология, 5 моллюски, 7, 32–33, 87, 108, 182, 184-85, 189. См. также аммониты, брахиоподы, головоногие моллюски, двустворчатые моллюски, брюхоногие моллюски, улитки и кальмары молочное вскармивание, 74, 161, 164рис., 173, 210 Монголия, 57 Монне, Клод, 188 Монте Сан Джорджио, 79, 134, 204 морские ежи, 33, 109, 177 морские жёлуди, 32 морские змеи, 54 морские свинки, 47 морфогенез, 149, 184 морфология, 12, 20, 25, 27, 28, 31–32, 34, 38, 41–43, 50–54, 84, 87, 89, 98–99, 110, 112, 114, 119, 121, 124, 134, 136, 138, 140, 143, 148, 152, 155, 170, 180, 182, 184, 186, 88, 191, 198–200, 202, 204, 207-8 морфометрия, 33, 94, 100–101, 188, 204, 206 Музей Естественной Истории, Лондон, 203 Музей Естествознания, Париж, 10 мутации, 3, 35, 86, 132, 137-38, 140, 145 мыши, 73, 86, 119, 130, 141, 148-49, 164, 199 мышцы, 9, 18, 31, 39–40, 129-30, 152, 153рис., 165рис. мышь. См. мыши Мэдден, Рик, 114 Мюллер, Йоханнес, 132,
Нагашима, Хироши, 152, 153рис. Нариокотоме, окрестности озера Туркана, Кения, 168 Нарита, Юичи, 130 насекомые, 164, 176, 179 насиживание, 48, 56-59 неандертальцы, 169рис., 170-72, постнатальный онтогенез, 172 недостающие звенья, 141-57, 209; — в эволюции глаза камбалы, 143-47; — в эволюции задней конечности кита, 154-57; — в эволюции крыла летучей мыши, 147-50; — в эволюции панциря черепахи, 150-54; — между рептилиями и птицами, 28 нелетающий, 77–78, 201 неоген, 6рис., 112, 203 нервный гребень, 21, 22рис., 86, 205 нёбо, 164рис., 165рис., 166 Новачек, Майк, 57 Новая Зеландия, 77-78 ноги, 9, 48–49, 57, 61, 69, 73, 76, 149, 154, 155. См. также конечности Норрелл, Марк, 57 нос, 120, 165рис., 166 носовая полость. См. нос носороги, 94 нотохорд, 62-63 Нутцель, Александр, 111 Нуэво Леон, Мексика, 39 Нью-Йорк, США, 28, 57
«О росте и форме», книга, 93, 94рис., 206 однопроходные, 131, 143, 159рис., 166-67. См. также ехидны и утконос озёра, 22, 37–38, 118, 122; Виктория, 118 окостенение, 48–49, 100, 151, 199, 204 Оксфорд, Англия, 67 Оксфордский Университет, 144, 163 олени, 87–89, 96рис., 101, 103, 119-20, 154, 205 оленьи рога, 89, 96рис., 101-3, 103рис. олигоцен, 60, 114-15, 115рис. Олсон, Эверетт, 41, 203 Олссон, Леннарт, 21, 201 омары, 176 онтогенез, 1-33, 20рис., 29рис., 30рис., 37, 38, 74, 76, 87, 92, 93, 96, 100, 114, 123, 144, 160, 171, 172, 191; — аммонитов, 185-88, 187рис.; — аспекты, 92; — в ископаемом состоянии, 46–65; — реконструкция, 126; — у бранхиозавров, 23; — у темноспондилов, 21-22 онтогенетическая траектория, 32, 95, 169рис. опоссум, 130 ордовикский период, ордовик, 6рис., 110-11, 191 ореодонты, 60, 97 «орстенские» местонахождения, 191 ортогенез, 28 Осборн, Генри Фейерфилд, 28, 57 осетрообразные, 135, 209 остеодермы, 73, 151 острова, 7, 77, 118-25, 207; — гигантизм, 119-21; — карликовость, 119-24, 121рис.,122-23, 208; — континентального происхождения, 118. См. также Балеарские острова, Бали, Джерси, Кипр, Крит, Куба, Мальта, Сицилия, Средиземноморья острова, Ченнел острова; — океанические, 26, 118. См. также Гавайи, Галапагосские острова, Маврикий, Реюньон и Флорес островные виды, 26–27, 77, 207 отнятие от груди, 173 Оуэн, Ричард, сэр, 48, 130
Падиан, Кевин, 75, 77 Пакистан, 61 палеоген, 6, палеогистология, 36, 66–84; — костей динозавров, 74–77; — методы, 68; — моа, 78; — оценка размера клеток, 84; — пахиплеврозавров, 78–83, 83рис.; — птерозавров, 75, 84; — Lystrosaurus, 117-18; — Myotragus,122; — Stupendemys, 69–70, 71рис. палеозой, 6, 36, 38, 51, 84 палеоцен, 167 пальцы, 9, 148-49 панцирь черепах 34, 70, 73, 150-54 параллельная генотипическая адаптация, 3 паратаксономия, 56 Париж, Франция, 10–11, 74 парсимония, 167, 180 Патагонский шарнир, 114, 115рис. пахиплеврозавры, 78–82, 83рис. педоморфизм, 122 пеликозавры, 97, 98рис. пенсильваний (верхний карбон), 177 пермский период, пермь, 6рис., 7, 45, 59, 97, 108, 117, 159, 178, 183, 203 перья, 8–9, 19, 87, 138, 198; — у динозавров. См. динозавры пингвины, 48, 54 плакодермы, 51, 52рис., 85, 101рис., 202, 205 плакодонты, 79, 204 планктотрофия, 110-11 пластичность, 25, 122, 130-33, 163, 179-80, 201. См. также фенотипическая пластичность пластичность индивидуального развития, 76, 132-33, 163 плацентарные млекопитающие, 51, 91, 131, 143, 159рис., 166-67, 209, 211 плацентация, 50-51 плезиозавры, 53, 53рис., 79, 94, 94рис., 133 плейстоцен, 39, 60, 77, 112, 120, 123 плечевая кость, 69, 83рис. плечевой пояс, 94рис., 94, 151-52 плеченогие, 33 плод, 11, 60, 132, 171, 208 позвонки, 44, 45, 61, 155; — количество, 129-33 позвоночные, 4, 9, 15, 16, 21, 23, 43, 69, 73, 84, 85, 86, 89, 94, 112, 138, 147, 176, 190; — наземные, 9, 31, 36–38, 67; — недостающие звенья, 141-57; — онтогенез, 46–65; — число позвонков, 129-33 позвоночный столб, 49, 53, 129; — разделение на отделы, 179, 156рис. полёт, 55, активный, 147-48 полости (остеоцитов), 70, 84 Помпеи, 170 Понс де Леон, Марсия, 168, 175 Поплен, Сесиль, 68 Портманн, Адольф, 171 Португалия, 90 последовательность развития, 19 пояс задних конечностей. См. таз преобразования в индивидуальном развитии, 12, 14 преформизм, 92, 93рис. придатки, 43–45, 51, 52рис., 178, 190–191. См. также конечности Придворного Шута сценарий, 106, 207 прозауроподы, 36, 48–49. См. также Massospondylus и Plateosaurus «Происхождение видов…», книга, 13, 40 Протеро, Дон, 39, 114 псовые, 99 птенцы, ископаемые, 46–50; — недавно вылупившиеся 50, 55-56 птерозавры, 75, 84 птицы, 8, 48, 50–57, 74, 127, 127рис.,130, 138, 142, 147, 158; — бескилевые, 77–78. См. также моа; сохранение яйцерождения, 55; — выводковые, 48; — гистология, 72рис.; — зародыш, 55рис.; — нелетающие, 77, 78; — происхождение, 28, 74–77; — птенцовые, 48; — слуховые косточки, 160; — стадии развития, 24рис. Пурнелл, Марк, 62 Пэйджел, Марк, 128 пятилучевая симметрия, 33
размер генома, 84 Раймонд, Кристина, 39 райниевые черты, 191 раковины моллюсков, 32–33, 87, 111, 166-93 ракообразные, 32, 179, 191, 192рис., 193. См. также омары, крабы, и Henningsmoenicaris Ранчо Ла Бреа, 39 Рауп, Дэвид, 185 Рафф, Элизабет, 67 регионализация, 133, 156рис., 178-79, 181рис., 209 редукция конечностей, 80, 155-57, 156рис. Реисс, Джон, 26 Рейсз, Роберт, 49, 210 реконструкция эволюционного древа, 8, 31–33, 41, 63, 198 репродуктивная изоляция, 119, 140, 181 репродуктивные стратегии, 49, 50, 202-3, 211. См. также живорождение, яйцеживорождение и яйцерождение рептилии, 12, 15, 16, 28, 45–46, 52, 54, 58, 72рис., 73–80, 99, 122, 130-33, 143, 150,-51, 159рис., 159-63, 201, 203-4; — водные, 52; — полуводные, 11. См. также морские рептилии; — хористодеры, 11, 13рис. Реюньон, 119 Риппел, Оливье, 134, 200 Ричардсон, Майк, 23, 199 родительская забота, 48, 56–57, 60, 158 родительский уход, после появления на свет, 49 рождение, 29, 30, 46, 47–50, 61, 167, 168, 170 173; — в незрелом состоянии, 171; — у неандертальцев, 172 Россия, 16 рост: добавочный, 33, 69, 80–81, 87, 177, 186, 189; — геометрический подход, 93–95, 206; — математический подход, 33, 93, 95–96; — подход на основе точек-ориентиров, 94-95 роста картина/характер, 22, 36, 42, 48, 69, 72, 74–77, 96–99, 108, 163, 172, 177, 211 роста фаза, её продление, 37, 96; — её укорачивание, 77, 96 ростовая серия, 72, 95; — ископаемая, 78–79, 100–104, 161, 204 рот, 59, 99, 161, 165рис., 166, 173, 203 Рот, Луиза, 124 ротовая полость. См. рот Роу, Тим, 160, 210 рука/кисть, 14, 149; — эволюция, 9-10 руки, 69, 79. См. также конечности рыбы, 13, 16, 22, 25, 51, 59, 67, 79, 99, 118, 135, 141, 155, 158, 163, 203-4, 209. См. также данио-рерио, камбалы, карпы, колюшки, кошачьи акулы, осетрообразные, плакодермы, хрящевые рыбы, Eusthenopteron, Oryzias и Saurichthys
Саар-Нахе бассейн, 38рис. сабельный зуб, 43 саблезубая кошка. См. Smilodon fatalis саламандры, 21, 37–38, 43–44, 141 самоампутация, 43 Сан Хосито, пещера, 39 Сандер, Мартин, 69, 76 Сансом, Роберт, 62 Саттон, Марк, 191 Северная Америка, 17, 77, 112, 114, 119, 147 сегментация, 128-29, 131, 179. См. также сегменты сегменты, 22рис., 33, 45, 129, 135, 177-81, 181рис., 191-93, 209 Сеймур, остров, 190 Сент-Илер, Этьенн Жоффруа, 10 сесамовидные кости, 39-40 Сирс, Карен, 149, 207 Сибирские траппы, 108 Сибирь, 60 силурийский период, силур, 6рис., 29рис. Симпсон, Джордж Гейлорд, 4, 206 синхротронная технология, 63, 174 Сицилия, 120 скаты. См. хрящевые рыбы скелетохронология, 71, 73, 75, 81, 88; — галапагосской морской игуаны, 81–82, 82рис. скорость обмена веществ, 76, 81, 84, 87, 132, 161, 163, 164рис., 166, 208 «слоновая птица», 49-50 слоны, 76, 119-20, 121рис., 131 слуховые косточки. См. ухо Смит, Мойя, 86 Соединённые Штаты Америки, 114 солёная вода, 22, 62, 199 Соллас, Игерна Джонсон, 67 Соллас, Уильям Джонсон, 67, 203 сомиты, 22рис., 45, 129, 133 сохранение (в ископаемом состоянии), 22, 38, 43, 69–60, 64–65, 127, 167, 198; — уклон в сторону, 93, 100; — необычное, 190-93 сохранение мягких тканей, 38, 60 специализации, 21, 87, 91, 143, 147, 150 Средиземноморье, 120, 122; — острова, 118, 120, 122, 124 стадии развития, 24рис. Стокар, Рудольф, 135 Столярски, Ярослав, 183 ступни, 9, 149, 155 ступня. См. ступни сцинки, 44, 80. См. также Scincella
таз, 49, 53, 60, 130, 154, 171, 202; — безымянная кость, 156, 156рис. таксономия, 4, 38, 56, 92-104, 120, 181, 184 Танзания, 168 Таунг, ребёнок из, 168 тафономия, 62-64 Тевиссен, Ханс, 154-56 телеология, 27 темноспондилы, 21–23, 36, 59 тероподы, 7, 8, 49, 54, 76, 77 Тессье, Джордж, 95-96 Техас, США, 59, 97 Техасский Университет. См. университет Тёрвей, Сэмюэль, 78 Тичино, Швейцария, 79 Тишлигер, Хельмут, 45 Томпсон, Д’Арси, 93-95 триасовый период, триас, 6, 7, 16, 17, 52, 79, 108, 109, 117, 130, 134, 135, 138, 161, 162, 183 трилобиты, 33, 128, 176-82, 181рис.; Тюбинген, Германия, 14, 69 Тюбингенский Университет. См. университет Тюрингенского леса бассейн, 23
Уайт Ривер, свита, 114 увеличение многообразия, 26, 36, 140; — морфологического, 84, 98, 136, 138; — характер/картина, 105-25, 207-8 улитки, 109 Университет, Калифорнийский, 144 Университет, Сент-Эндрюсский, 93 Университет, Техасский, 160 Университет, Туринский, 71, Университет, Тюбингенский, 29 Университет, Цюрихский, 69, 173 Урди, Северин, 187, уродства, 11, 12, 120 Урумако, Венесуэла, 70 утки, 48; лапы, 149; число шейных позвонков, 130 утконос, 7, 51, 143, 166 ухо, 14–17, 17рис., 160, 199, 210; Уэбстер, Марк, 180, 181 Уэллс, Джон, 177 Уэльс, 17 Уэстон, Элеонор, 123
фактор роста фибробластов, 137-38, 139рис., 149; Fgf8/fgf8 149, 156 фенотипическая пластичность, 23, 35–37, 114, 187-88 фенотипы, 3, 8, 25, 35–37, 37рис., 126, 128,-29, 136-38, 140, 196, 201, 208-9 фетализации гипотеза, 170-71 филогенез, 14, 19–20, 20рис.29рис., 30рис., 31–33, 53, 64, 97, 132, 170, 184, 197-98, 210-11; — заключение в филогенетические скобки, 126-31, 127рис. Фишер, Дэн, 88 Флорес, остров, Индонезия, 123 фораминиферы, 66 Форти, Ричард, 178 Франция, 144, 182 Фридман, Мэтт, 144
Хаксли, Джулиан, 95–96, 206 Хаммер, Ойвинд, 185 Хауг, Йоахим, 191-92 Хельсинки, Финляндия, 41 хвосты, 13, 62, 79, 130, 155; — позвонки, 61; — самоампутация, 43-45 химеры. См. хрящевые рыбы хитин, 182 хоботные, 124 хомяки, 47 Хорнер, Джек, 75 хрящ, 39, 58–59, 149, 205; — обызвествлённый, 80, 83рис.; — стержень, 44-45 хрящевые рыбы, 86, 116 Хьюджи, Ясмина, 79, 81 Хьюз, Найджел, 179 Хэд, Джейсон, 116
целаканты, 13 Циммерман, Вальтер, 28 Циммермана схема, 29рис. Цюрих, Швейцария, 21, 33, 69, 81, 134, 168 Цюрихский Университет. См. университет
человекообразные обезьяны, 43, 167-71, 169рис., 195 челюсти, 11, 14–18, 17рис., 18рис., 47, 59, 99, 135, 160, 167, 199. См. также зубная кость челюстной сустав, 16–18, 17рис., 18рис., 99, 199 челюстной сустав, между чешуйчатой и зубной костями, 17 Ченнел, острова, 119 черепа, 16–21, 18рис., 41–43, 48–50, 57, 60–61, 85, 89, 94, 97-100, 102рис., 104, 120, 121рис., 134-35, 144, 151-52, 155, 165рис., 168-72, 169рис., 210 черепахи, 12, 51, 56, 73, 129, 130, 163, 164. См. также палеогистология; морские черепахи, 54; — эволюция анатомии панциря, 34, 150-54, 153рис. четвероногие, 23, 84, 117, 163 «четырёхмерная модель», 191, 192рис. чешуи, 87, 134-38, 136рис., 151 См. также чешуйчатый покров чешуйчатый покров, 136рис., 140, 209. См. также чешуи Чёрной Королевы сценарий, 106, 207 Чифелли, Ричард, 167 членистоногие, 33, 129, 176-93. См. также ракообразные и трилобиты
Швеция, 109, 191 Швейцария, 79, 107, 134, 135 Швейцарская высшая техническая школа, Цюрих, 109 Шейер, Торстен, 69, 132 шимпанзе, 5, 142, 168-74, 169рис. Шмид, Лиени, 135-38 Шотландия, 93, 191 Шох, Райнер, 36 Шпеман, Ганс, 144 Штуттгарт, Германия, 10, 36 Шульц, Адольф, 173 Шульца правило, 173, 174рис.
Эво-дево, 34–45, 135, 200-201 эволюабельность, 34, 42, 200 эволюционная биология, 5, 15, 34, 142, 144 эволюционная новизна, 9, 34 эволюционное древо, 27, 98; — ископаемых организмов, 27, 39, 60, 94, 97–99, 112, 203, 206 эволюционные инновации, 19, 31, 34, 148, 152, 200, 209 эволюционные реверсии, 53 Эймер, Теодор, 28 экзоскелет, 135, 176-77, 190, 209 экологическая биология развития, 35 эктодисплазин, 137-38, 205; — сигнальный путь, 137-40, 139рис. эктотермия, 72рис., 74–75, 122 эмаль, 85–86, 89, 91 эмбриология, 11-12 эмбрионы, 11, 24рис., 25, 46–47, 50–51, 56, 59, 61–64, 67, 86, 93рис., 144, 149, 152, 153рис., 160, 163, 199, 204-5; — аллигатора, 20; — дельфина, 156; — динозавра, 48–49, 54, 59; — зауропод, 133; ихтиозавра, 52; — летучей мыши 148-50, 15рис.; — лошади, 60; — лягушки, 21, 22рис.; — млекопитающих, 59–60; — морских ежей, 64; — птицы, 50, 55, 55рис., 55; — сумчатых, 160; — уродливые, 11–12, 13рис.; — человека, 11, 15рис. энамелоид, 85–86, эндоскелеты, 176-77 эндотермия, 56, 72рис., 74–76, 158, 162 эоцен, 27, 41, 45, 60, 97, 114-15, 115рис., 144, 147-48, 154, 190, 203 Эпель, Давид, 35 эпигенетика, 37рис., 145, 160 эпистаз, 37рис. эхолокация, 147
Юго-Восточная Азия, 120, 155 Южная Америка, 7, 60, 113-14, 206 Южная Африка, 16, 48, 168, 198 Южная Дакота, США, 60 Юкатан, полуостров, Мексика, 107 юрский период, юра 6рис., 17, 29рис., 45, 48, 52, 54, 76, 90, 116, 132, 158, 162-63, 166, 191, 210
Яблонски, Дэвид, 105 ядровые камни. См. Steinkerne яйца, 54–59, 92, 110, 116, 166-67 яйца теропод, 50 яйцеживорождение, 51, яйцерождение, 53–55, 116, 127 яйцо, задержка в организме, 56 яйцо, скорлупа, 50, 54, 56, 73–74, 167, 202-3 Япония, 120 ящерицы, 24рис., 25, 43–45, 48, 51, 54, 80–81, 118, 141, 151, 201, 202
Aepyornis, 50, 214 Afrotheria, 131 Amblyrhynchus, 80; гистология, 82рис. Apatosaurus, 76 Archaeopteryx, 28, 45, 54, 101, Arctica, 189, 190 Argyrolagus, 133рис. Australopithecus, 168, 169рис.
«Basin and Range», книга, 2 Bison, 39 Bmp. См. костный морфогенетический белок Bombina, 22рис. Bothriolepis: ростовая серия, 101рис. Brachypotherium: замещение зубов, 88рис. Branchiostoma, 62
Cambropallas, 181рис. Camelops, 39 Cetartiodactyla, 60 Conolophus, 80 Crossaster, 109 Cryptocleidus: плечевой пояс, 94рис. Cucullaea, 190
Danio. См. данио-рерио Deinosuchus, 77 Dendrobates, 22рис. Diarthrognathus, 17–18, 199 Dimetrodon, 162-63, 210 Dinornis, 77-78 Dinosauria. См. динозавры «Dinosaurs of the Flaming Cliffs», книга, 57 Diplodocus, 48 Diprotodon: ростовая серия, 102рис. Dremotherium: зубы, 90рис.
Edaphosaurus, 98рис. Elephas, 119, 121рис. Equus, 39, 98 Eryx, 53 Euryapteryx, 78 Eusthenopteron, 67, 203
Fgf8/fgf8. См. Фактор роста фибробластов «Freaks of Nature», книга, 25, 36
Gasterosteus. См. колюшки Gobiconodon, 162 Gremlin, 149
Haldanodon, 90 Henningsmoenicaris, 191-92, 192рис., Heteronectes, 145рис. Historia animalium, 93 Homininae, 167-68 Homo, 4, 123, 124, 168, 169рис.,172,174, 208. См. также неандертальцы и люди; Hoploscpahites, 187рис. Hox-гены, 130-32, 178 Hyaenodon, 90–91, 206 Hyopsodus, 41
Incisoscutum, 52рис., 202 «Into the Jungle», книга, 57
Lampetra, 62 «Leзons d’anatomie», книга, 11 Lystrosaurus, 7, 117, 159, 159рис.
Maiacetus, 61 Mamenchisaurus, 76 Mammuthus. См. мамонты Megaloceros, 89; — аллометрический рост рогов, 96рис. Merycoidodon, 60 Monodelphis, 17рис. Morganucodon, 17, 18рис., 159рис., 162 «Morphological Integration», книга, 41 Myotragus, 122, 124
Nautilus, 184 Neusticosaurus, 83рис., 204 Nimbadon, 102рис. «Not by design», книга, 26 Nothrotheriops, 39
Odontochelys, 130, 151-52, 153рис.,210 Ophiacodon, 163, 210 «Origination Patterns and Multilevel Processes in Macroevolution», книга, 105 Oryzias, 137 Oviraptor, 57, 58рис.
Panthera, 39 Paramylodon, 39 Plateosaurus, 36 Protoceratops, 97
«Recherches sur les poissons fossils», книга, 13 Rousettus, 150рис.
Sahelanthropus, 168, 169рис., 171 Sanajeh, 58 Saurichthys, 134-38, 136рис., 137рис. Scala naturae, 141 Scincella, 44 Sclerocephalus, 37–38 38рис. Sinoconodon, 162 sonic hedgehog, 86, 156, 156рис. Steinkerne, 111 Stupendemys, 69–71, 71рис.
Tanystropheus, 45, 133, 133рис. «The Lessons of History», книга, 1 Thrinaxodon, 159рис., 161 Titanoboa, 115-16 Trimerorhachis, 59 Tyrannosaurus, 8, 74, 76
Varanus, 77
Zlichovaspis, 181рис. Embryos in Deep Time. The Rock Recordof Biological Development. Marcelo Sánchez Эмбрионы из глубин времени: каменная летопись биологии развития. Санчес, Марсело Р. Издательство Калифорнийского Университета, одно из самых выдающихся университетских издательств в Соединённых Штатах, повышает качество жизни во всём мире, пропагандируя образованность в гуманитарных, общественных и естественных науках. Его деятельность поддерживают Фонд Издательства Калифорнийского Университета и безвозмездные пожертвования частных лиц и организаций. Дополнительную информацию можно получить на сайте www.ucpress.edu. Издательство Калифорнийского Университета Беркли и Лос-Анджелес, Калифорния Перевод П. Волкова, 2013 г.
Последние комментарии
1 час 20 минут назад
3 часов 7 минут назад
4 часов 21 минут назад
5 часов 26 минут назад
6 часов 35 минут назад
18 часов 33 минут назад